멜라닌
Melanin| 멜라닌 | |
|---|---|
 유멜라닌의 한 가지 가능한 구조 | |
| 재료 종류 | 이종 생체 고분자 |


멜라닌(/mmllnn/n/![]() (listen); 그리스어: μμέα,, 로마자: melas, light. 'black, dark')은 대부분의 유기체에서 발견되는 천연 색소 그룹을 가리키는 광범위한 용어이다.유멜라닌은 아미노산 티로신의 산화에 이어 중합으로 알려진 다단계 화학 과정을 통해 생산됩니다.멜라닌 색소는 멜라노사이트로 알려진 특별한 세포 그룹에서 생산됩니다.기능적으로, 유멜라닌은 자외선에 대한 보호 역할을 한다.멜라닌(뉴로멜라닌)은 유기체 내의 모든 세포 기능의 촉매 역할을 한다.
(listen); 그리스어: μμέα,, 로마자: melas, light. 'black, dark')은 대부분의 유기체에서 발견되는 천연 색소 그룹을 가리키는 광범위한 용어이다.유멜라닌은 아미노산 티로신의 산화에 이어 중합으로 알려진 다단계 화학 과정을 통해 생산됩니다.멜라닌 색소는 멜라노사이트로 알려진 특별한 세포 그룹에서 생산됩니다.기능적으로, 유멜라닌은 자외선에 대한 보호 역할을 한다.멜라닌(뉴로멜라닌)은 유기체 내의 모든 세포 기능의 촉매 역할을 한다.
멜라닌에는 5가지 기본 유형이 있습니다: 유멜라닌, 페오멜라닌, 뉴로멜라닌, 알로멜라닌, 그리고 피오멜라닌.[1]가장 일반적인 유형은 유멜라닌이며, 그 중 갈색 유멜라닌과 검은색 유멜라닌 두 종류가 있습니다.멜라노사이트가 열성형식으로 유도되어 기능하지 않을 때 생성되는 페오멜라닌은 일부 피부색 또는 모발색에 부여되는 적황색의 대부분을 담당하는 폴리벤조티아진 부분을 포함하는 시스테인 유도체이다.뉴로멜라닌은 뇌에서 발견됩니다.파킨슨병과 [2]같은 신경변성 질환의 치료 효과를 조사하기 위한 연구가 수행되었다.알로멜라닌과 피오멜라닌은 무질소 멜라닌의 두 종류이다.
인간의 피부에서는 자외선에 노출됨으로써 흑색 발생이 시작되어 피부가 검어진다.유멜라닌은 빛의 효과적인 흡수제이다. 색소는 흡수된 자외선의 [3]99.9% 이상을 방출할 수 있다.이러한 특성 때문에, 유멜라닌은 UVA와 UVB 방사선 손상으로부터 피부 세포를 보호하여 엽산 고갈과 피부 열화의 위험을 감소시키는 것으로 생각됩니다.자외선에 노출되는 것은 멜라노사이트(멜라닌 세포)의 암인 악성 흑색종의 위험 증가와 관련이 있는 것으로 간주된다.연구에 따르면 멜라닌 농도가 높은 사람, 즉 피부톤이 [4]어두운 사람일수록 피부암 발병률이 낮은 것으로 나타났다.
인간

사람에게 멜라닌은 피부색을 결정하는 주요 요인이다.그것은 또한 머리카락, 눈의 홍채 아래에 있는 색소 조직, 그리고 내이의 선조 혈관에서도 발견됩니다.뇌에서 멜라닌을 가진 조직은 뇌간 영역 내에 있는 수질과 색소를 함유하는 뉴런을 포함합니다. 예를 들어, 강압위입니다.그것은 또한 [5]부신의 망상신경대에서도 발생한다.
피부의 멜라닌은 표피의 기저층에서 발견되는 멜라노사이트에 의해 생성됩니다.일반적으로 인간은 피부에 비슷한 농도의 멜라노사이트를 가지고 있지만, 일부 개인과 민족 집단의 멜라노사이트는 다양한 양의 멜라닌을 생성한다.어떤 사람들은 그들의 몸에 멜라닌 합성이 거의 없거나 전혀 없는 상태로, [6]알비니즘이라고 알려져 있다.
멜라닌은 더 작은 성분 분자의 집합체이기 때문에, 이러한 성분 분자의 다른 비율과 결합 패턴을 가진 많은 종류의 멜라닌이 있습니다.페오멜라닌과 유멜라닌은 모두 인간의 피부와 머리카락에서 발견되지만, 유멜라닌은 사람에게 가장 풍부한 멜라닌일 뿐만 아니라 알비니즘이 [7]결핍될 가능성이 가장 높은 형태이다.
유멜라닌
유멜라닌 중합체는 오랫동안 5,6-디히드록시인돌([8]DHI)과 5,6-디히드록시인돌-2-카르본산(DHICA) 중합체를 포함하는 것으로 생각되어 왔다.
유멜라닌에는 갈색 유멜라닌과 검은색 유멜라닌 두 종류가 있습니다.이 두 종류의 유멜라닌은 화학적으로 고분자 결합 패턴이 다르다.다른 색소가 없을 때 소량의 검은 유멜라닌이 흰머리를 일으킨다.다른 색소가 없을 때 소량의 갈색 유멜라닌은 노란색([9]금발) 머리카락을 일으킨다.유멜라닌은 피부나 머리카락 등에 존재한다.
페오멜라닌

페오멜라닌(또는 파오멜라닌)은 황색에서 불그스름한 [10]색상의 범위를 제공합니다.페오멜라닌은 특히 입술, 젖꼭지, 음경 귀두, [11]질에 집중되어 있다.그렇지 않으면 금발의 원인이 될 소량의 갈색 유멜라닌이 붉은 페오멜라닌과 섞이면, 그 결과는 주황색 머리카락이 되는데, 이것은 전형적으로 "빨간색" 또는 "진저" 머리카락이라고 불립니다.페오멜라닌은 피부에도 존재하며, 결과적으로 레드헤드는 종종 그들의 피부에도 더 분홍빛을 띤다.
화학적인 측면에서 페오멜라닌은 아미노산 L-시스테인이 존재할 [12]때 DHI 및 DHICA 대신 생성되는 벤조티아진 및 벤조티아졸 단위를 포함하는 올리고머 구조가 에우멜라닌과 다르다.
