코클레아과
Cochlea| 코클레아과 | |
|---|---|
 달팽이관 단면 | |
달팽이관이 보이는 내이의 일부 | |
| 세부 사항 | |
| 발음 | /kkkli, kokli/[1] |
| 일부 | 내이 |
| 시스템. | 청각계 |
| 식별자 | |
| 라틴어 | 코클레아과 |
| 메쉬 | D003051 |
| NeuroLex ID | birnlex_1190 |
| TA98 | A15.3.03.025 |
| TA2 | 6964 |
| FMA | 60201 |
| 해부학 용어 | |

 |
| 이 기사는 의 구조를 문서화한 시리즈 중 하나입니다. |
| 사람의 귀 |
|---|

달팽이관은 청각에 관여하는 내이의 일부분이다.이것은 골미로의 나선형 공동으로, 인간의 경우 축인 [2][3]모디올러스를 중심으로 2.75바퀴를 돈다.달팽이관의 핵심 요소는 코티 기관으로 청각 기관이며, 코티 기관은 달팽이관의 코일 테이퍼 튜브에서 유체 챔버를 분리하는 칸막이를 따라 분포되어 있습니다.
달팽이 껍데기라는 이름은 고대 그리스어 '코클리아스'에서 유래했다.
구조.

달팽이관(복수는 달팽이관)은 나선형의 속이 빈 원추형 뼈의 방으로, 파동이 기저부(중이와 타원형 창 근처)에서 꼭대기(나선의 꼭대기 또는 중심)로 전파됩니다.달팽이관의 나선관은 내이의 골미로의 한 부분으로 길이가 약 30mm이고 2개를 만든다.모디루스에 대해서 3/4바퀴 돌아요.달팽이관 구조에는 다음이 포함됩니다.
- 3개의 스칼라 또는 챔버:
- 고막과 전정관이 합쳐지는 위치인 헬리코트렘은 달팽이관 꼭대기에서
- 전정관과 달팽이관을 분리하는 레이스너막
- 달팽이관과 고막을 분리하는 주요 구조 요소인 삼투압 나선형 라미나
- 기저막은 달팽이관과 고막을 분리하고 달팽이관 구획의 기계적 파동 전파 특성을 결정하는 주요 구조 요소이다.
- 코티 기관, 감각 상피, 기저막의 세포층, 감각 모세포가 주변 림프와 내림프 사이의 전위차에 의해 작동되는
- 스테레오실리아라고 하는, 코티 기관의 감각 세포
- 나선 인대.
달팽이관은 달팽이 껍질처럼 생긴 내이의 일부입니다.[4]달팽이관은 진동의 형태로 소리를 받고, 이것은 입체성애의 움직임을 일으킨다.그리고 나서 스테레오실리아는 이러한 진동을 해석하기 위해 뇌로 전달되는 신경 자극으로 변환합니다.세 개의 유체 부분 중 두 개는 관이고 세 번째는 청각 신경을 따라 뇌로 이동하는 압력 자극을 감지하는 '코티 기관'이다.이 두 개의 운하는 전정관과 고막관이라고 불린다.
미세해부술
중공 달팽이관의 벽은 얇고 섬세한 상피 조직의 안감으로 뼈로 만들어져 있습니다.이 코일형 튜브는 내부 막 칸막이에 의해 대부분의 길이에 걸쳐 분할됩니다.이 분할막으로 두 개의 유체 충전 외부 공간(덕트 또는 스칼라)이 형성됩니다.달팽이 껍데기 모양의 코일 튜브의 상부에 유체의 방향이 역전되어 전정관이 고관으로 변화한다.이 부위를 헬리코트레마라고 합니다.헬리코트레마에서 이러한 유체가 계속되면 타원형 창에 의해 전정관 안으로 밀려드는 유체가 고막관의 움직임과 둥근 창의 처짐을 통해 다시 이동할 수 있습니다. 유체는 거의 압축할 수 없고 골격 벽은 강하기 때문에 보존된 유체의 양이 어딘가로 빠져나가는 것이 중요합니다.
대부분의 달팽이관을 나누는 세로 칸막이는 그 자체로 세 번째 '덕트'인 유체 충전 튜브입니다.이 중앙 기둥은 달팽이관이라고 불립니다.그것의 액체인 내림프 또한 전해질과 단백질을 포함하고 있지만, 화학적으로 페리림프와는 상당히 다릅니다.페리림프에는 나트륨 이온이 풍부한 반면, 내림프에는 이온성, 전위를 생성하는 칼륨 이온이 풍부합니다.