트리코크롬
트리코크롬은 에멜라닌과 페오멜라닌과 같은 대사 경로에서 생성되는 색소이지만, 이러한 분자와는 달리 분자량이 낮습니다.그것들은 빨간 사람의 [13]머리카락에서 발생한다.
뉴로멜라닌
뉴로멜라닌(NM)은 뇌의 카테콜아민 작동성 뉴런의 특정 집단에서 생성되는 어두운 불용성 고분자 색소이다.인간은 가장 많은 양의 NM을 가지고 있는데, 이것은 다른 영장류에게는 적은 양으로 존재하며 다른 많은 [14]종들에는 전혀 존재하지 않습니다.인간의 NM이 철과 같은 전이 금속과 다른 잠재적으로 독성 분자와 효율적으로 결합하는 것으로 나타났지만 생물학적 기능은 아직 알려지지 않았다.그러므로, 그것은 아포토시스 [15]및 그와 관련된 파킨슨병에 결정적인 역할을 할 수 있다.
기타 생물
멜라닌은 다양한 유기체에서 매우 다양한 역할과 기능을 가지고 있습니다.멜라닌의 한 형태는 포식자에 대한 방어 메커니즘으로 많은 두족류에 의해 사용되는 잉크를 구성합니다.멜라닌은 또한 태양으로부터의 자외선과 활성산소종과 같은 세포 손상을 수반하는 스트레스로부터 박테리아와 곰팡이와 같은 미생물을 보호합니다.멜라닌은 또한 고온, 화학적 스트레스(중금속, 산화제 등) 및 생화학적 위협(침입 [16]미생물에 대한 숙주 방어 등)에 의한 손상으로부터 보호합니다.그러므로, 많은 병원성 미생물에서 멜라닌은 숙주의 면역 반응으로부터 미생물을 보호함으로써 독성과 병원성에 중요한 역할을 하는 것으로 보입니다.무척추동물에서, 침입한 병원균에 대한 선천적인 면역 방어 시스템의 주요 측면은 멜라닌을 포함한다.감염 후 몇 분 이내에, 미생물은 멜라닌(멜란화) 안에 캡슐화되어, 이 캡슐의 형성 중 유리기 부산물의 생성은 그들을 [17]죽이는 데 도움을 줄 것으로 생각됩니다.방사능 영양균이라고 불리는 몇몇 종류의 곰팡이는 그들이 감마선을 포착하고[18] 성장을 [19]위해 이 에너지를 이용할 수 있게 해주는 광합성 색소로 멜라닌을 사용할 수 있는 것으로 보입니다.
새의 어두운 깃털은 멜라닌에 기인하며, 염색되지 않은 깃털이나 카로티노이드 [20]색소를 포함한 깃털보다 박테리아에 의해 쉽게 분해되지 않습니다.멜라닌을 함유한 깃털은 또한 멜라닌 과립이 [21][22]깃털을 형성하는 케라틴 가닥 사이의 공간을 메우는데 도움을 주기 때문에 그렇지 않은 깃털보다 39% 더 마모에 강하다.조류에서 페오멜라닌 합성은 항산화 글루타티온(GSH)의 합성에 필요하지만 식단에서 과도하게 섭취하면 독성이 있을 수 있는 반필수 아미노산인 시스테인을 섭취하는 것을 의미한다.실제로 식단에서 단백질 함량이 높은 많은 육식성 조류들은 페오멜라닌을 기반으로 [23]한 색깔을 보인다.
멜라닌은 또한 포유류의 색소 [24]침착에 중요하다.포유류의 털 모양은 멜라닌의 [25][26]분포를 조절하는 아구티 유전자에 의해 결정됩니다.이 유전자의 메커니즘은 포유류의 털 [27]패턴의 다양성에 대한 통찰력을 제공하기 위해 생쥐를 대상으로 광범위하게 연구되어 왔다.
절지동물의 멜라닌은 층에 축적되어 교대 굴절률의 브래그 반사체를 생성하는 것이 관찰되었다.이 패턴의 크기가 가시광선의 파장과 일치할 때, 구조적인 색채가 발생합니다. 즉, 많은 종들에게 무지개 색을 [28]입힙니다.
거미류는 멜라닌이 쉽게 검출되지 않는 몇 안 되는 그룹 중 하나이지만, 연구자들은 거미가 실제로 멜라닌을 [29]생성한다는 데이터를 발견했습니다.
나무호랑이 나방을 포함한 몇몇 나방 종들은 그들의 체온 조절을 강화하기 위해 자원을 멜라닌으로 바꾼다.나무호랑이 나방은 위도 범위가 넓기 때문에, 더 많은 북부 개체군이 흑색화 비율을 보이는 것이 관찰되었습니다.나무호랑이 나방의 노란색과 흰색 수컷 표현형 모두에서 멜라닌이 더 많은 개체는 열을 가두는 능력이 높았지만 더 약하고 덜 효과적인 아포오스틱 [30]신호로 인해 포식율이 증가했습니다.
멜라닌은 비 UV [31]방사선에 의한 DNA 손상으로부터 드로소필라 파리와 쥐를 보호합니다.드로소필라 모델에서 중요한 연구에는 홉우드 외,[31] 1985가 포함된다.감마선에 대한 멜라닌의 방사선 방호 효과에 대한 우리의 이해의 대부분은 Irma Mosse의 [32][33][34][35][36][37][38]: 1151 실험실과 연구 그룹으로부터 온다.모세는 소련 시절 방사능 생물학을 전공했고 체르노빌에서 방사성 영양 미생물이 발견되면서 정부 자금의 지원을 받아 2022년 현재[update] 벨라루스 유전 및 세포학 [37]연구소 산하로 운영되고 있다.그녀의 가장 중요한 기여는 마우스에 대한[32][33][34][35][36][37][38]: 1151 Mosse 등, 2000년이지만 Mosse 등, 1994년,[36] Mosse 등, 1997년,[36] Mosse 등, 1998년,[35] Mosse 등, 2001년,[36] 2002년,[35][36] Mosse 등, 2006년,[35][36] Mosse 등, 2007년[36], Mosse 등, 2008년 [36]등이다.
식물

식물에 의해 생성된 멜라닌은 알칼리 융합에서 카테콜을 생산할 수 있기 때문에 때때로 '카테콜 멜라닌'이라고 불립니다.바나나와 같은 과일의 효소적 갈변에서 흔히 볼 수 있다.밤껍질 멜라닌은 항산화 [39]및 착색제로 사용할 수 있다.생합성에는 티로신 및 카테콜아민으로부터의 티로시나아제형 폴리페놀산화효소에 의해 인돌-5,6-퀴논이 산화되어 카테콜 멜라닌이 생성된다.그럼에도 불구하고 많은 식물들은 멜라닌의 [40]생성을 억제하는 화합물을 포함하고 있다.
단일 단량체로 해석
멜라닌은 단일한 구조나 화학측정법을 가지고 있지 않다는 것이 현재 알려져 있다.그럼에도 불구하고 PubChem과 같은 화학 데이터베이스에는 구조식 및 경험식(일반적으로 3,8-디메틸-2,7-디히드로벤조[1,2,3-cd:4,5,6-cdd]]디인돌-4,5,9,10-테트론)이 포함된다.이것은 측정된 원소 성분과 멜라닌의 일부 특성을 설명하는 단일 단량체로 생각될 수 있지만,[41] 자연에서는 발견되지 않을 것 같습니다.솔라노는[42] 이 잘못된 추세가 [43]1948년의 경험적 공식의 보고에서 비롯되었다고 주장하지만, 다른 역사적 세부 사항은 제공하지 않는다.
![3,8-Dimethyl-2,7-dihydrobenzo[1,2,3-cd:4,5,6-c′d′]diindole-4,5,9,10-tetrone](/wiki/File:Melanin.svg) | |
![3,8-Dimethyl-2,7-dihydrobenzo[1,2,3-cd:4,5,6-c′d′]diindole-4,5,9,10-tetrone ball and stick model](/wiki/File:Melanin_ball_and_stick.png) | |
| 이름 | |
|---|---|
| 우선 IUPAC 이름 3,8-디메틸-2,7-디히드로벤조[1,2,3-cd:4,5,6-cdd]]디인돌-4,5,9,10-테트론 | |
| 식별자 | |
| 켐스파이더 | |
PubChem CID | |
| 특성. | |
| C18H10N2O4 | |
| 몰 질량 | 318.288g/120−1 |
| 밀도 | 1.6~1.8g/cm3 |
| 녹는점 | -20 °C 미만(-4 °F, 253 K) |
| 비등점 | 450 ~ 550 °C (842 ~1,022 °F, 723 ~823 K) |
달리 명시되지 않은 한 표준 상태(25°C[77°F], 100kPa)의 재료에 대한 데이터가 제공됩니다. | |
생합성 경로
유멜라닌과 페오멜라닌 모두에 대한 생합성 경로의 첫 단계는 티로시나아제에 [44]의해 촉매된다.
도파퀴논은 벤조티아진과 페오멜라닌으로 가는 두 가지 경로를 통해 시스테인과 결합할 수 있다.
- 도파퀴논+시스테인→5-S-시스테인도파→벤조티아진 중간체→페오멜라닌
- 도파퀴논+시스테인→2-S-시스테인도파→벤조티아진 중간체→페오멜라닌
또한, 도파퀴논은 류코도파크롬으로 전환될 수 있고 에멜라닌으로 가는 두 가지 경로를 따라갈 수 있다.
- 도파퀴논 → 류코도파크롬 → 도파크롬 → 5,6-디히드록시인돌-2-디옥실산 → 퀴논 → 유멜라닌
- 도파퀴논 → 류코도파크롬 → 도파크롬 → 5,6-디히드록시인돌 → 퀴논 → 유멜라닌
상세한 대사 경로는 KEGG 데이터베이스에서 확인할 수 있다(외부 링크 참조).

L-티로신

L-DOPA
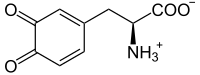
L-도파퀴논

L-류코도파크롬

L-도파크롬

.svg)



현미경 외관
멜라닌은 갈색이며, 굴절되지 않으며, 800나노미터 미만의 직경을 가진 개별 과립을 가지고 있습니다.이것은 멜라닌을 더 크고, 걸쭉하고, 굴절성이 있으며, 녹색에서 노란색 또는 적갈색까지 다양한 일반적인 혈액 분해 색소와 구별합니다.색소가 많이 들어간 병변에서는 멜라닌의 밀도가 높은 집합체가 조직학적 세부사항을 모호하게 할 수 있습니다.과망간산칼륨 희석액은 효과적인 멜라닌 [45]표백제이다.
유전적 장애 및 질병 상태
안구피부알비닌증에는 대략 9가지 종류가 있는데, 대부분 상염색체 열성 질환이다.어떤 민족은 다른 형태의 발병률이 더 높다.예를 들어, OCA2라고 불리는 가장 흔한 유형은 흑인 아프리카 혈통과 백인 유럽인들 사이에서 특히 자주 발생한다.OCA2를 가진 사람들은 보통 하얀 피부를 가지고 있지만 종종 OCA1만큼 창백하지 않다.그들(OCA2 또는 OCA1? 역사상의 코멘트 참조)은 옅은 금발에서 황금색, 딸기색 금발 또는 갈색 머리를 가지고 있으며, 가장 일반적으로 푸른 눈을 가지고 있다.현대 유럽인의 98.7-100%는 비염색성 안피부 알빈증의 원인인 파생알레알레르 SLC24A5의 매개체이다.피부, 머리카락, 눈에 멜라닌 색소가 선천적으로 감소하거나 없는 것이 특징인 상염색체 열성 질환입니다.흑인의 OCA2의 추정 빈도는 10,000분의 1로 백인 미국인의 [46]36,000분의 1과 대조됩니다.일부 아프리카 국가에서는 2000명 중 1명에서 5000명 [47]중 1명꼴로 발병 빈도가 더 높다.알비니즘의 또 다른 형태인 "황색 안피성 알비니즘"은 주로 스위스와 독일 혈통을 가진 아미시인들 사이에서 더 널리 퍼져 있는 것으로 보인다.이 IB형 질환을 가진 사람들은 태어날 때 보통 하얀 머리와 피부를 가지고 있지만,[47] 유아기에 정상적인 피부 색소가 빠르게 발달합니다.