머리카락 세포는 코티 기관 안에 달팽이관 코일의 전체 길이를 따라 4열로 배열되어 있습니다.3열은 외부모세포(OHC)로 구성되고 1열은 내부모세포(IHC)로 구성됩니다.내부 털 세포는 달팽이관의 주요 신경 출력을 제공합니다.대신에, 바깥의 머리카락 세포는 주로 뇌로부터 신경 입력을 '수신'하는데, 이것은 달팽이관의 기계적 "전 증폭기"의 일부로서 그들의 운동성에 영향을 미칩니다.OHC에 대한 입력은 안쪽의 올리보콜리어 번들을 통해 올리보 본체에서 이루어집니다.
달팽이관은 귀 자체만큼이나 복잡합니다.달팽이관은 기저막, 혈관 선조막 및 라이스너막으로 세 면으로 둘러싸여 있습니다.선조 혈관증은 모세혈관과 분비 세포의 풍부한 층입니다; 레이스너의 막은 내림프로부터 내림프를 분리하는 얇은 막입니다; 그리고 기저막은 기계적으로 다소 단단한 막으로, 청력을 위한 수용 기관, 코르티 기관을 지지하고, c의 기계적 파동 전파 특성을 결정합니다.오클리어 시스템
기능.
달팽이관은 타원형 창을 통해 중이에서 오는 진동에 반응하여 움직이는 물 같은 액체인 내림프로 채워져 있습니다.액체가 움직이면서 달팽이관 칸막이는 움직인다; 수천 개의 머리카락 세포는 그들의 입체 성향을 통해 움직임을 감지하고, 그 움직임을 신경 전달 물질을 통해 수천 개의 신경 세포로 전달되는 전기 신호로 변환한다.이러한 일차 청각 뉴런은 신호를 활동 전위라고 알려진 전기 화학적 자극으로 변환하고, 이것은 더 많은 처리를 위해 청각 신경을 따라 뇌간의 구조물로 이동합니다.
청문회
중이의 등골(등골)은 달팽이관 바깥쪽에 있는 타원형 회향(오목창)으로 진동을 전달하고, 이는 전정관( 달팽이관 위쪽 방)의 요정을 진동시킵니다.달팽이관 환경은 유체-막 시스템이고 공기를 통한 것보다 유체-막 파동을 통해 소리를 이동시키는 데 더 많은 압력이 필요하기 때문에 골격은 달팽이관에 음파를 효율적으로 결합하는 데 필수적입니다.고막(드럼)에서 타원창(등골)까지의 면적비를 20감소함으로써 압력증가를 실현한다.압력 = 힘/면적으로서, 공기 중의 원래 음파 압력에서 약 20배의 압력 게인이 발생합니다.이 이득은 임피던스 매칭의 한 형태로 공기를 통해 이동하는 음파를 유체 막 시스템에서 이동하는 음파와 일치시킵니다.
달팽이관의 밑부분에서 각 '덕트'는 중이강을 향해 있는 막질의 간문에서 끝납니다.전정관은 등골의 발판이 있는 타원형 창문에서 끝난다.압력이 골격 체인을 통해 전달되면 발판이 진동합니다.주변 림프의 파동은 발판에서 벗어나 헬리코트렘 쪽으로 이동한다.그 유체파가 덕트를 분리하는 달팽이관 칸막이를 위아래로 움직이기 때문에, 그 파동은 고막관의 주변 림프에 대응하는 대칭 부분을 가지고 있습니다. 고막은 둥근 창문으로 끝납니다. 타원형 창문이 부풀어 오를 때 튀어나옵니다.
전정관의 주변 림프와 달팽이관의 내림프는 기계적으로 단일 덕트로 작용하며 매우 얇은 레이스너의 막에 의해서만 분리되어 있습니다.달팽이관 내 내림프의 진동은 음파 주파수에 따라 타원형 창으로부터 거리를 정점으로 하는 패턴으로 기저막을 대체한다.코르티 기관은 외부 모발 세포로 인해 이러한 진동을 더욱 증폭시킵니다.내모세포는 유체 내의 진동에 의해 변위되어 끝과 연결된 채널을 통해 K+의 유입에 의해 탈분극되어 신경전달물질을 통해 나선신경절의 1차 청각뉴런으로 신호를 보낸다.