안구 알빈증은 눈의 색소 침착뿐만 아니라 시력에도 영향을 미칩니다.알비니즘이 있는 사람들은 보통 20/60에서 20/400 범위 내에서 검사를 잘 하지 못한다.게다가, 푸에르토리코 출신 사람들 사이에서 약 2,700명 중 1명꼴로 가장 많이 발생하는 두 가지 형태의 알비니즘은 흑색종과 관련된 사망 이상의 사망률과 관련이 있다.
알비니즘과 귀머거리 사이의 연관성은 잘 알려져 있지만 잘 알려져 있지 않다.찰스 다윈은 1859년에 쓴 '종의 기원에 관한 논문'에서 "완전히 하얗고 푸른 눈을 가진 고양이는 일반적으로 [48]귀가 들리지 않는다"고 말했다.인간의 경우, 저색소 침착과 난청은 북미의 [49]호피족 사이에서 주로 관찰되는 희귀한 바덴버그 증후군에서 함께 발생한다.호피 인디언의 알비니즘 발병률은 약 200명 중 1명으로 추정되고 있다.개와 설치류를 포함한 다른 포유동물에서도 비슷한 형태의 알비니즘과 청각장애가 발견되었다.그러나 멜라닌 합성에 필요한 효소가 부족한 대부분의 사람들은 정상적인 청각 [50]기능을 가지고 있기 때문에 멜라닌 그 자체가 저색소 침착과 관련된 난청의 직접적인 원인이 되는 것으로 보이지 않는다.대신, 내이의 선조 혈관 내에 멜라노사이트가 없는 것은 [51]달팽이관 장애를 초래하지만, 그 이유는 완전히 이해되지 않는다.
신경운동 기능에 영향을 미치는 질환인 파킨슨병은 도파민성 및 노르아드레날린성 색소 신경세포의 특이적 탈락의 결과로 흑색소 및 강압궤적에서 뉴로멜라닌이 감소한다.이것은 도파민과 노르에피네프린 합성을 감소시킨다.인종과 흑인의 뉴로멜라닌 수치 사이의 상관관계가 보고되지 않았지만, 백인에 비해 흑인의 파킨슨병 발병률이 현저히 낮은 것은 피부 멜라닌이 흑인의 뉴로멜라닌을 외부 [52]독소로부터 보호하는 역할을 할 수 있다는 것을 암시했다.
멜라닌 결핍 외에 멜라닌 중합체의 분자량은 산화 스트레스, 빛에 대한 노출, 멜라닌 기질 단백질과의 관련성 섭동, pH의 변화 또는 국소적인 금속 이온 농도와 같은 다양한 요인에 의해 감소될 수 있다.정상 항산화성 폴리머를 친산화성 폴리머로 변화시키기 위해 분자량의 감소 또는 안구 멜라닌의 중합도의 감소가 제안되었다.멜라닌은 산화방지제 상태에서 황반변성과 [53]흑색종의 원인 및 진행에 관여하는 것으로 알려져 있다.파킨슨병의 중요한 단발성 치료제인 라사길린은 멜라닌 결합성, 흑색종양 감소성 [54]등을 가지고 있다.
그러나, 높은 유멜라닌 수치는 비타민 D 결핍에 대한 높은 기질을 넘어 단점이 될 수 있습니다.검은 피부는 좌현 얼룩을 레이저로 제거하는 복잡한 요인입니다.일반적으로 백피부 치료에 효과적인 레이저는 아시아계 또는 아프리카계 혈통의 좌현 얼룩 제거에 덜 성공합니다.피부가 어두운 사람에게 고농도의 멜라닌은 단순히 레이저 방사선을 확산시키고 흡수하여 표적 조직에 의한 빛의 흡수를 방해한다.비슷한 방법으로, 멜라닌은 피부가 어두운 사람들에게 다른 피부 질환의 레이저 치료를 복잡하게 만들 수 있다.
주근깨와 점들은 피부에 멜라닌이 국소적으로 농축된 곳에 형성된다.그들은 창백한 피부와 매우 관련이 있다.
니코틴은 멜라닌 합성에 있어서의 전구체 기능이나 멜라닌의 불가역적인 결합으로 인해 멜라닌 함유 조직에 친화력을 가진다.이것은 니코틴 의존성의 증가를 뒷받침하고 어두운 색소의 [55]사람들에게서 금연율을 낮추기 위해 제안되었다.
인간의 적응
생리학
멜라노사이트는 멜라닌 과립을 멜라닌이라고 불리는 특별한 세포 소포에 삽입합니다.그리고 나서 이것들은 인간 표피의 각질세포로 옮겨진다.각 수용 세포에 있는 멜라노솜은 세포핵 위에 축적되어 태양의 자외선의 이온화 방사선에 의해 야기되는 돌연변이로부터 핵 DNA를 보호합니다.일반적으로, 적도에 가까운 지구 지역에서 오랜 기간 동안 조상들이 살았던 사람들의 피부에는 더 많은 양의 유멜라닌이 있다.이것은 그들의 피부를 갈색이나 검은색으로 만들고 밝은 피부의 사람들에게 [56]흑색종을 더 자주 일으키는 높은 수준의 태양 노출로부터 그들을 보호합니다.
색소침착의 모든 효과가 유리한 것은 아니다.색소침착은 더운 기후에서 열 부하를 증가시키고, 어두운 피부를 가진 사람들은 매우 밝은 피부를 가진 사람들보다 햇빛으로부터 30% 더 많은 열을 흡수하지만, 이 요소는 더 많은 양의 땀으로 상쇄될 수 있습니다.추운 기후에서 어두운 피부는 방사선에 의해 더 많은 열 손실을 수반합니다.색소침착은 또한 비타민 D의 합성을 방해한다.색소침착이 열대지방의 생활에 전적으로 이롭지 않은 것으로 보이기 때문에, 그 생물학적 중요성에 대한 다른 가설들이 발전해 왔다. 예를 들어 기생충과 열대성 [57]질병에 대한 적응에 의해 유발되는 이차적인 현상이다.
진화 기원
초기 인류는 약 120만 년 전에 자외선의 영향을 증가시킨 체모 감소에 적응하면서 피부색이 어두워지도록 진화했다.털이 없는 것이 발달하기 전에, 초기 인류는 다른 [58]영장류에서 볼 수 있는 것과 비슷하게 털 아래 피부가 상당히 밝았다.가장 최근의 과학적 증거는 해부학적으로 현대 인류가 20만 년에서 10만 [59]년 사이에 아프리카에서 진화했고, 그리고 나서 80,000년에서 50,000년 사이의 한 번의 이동을 통해 세계의 나머지 지역에 특정 고대 인류와 교배했다는 것을 보여준다.[60]최초의 현생 인류는 비교적 많은 수의 유멜라닌을 생성하는 멜라노사이트를 가지고 있었으며, 오늘날 아프리카의 원주민들과 비슷한 어두운 피부를 만들어냈을 것으로 보인다.이러한 초기 사람들 중 일부가 아시아와 유럽 지역에 이주하고 정착함에 따라, 태양으로부터의 방사선이 덜 강한 기후에서 유멜라닌 생산에 대한 선택적 압력이 감소하였다.이것은 결국 현재의 인간 피부색 범위를 만들어냈다.인간의 창백한 피부와 관련된 것으로 알려진 두 가지 일반적인 유전자 변종 중 Mc1r은 양성 선택을 [61]받지 않은 것으로 보이는 반면 SLC24A5는 양성 선택을 [62]받은 것으로 보인다.
영향들
북쪽으로 이주한 사람들처럼, 밝은 피부를 가진 사람들은 훨씬 더 강한 태양 방사선에 적응한다.자연은 자외선이 약할 때 멜라닌을 적게 선택한다.대부분의 사람들의 피부는 자외선에 노출되면 검어지며, 필요할 때 더 많은 보호를 준다.이것이 선탠의 생리적인 목적이다.피부를 보호하는 더 많은 유멜라닌을 생산하는 어두운 피부의 사람들은 햇빛에 의한 더 큰 보호와 피부암의 잠재적으로 치명적인 형태인 흑색종의 발생, 그리고 리보플라빈, 카로티노이드와 같은 특정 비타민의 광분해와 같은 강한 태양 방사선에 노출되는 것과 관련된 다른 건강 문제를 가지고 있습니다.코페롤과 [63]엽산입니다
눈의 멜라닌, 홍채, 맥락막은 자외선 및 고주파 가시광선으로부터 그들을 보호하는데 도움을 준다; 회색, 파란색, 녹색 눈을 가진 사람들은 태양과 관련된 눈 문제의 위험이 더 높다.또, 시간이 지남에 따라 눈의 수정체가 노랗게 되어, 한층 더 보호된다.그러나 렌즈는 시간이 지남에 따라 경직되어, 아마도 UV 노출로 인한 단백질 가교로 인한 손상(먼 곳에서 가까운 곳으로 초점을 맞추기 위해 모양을 바꾸는 능력)의 대부분을 잃습니다.
최근의 연구는 멜라닌이 광보호 [64]이외의 보호 역할을 할 수 있다는 것을 시사한다.멜라닌은 강력한 킬레이트 리간드 에틸렌디아민테트라아세테이트(EDTA)보다 훨씬 더 효율적으로 카르본산 및 페놀성 수산기를 통해 금속 이온을 킬레이트할 수 있다.따라서 독성이 있을 수 있는 금속 이온을 격리하여 세포의 나머지 부분을 보호할 수 있습니다.이 가설은 파킨슨병에서 관찰된 뉴로멜라닌의 손실이 뇌의 철분 수치 증가를 동반한다는 사실에 의해 뒷받침된다.
물리적 특성 및 기술 응용 프로그램
매트릭스 발판 멜라노단백질에 [65]공유결합된 고도로 가교된 헤테로폴리머를 지지하는 증거가 존재한다.멜라닌이 항산화 물질로 작용하는 능력은 멜라닌의 중합 정도 [66]또는 분자량에 정비례한다고 제안되어 왔다.멜라닌 단량체의 효과적인 중합에 대한 차선의 조건은 황반변성 및 [67]흑색종의 원인 및 진행에 관여한 저분자량의 친산화성 멜라닌의 형성을 초래할 수 있다.망막색소상피(RPE)에서 흑색화를 업 레귤레이션하는 시그널링 경로도 RPE에 의한 로드 외측 세그먼트 식세포증의 다운 레귤레이션에 관련될 수 있다.이 현상은 부분적으로 [68]황반변성에서의 포볼 스페어링에 기인한다.
흑색종 전이에서의 역할
Sarna의 연구팀이 실시한 연구는 색소가 많이 들어간 흑색종 세포는 약 4.93의 영률을 가지고 있는 반면, 색소가 없는 세포에서는 0.[69]98에 불과하다는 것을 증명했다.또 다른 실험에서 그들은 흑색종 세포의 탄성이 전이 및 성장에 중요하다는 것을 발견했다. 즉, 색소화되지 않은 종양이 색소화 된 종양보다 더 크고 확산되는 것이 훨씬 쉬웠다.그들은 흑색종 종양에는 색소세포와 색소세포가 모두 있기 때문에 약물에 내성이 있고 [69]전이될 수 있다는 것을 보여주었다.
「 」를 참조해 주세요.
레퍼런스
- ^ Cao, Wei; Zhou, Xuhao; McCallum, Naneki C.; Hu, Ziying; Ni, Qing Zhe; Kapoor, Utkarsh; Heil, Christian M.; Cay, Kristine S.; Zand, Tara; Mantanona, Alex J.; Jayaraman, Arthi (9 February 2021). "Unraveling the Structure and Function of Melanin through Synthesis". Journal of the American Chemical Society. 143 (7): 2622–2637. doi:10.1021/jacs.0c12322. hdl:1854/LU-8699336. ISSN 0002-7863. PMID 33560127. S2CID 231872855.
- ^ Haining, Robert L.; Achat-Mendes, Cindy (March 2017). "Neuromelanin, one of the most overlooked molecules in modern medicine, is not a spectator". Neural Regeneration Research. 12 (3): 372–375. doi:10.4103/1673-5374.202928. PMC 5399705. PMID 28469642.