코티 기관의 모발 세포는 기저막의 [5]강직성 정도 때문에 달팽이관 내 위치를 통해 특정 음 주파수에 맞춰 조절됩니다.이러한 강직성은 무엇보다도 기저막의 [6]두께와 폭에 기인하며, 달팽이관 길이를 따라 타원형 창문의 시작 부분에서 가장 가까운 강직성입니다. 여기서 등뼈는 고막에서 오는 진동을 도입합니다.강성이 높기 때문에 고주파 진동만으로 기저막을 움직여 모세포를 움직일 수 있습니다.파동이 달팽이관의 꼭대기(헬리코트레마)를 향해 더 멀리 이동할수록 기저막의 강도는 낮아집니다.따라서 낮은 주파수는 튜브를 따라 이동하며, 강성이 감소된 곳에서 가장 쉽게 이동됩니다. 즉, 기저막이 점점 더 강해질수록 파동은 느려지고, 더 잘 반응합니다.주파수를 낮춥니다.또한 포유동물에서는 달팽이관이 감겨 있어 유체가 채워진 [7]코일을 통과할 때 저주파 진동이 강화되는 것으로 나타났습니다.이러한 음향 수신의 공간적 배열을 토노토피라고 합니다.
매우 낮은 주파수(20Hz 미만)의 경우, 파동은 달팽이관의 전체 경로를 따라 전파됩니다. 즉, 전정관 및 고막관으로부터 헬리코트레마까지 차동적으로 위쪽으로 전파됩니다.이 낮은 주파수는 여전히 어느 정도 코르티 기관을 활성화하지만 음높이의 인식을 유도하기에는 너무 낮다.고주파수는 강성 매개 색조술로 인해 헬리코트레마에 전파되지 않습니다.
매우 큰 소음으로 인해 기저막이 매우 강하게 움직이면 머리카락 세포가 죽을 수 있습니다.이는 부분적인 청력 상실의 흔한 원인이며 총기나 중장비 사용자가 귀마개나 귀마개를 자주 착용하는 이유이다.
모세포증폭
달팽이관은 소리를 "수신"할 뿐만 아니라, 건강한 달팽이관은 필요할 때 소리를 생성하고 증폭시킵니다.유기체가 매우 미미한 소리를 들을 수 있는 메커니즘이 필요한 경우, 달팽이관은 OHC의 역변환에 의해 증폭되어 전기 신호를 양의 피드백 구성으로 역변환합니다.OHC는 외부 막에 프레스틴이라고 불리는 단백질 모터가 있습니다; 이것은 유체-막 파동으로 다시 결합되는 추가적인 움직임을 생성합니다.이 "능동 증폭기"는 귀의 [8][9]약한 소리를 증폭시키는 능력에 필수적입니다.
능동 증폭기는 또한 달팽이관에서 중이를 통해 다시 귓구멍으로 음파 진동이 방출되는 현상을 초래한다.
이음 방출
이음향 방출은 타원형 창문을 통해 달팽이관을 통해 나오는 파동이 중이를 통해 다시 고막으로 전파되고, 이 파동은 마이크에 의해 잡힐 수 있습니다.이음향 방출은 달팽이관이 잘 작동할 때 나타나고 OHC 활동 상실을 겪을 때는 덜 발생하기 때문에 청각 장애에 대한 일부 테스트에서 중요하다.
갭 접합부의 역할
달팽이관에서 발현되는 코넥신이라고 불리는 간극 접합 단백질은 청각 [10]기능에 중요한 역할을 합니다.간극 접합 유전자의 돌연변이는 신드롬성 및 비신드롬성 [11]난청의 원인이 되는 것으로 밝혀졌다.코클라에서 발견되는 두 개의 뚜렷한 갭 접합계에는 코넥신 30과 코넥신 26을 포함한 특정 코넥신이 널리 분포한다.상피세포 갭 접합 네트워크는 비감각적 상피세포를 결합하고, 접속조직 갭 접합 네트워크는 접속조직 셀을 결합합니다.간극 접합 채널은 모발 [12]세포에서 기계적 전달 후 칼륨 이온을 내림프까지 재활용합니다.중요한 것은, 간극 접합 채널이 달팽이관 지지 세포 사이에서 발견되지만, 청각 모발 [13]세포 사이에는 발견되지 않는다.