- ^ Meredith P, Riesz J (2004). "Radiative relaxation quantum yields for synthetic eumelanin". Photochemistry and Photobiology. 79 (2): 211–6. arXiv:cond-mat/0312277. doi:10.1111/j.1751-1097.2004.tb00012.x. PMID 15068035. S2CID 222101966.
- ^ Brenner M, Hearing VJ (2008). "The protective role of melanin against UV damage in human skin". Photochemistry and Photobiology. 84 (3): 539–49. doi:10.1111/j.1751-1097.2007.00226.x. PMC 2671032. PMID 18435612.
- ^ Solano, F. (2014). "Melanins: Skin Pigments and Much More—Types, Structural Models, Biological Functions, and Formation Routes". New Journal of Science. 2014: 1–28. doi:10.1155/2014/498276.
- ^ Cichorek, Mirosława; Wachulska, Małgorzata; Stasiewicz, Aneta; Tymińska, Agata (20 February 2013). "Skin melanocytes: biology and development". Advances in Dermatology and Allergology. 30 (1): 30–41. doi:10.5114/pdia.2013.33376. PMC 3834696. PMID 24278043.
- ^ "oculocutaneous albinism". Genetics Home Reference. Retrieved 25 September 2017.
- ^ Meredith, Paul; Sarna, Tadeusz (1 December 2006). "The physical and chemical properties of eumelanin". Pigment Cell Research. 19 (6): 572–594. doi:10.1111/j.1600-0749.2006.00345.x. PMID 17083485.
- ^ Ito, S.; Wakamatsu, K. (December 2011). "Diversity of human hair pigmentation as studied by chemical analysis of eumelanin and pheomelanin". Journal of the European Academy of Dermatology and Venereology. 25 (12): 1369–1380. doi:10.1111/j.1468-3083.2011.04278.x. ISSN 1468-3083. PMID 22077870. S2CID 5121042.
- ^ Slominski A, Tobin DJ, Shibahara S, Wortsman J (October 2004). "Melanin pigmentation in mammalian skin and its hormonal regulation". Physiological Reviews. 84 (4): 1155–228. doi:10.1152/physrev.00044.2003. PMID 15383650. S2CID 21168932.
- ^ "pheomelanin". MetaCyc Metabolic Pathway Database. 2010.[인용필수]
- ^ Greco G, Panzella L, Verotta L, d'Ischia M, Napolitano A (April 2011). "Uncovering the structure of human red hair pheomelanin: benzothiazolylthiazinodihydroisoquinolines as key building blocks". Journal of Natural Products. 74 (4): 675–82. doi:10.1021/np100740n. PMID 21341762.
- ^ Prota, G.; Searle, A. G. (1978). "Biochemical sites of gene action for melanogenesis in mammals". Annales de Génétique et de Sélection Animale. 10 (1): 1–8. doi:10.1186/1297-9686-10-1-1. PMC 2757330. PMID 22896083.
- ^ Fedorow H, Tribl F, Halliday G, Gerlach M, Riederer P, Double KL (2005). "Neuromelanin in human dopamine neurons: comparison with peripheral melanins and relevance to Parkinson's disease". Prog Neurobiol. 75 (2): 109–124. doi:10.1016/j.pneurobio.2005.02.001. PMID 15784302. S2CID 503902.
- ^ Double KL (2006). "Functional effects of neuromelanin and synthetic melanin in model systems". J Neural Transm. 113 (6): 751–756. doi:10.1007/s00702-006-0450-5. PMID 16755379. S2CID 23096297.
- ^ Hamilton AJ, Gomez BL (March 2002). "Melanins in fungal pathogens". Journal of Medical Microbiology. 51 (3): 189–91. doi:10.1099/0022-1317-51-3-189. PMID 11871612.
- ^ Cerenius L, Söderhäll K (April 2004). "The prophenoloxidase-activating system in invertebrates". Immunological Reviews. 198: 116–26. doi:10.1111/j.0105-2896.2004.00116.x. PMID 15199959. S2CID 10614298.
- ^ Castelvecchi, Davide (26 May 2007). "Dark Power: Pigment seems to put radiation to good use". Science News. 171 (21): 325. doi:10.1002/scin.2007.5591712106.
- ^ Dadachova E, Bryan RA, Huang X, et al. (2007). "Ionizing radiation changes the electronic properties of melanin and enhances the growth of melanized fungi". PLOS ONE. 2 (5): e457. Bibcode:2007PLoSO...2..457D. doi:10.1371/journal.pone.0000457. PMC 1866175. PMID 17520016.
- ^ Gunderson, Alex R.; Frame, Alicia M.; Swaddle, John P.; Forsyth, Mark H. (1 September 2008). "Resistance of melanized feathers to bacterial degradation: is it really so black and white?". Journal of Avian Biology. 39 (5): 539–545. doi:10.1111/j.0908-8857.2008.04413.x.
- ^ Bonser, Richard H. C. (1995). "Melanin and the Abrasion Resistance of Feathers". Condor. 97 (2): 590–591. doi:10.2307/1369048. JSTOR 1369048.
- ^ Galván, Ismael; Solano, Francisco (8 April 2016). "Bird Integumentary Melanins: Biosynthesis, Forms, Function and Evolution". International Journal of Molecular Sciences. 17 (4): 520. doi:10.3390/ijms17040520. PMC 4848976. PMID 27070583.
- ^ Rodríguez‐Martínez, Sol; Galván, Ismael (2020). "Juvenile pheomelanin-based plumage coloration has evolved more frequently in carnivorous species". Ibis. 162 (1): 238–244. doi:10.1111/ibi.12770. hdl:10261/207451. ISSN 1474-919X. S2CID 202018215.
- ^ Jimbow, K; Quevedo WC, Jr; Fitzpatrick, TB; Szabo, G (July 1976). "Some aspects of melanin biology: 1950–1975". The Journal of Investigative Dermatology. 67 (1): 72–89. doi:10.1111/1523-1747.ep12512500. PMID 819593.
- ^ Meneely, Philip (2014). Genetic Analysis: Genes, Genomes, and Networks in Eukaryotes. Oxford University Press. ISBN 9780199681266.
- ^ Griffiths, Anthony JF; Miller, Jeffrey H.; Suzuki, David T.; Lewontin, Richard C.; Gelbart, William M. (2000). "Gene interaction in coat color of mammals".
{{cite journal}}:Cite 저널 요구 사항journal=(도움말) - ^ Millar, S. E.; Miller, M. W.; Stevens, M. E.; Barsh, G. S. (October 1995). "Expression and transgenic studies of the mouse agouti gene provide insight into the mechanisms by which mammalian coat color patterns are generated". Development. 121 (10): 3223–3232. doi:10.1242/dev.121.10.3223. PMID 7588057.
- ^ Neville, A. C. (2012). Biology of the Arthropod Cuticle. Springer Science & Business Media. ISBN 9783642809101.
- ^ Hsiung, B.-K.; Blackledge, T. A.; Shawkey, M. D. (2015). "Spiders do have melanin after all". Journal of Experimental Biology. 218 (22): 3632–3635. doi:10.1242/jeb.128801. PMID 26449977.
- ^ Hegna, Robert H.; Nokelainen, Ossi; Hegna, Jonathan R.; Mappes, Johanna (2013). "To quiver or to shiver: increased melanization benefits thermoregulation, but reduces warning signal efficacy in the wood tiger moth". Proc. R. Soc. B. 280 (1755): 20122812. doi:10.1098/rspb.2012.2812. PMC 3574392. PMID 23363631.
- ^ a b Mosse, Irma B.; Dubovic, Boris V.; Plotnikova, Svetlana I.; Kostrova, Ludmila N.; Molophei, Vadim; Subbot, Svetlana T.; Maksimenya, Inna P. (20–25 May 2001). Obelic, B.; Ranogajev-Komor, M.; Miljanic, S.; Krajcar Bronic, I. (eds.). Melanin is Effective Radioprotector against Chronic Irradiation and Low Radiation Doses. IRPA Regional Congress on Radiation Protection in Central Europe: Radiation Protection and Health. INIS. Dubrovnik (Croatia): Croatian Radiation Protection Association. p. 35 (of 268).
- ^ a b Gessler, N. N.; Egorova, A. S.; Belozerskaya, T. A. (2014). "Melanin pigments of fungi under extreme environmental conditions (Review)". Applied Biochemistry and Microbiology. Pleiades Publishing. 50 (2): 105–113. doi:10.1134/s0003683814020094. ISSN 0003-6838. PMID 25272728. S2CID 8570835.
- ^ a b Nenoi, M; Wang, B; Vares, G (12 June 2014). "In vivo radioadaptive response". Toxicology. Sage. 34 (3): 272–283. doi:10.1177/0960327114537537. ISSN 0960-3271. PMC 4442823. PMID 24925363.
- ^ a b Liu, Heng; Yang, Youyuan; Liu, Yu; Pan, Jingjing; Wang, Junqing; Man, Fengyuan; Zhang, Weiguo; Liu, Gang (7 February 2020). "Melanin‐Like Nanomaterials for Advanced Biomedical Applications: A Versatile Platform with Extraordinary Promise". Advanced Science. Wiley-VCH. 7 (7): 1903129. doi:10.1002/advs.201903129. ISSN 2198-3844. PMC 7141020. PMID 32274309.
- ^ a b c d e Mosse, Irma B. (2012). "Genetic effects of ionizing radiation – some questions with no answers". Journal of Environmental Radioactivity. Elsevier. 112: 70–75. doi:10.1016/j.jenvrad.2012.05.009. ISSN 0265-931X. PMID 22683898.
- ^ a b c d e f g h i Mosse, Irma; Kilchevsky, Alexander; Nikolova, Nevena; Zhelev, Nikolai (14 December 2016). "Some problems and errors in cytogenetic biodosimetry". Biotechnology & Biotechnological Equipment. Taylor & Francis. 31 (3): 460–468. doi:10.1080/13102818.2016.1259018. ISSN 1310-2818. S2CID 59398089.
- ^ a b c Mosse, Irma (18 January 2022). "Radiobiology in my life – Irma Mosse". International Journal of Radiation Biology. Taylor & Francis. 98 (3: Women in Radiobiology): 474–478. doi:10.1080/09553002.2022.2026517. ISSN 0955-3002. PMID 34994663. S2CID 245823003.
- ^ a b Dadachova, Ekaterina; Casadevall, Arturo (2011). Horikoshi, Kōki (ed.). Extremophiles handbook. Tokyo New York City: Springer. pp. xxix+1247. ISBN 978-4-431-53898-1. OCLC 700199222. ISBN 978-4-431-53897-4.OCLC 711778164.
- ^ Yao, Zeng-Yu; Qi, Jian-Hua (22 April 2016). "Comparison of Antioxidant Activities of Melanin Fractions from Chestnut Shell". Molecules. 21 (4): 487. doi:10.3390/molecules21040487. PMC 6273334. PMID 27110763.
- ^ Kim, Y.-J.; Uyama, H. (15 May 2005). "Tyrosinase inhibitors from natural and synthetic sources: structure, inhibition mechanism and perspective for the future". Cellular and Molecular Life Sciences. 62 (15): 1707–1723. doi:10.1007/s00018-005-5054-y. PMID 15968468. S2CID 8280251.
- ^ Solano, F. (2014). "Melanins: Skin Pigments and Much More—Types, Structural Models, Biological Functions, and Formation Routes". New Journal of Science. 2014 (498276): 1–28. doi:10.1155/2014/498276.
- ^ Solano, F. (2014). "Melanins: Skin Pigments and Much More—Types, Structural Models, Biological Functions, and Formation Routes". New Journal of Science. 2014 (498276): 1–28. doi:10.1155/2014/498276.
- ^ Mason, H. S. (1948). "The chemistry of melanin. Mechanism of the oxidation of dihydroxyphenylalanine by tyrosinase". Journal of Biological Chemistry. 172 (1): 83–99. doi:10.1016/S0021-9258(18)35614-X. PMID 18920770.
- ^ Zaidi, Kamal Uddin; Ali, Ayesha S.; Ali, Sharique A.; Naaz, Ishrat (2014). "Microbial Tyrosinases: Promising Enzymes for Pharmaceutical, Food Bioprocessing, and Environmental Industry". Biochemistry Research International. 2014: 1–16 (see Fig. 3). doi:10.1155/2014/854687. PMC 4033337. PMID 24895537.
- ^ "Melanin". pubchem.ncbi.nlm.nih.gov. Retrieved 25 September 2017.
- ^ "Oculocutaneous Albinism". Archived from the original on 23 December 2008.
- ^ a b "Ocular Manifestations of Albinism: Background, Pathophysiology, Epidemiology". 18 June 2018 – via eMedicine.
{{cite journal}}:Cite 저널 요구 사항journal=(도움말) - ^ "Causes of Variability". Archived from the original on 23 September 2006. Retrieved 18 September 2006.
- ^ 엔트레스진 300700
- ^ 엔트레스진 606933
- ^ Cable J, Huszar D, Jaenisch R, Steel KP (February 1994). "Effects of mutations at the W locus (c-kit) on inner ear pigmentation and function in the mouse". Pigment Cell Research. 7 (1): 17–32. doi:10.1111/j.1600-0749.1994.tb00015.x. PMID 7521050.
- ^ "Lewy Body Disease". Archived from the original on 21 July 2009.
- ^ Meyskens FL, Farmer P, Fruehauf JP (June 2001). "Redox regulation in human melanocytes and melanoma" (PDF). Pigment Cell Research. 14 (3): 148–54. doi:10.1034/j.1600-0749.2001.140303.x. PMID 11434561.
- ^ Meier-Davis SR, Dines K, Arjmand FM, et al. (December 2012). "Comparison of oral and transdermal administration of rasagiline mesylate on human melanoma tumor growth in vivo". Cutaneous and Ocular Toxicology. 31 (4): 312–7. doi:10.3109/15569527.2012.676119. PMID 22515841. S2CID 30344869.
- ^ King G, Yerger VB, Whembolua GL, Bendel RB, Kittles R, Moolchan ET (June 2009). "Link between facultative melanin and tobacco use among African Americans". Pharmacology Biochemistry and Behavior. 92 (4): 589–96. doi:10.1016/j.pbb.2009.02.011. PMID 19268687. S2CID 3070838.
- ^ "Human Skin Color Variation". The Smithsonian Institution's Human Origins Program. 20 June 2012. Retrieved 24 August 2019.
- ^ Berth-Jones, J. (2010), "Constitutive pigmentation, human pigmentation and the response to sun exposure", in Tony Burns; Stephen Breathnach; Neil Cox; Christopher Griffiths (eds.), Rook's Textbook of Dermatology, vol. 3 (8th ed.), Wiley-Blackwell, p. 58.9, ISBN 978-1-4051-6169-5
- ^ Wade, Nicholas (19 August 2003). "Why Humans and Their Fur Parted Ways". The New York Times. ISSN 0362-4331. Retrieved 24 August 2019.
- ^ Tishkoff SA, Reed FA, Friedlaender FR, et al. (May 2009). "The genetic structure and history of Africans and African Americans". Science. 324 (5930): 1035–44. Bibcode:2009Sci...324.1035T. doi:10.1126/science.1172257. PMC 2947357. PMID 19407144.
- ^ "A Single Migration From Africa Populated the World, Studies Find". The New York Times. 22 September 2016.
- ^ Harding RM, Healy E, Ray AJ, et al. (April 2000). "Evidence for variable selective pressures at MC1R". American Journal of Human Genetics. 66 (4): 1351–61. doi:10.1086/302863. PMC 1288200. PMID 10733465.
- ^ Lamason RL, Mohideen MA, Mest JR, et al. (December 2005). "SLC24A5, a putative cation exchanger, affects pigmentation in zebrafish and humans". Science. 310 (5755): 1782–6. Bibcode:2005Sci...310.1782L. doi:10.1126/science.1116238. PMID 16357253. S2CID 2245002.
- ^ Jablonski, Nina G.; Chaplin, George (11 May 2010). "Human skin pigmentation as an adaptation to UV radiation". Proceedings of the National Academy of Sciences. 107 (Supplement 2): 8962–8968. Bibcode:2010PNAS..107.8962J. doi:10.1073/pnas.0914628107. PMC 3024016. PMID 20445093.
- ^ Liu Y, Hong L, Kempf VR, Wakamatsu K, Ito S, Simon JD (June 2004). "Ion-exchange and adsorption of Fe(III) by Sepia melanin". Pigment Cell Research. 17 (3): 262–9. doi:10.1111/j.1600-0749.2004.00140.x. PMID 15140071.
- ^ Donatien PD, Orlow SJ (August 1995). "Interaction of melanosomal proteins with melanin". European Journal of Biochemistry. 232 (1): 159–64. doi:10.1111/j.1432-1033.1995.tb20794.x. PMID 7556145.
- ^ Sarangarajan R, Apte SP (2005). "Melanin aggregation and polymerization: possible implications in age-related macular degeneration". Ophthalmic Research. 37 (3): 136–41. doi:10.1159/000085533. PMID 15867475. S2CID 27499198.
- ^ Meyskens FL, Farmer PJ, Anton-Culver H (April 2004). "Etiologic pathogenesis of melanoma: a unifying hypothesis for the missing attributable risk" (PDF). Clinical Cancer Research. 10 (8): 2581–3. doi:10.1158/1078-0432.ccr-03-0638. PMID 15102657. S2CID 26079375.
- ^ Sarangarajan R, Apte SP (2005). "Melanization and phagocytosis: implications for age related macular degeneration". Molecular Vision. 11: 482–90. PMID 16030499.
- ^ a b Sarna, Michal; Krzykawska-Serda, Martyna; Jakubowska, Monika; Zadlo, Andrzej; Urbanska, Krystyna (26 June 2019). "Melanin presence inhibits melanoma cell spread in mice in a unique mechanical fashion". Scientific Reports. 9 (1): 9280. Bibcode:2019NatSR...9.9280S. doi:10.1038/s41598-019-45643-9. ISSN 2045-2322. PMC 6594928. PMID 31243305.
외부 링크
- "Absorption spectrum of melanin". Department of Computer Science and Technology.
- "Tyrosine metabolism—Reference pathway". Kyoto Encyclopedia of Genes and Genomes.
- "Melanogenesis—Reference pathway". Kyoto Encyclopedia of Genes and Genomes.