임상적 의의
이 섹션은 확장해야 합니다.추가하시면 됩니다. (2015년 9월) |
난청
바이오닉스
2009년 Massachusetts Institute of Technology의 엔지니어들은 기존 기술에 필요한 전력의 극히 일부만을 사용하면서 매우 광범위한 무선 주파수를 빠르게 분석할 수 있는 전자 칩을 개발했습니다. 그 설계는 특히 달팽이관을 [14][15]모방한 것입니다.
기타 동물
달팽이관의 코일 모양은 포유류에게 독특하다.조류와 다른 비동물 척추동물에서, 청력을 위한 감각 세포를 포함하는 구획은 감겨있지 않음에도 불구하고 때때로 "코클레아"라고도 불립니다.대신, 그것은 달팽이관이라고도 불리는 블라인드 엔드 튜브를 형성합니다.이 차이는 포유동물과 비동물 척추동물 사이의 청각 주파수 범위의 차이와 병행하여 진화한 것으로 보인다.포유류의 우수한 주파수 범위는 부분적으로 외부 털 세포의 활발한 세포-체 진동에 의해 소리의 사전 증폭이라는 독특한 메커니즘 때문이다.그러나 주파수 분해능은 포유류에서 대부분의 도마뱀이나 조류보다 낫지 않지만, 주파수 상한은 때로는 훨씬 높다.대부분의 새들은 4-5kHz 이상의 소리를 듣지 못하는데, 현재 알려진 최대치는 외양간 부엉이에서 11kHz입니다.어떤 해양 포유류들은 200킬로헤르츠까지 듣습니다.짧고 곧은 칸막이보다 긴 코일 칸막이는 추가적인 청력 범위를 위한 더 많은 공간을 제공하고 포유류의 [16]청력과 관련된 고도로 파생된 행동 중 일부를 가능하게 했다.
달팽이관 연구는 기본적으로 모세포 수준에 초점을 맞춰야 하기 때문에 다양한 종의 모세포 간의 해부학적, 생리학적 차이점에 주목하는 것이 중요하다.예를 들어, 새들에게는, 바깥과 안쪽의 털세포 대신에, 키가 크고 짧은 털세포가 있다.이 비교 데이터와 관련하여 몇 가지 유사점이 있다.첫째, 키높은 모발세포는 내부모발세포와 기능이 매우 유사하며, 구심성 청각신경섬유 내부가 결여된 짧은 모발세포는 외부모발세포와 유사하다.하지만 피할 수 없는 한 가지 차이점은 조류에서는 모든 털세포가 구조막에 부착되어 있는 반면 포유동물에서는 외부 털세포만 구조막에 부착되어 있다는 것이다.
역사
달팽이 껍데기라는 이름은 달팽이 껍데기를 뜻하는 라틴어에서 유래했으며, 달팽이 껍데기는 그리스어의 χχςςα kokhlias(snail, snrew)[17]에서 유래했다. 달팽이 껍데기는 모노렘을 제외하고 포유동물에서 감겨 있다.
기타 이미지

오른쪽 삼투압 미로.측면도

우측 삼투압 미로 내부
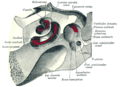
위에서 보면 달팽이관과 전정입니다.
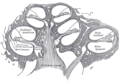
달팽이관의 단면입니다.




「 」를 참조해 주세요.
레퍼런스
- ^ "cochlea". Dictionary.com Unabridged (Online). n.d.
- ^ Anne M. Gilroy; Brian R. MacPherson; Lawrence M. Ross (2008). Atlas of anatomy. Thieme. p. 536. ISBN 978-1-60406-151-2.
- ^ Moore & Dalley (1999). Clinically Oriented Anatomy (4th ed.). p. 974. ISBN 0-683-06141-0.
- ^ The Kingfisher children's encyclopedia. Kingfisher Publications. (3rd ed., fully rev. and updated ed.). New York: Kingfisher. 2012 [2011]. ISBN 9780753468142. OCLC 796083112.
{{cite book}}: CS1 유지보수: 기타 (링크) - ^ Guenter Ehret (Dec 1978). "Stiffness gradient along the basilar membrane as a way for spatial frequency analysis within the cochlea" (PDF). J Acoust Soc Am. 64 (6): 1723–6. doi:10.1121/1.382153. PMID 739099.
- ^ Camhi, J. Neuroethology: 신경세포와 동물의 자연스러운 행동.시나우어 어소시에이츠, 1984.
- ^ Manoussaki D, Chadwick RS, Ketten DR, Arruda J, Dimitriadis EK, O'Malley JT (2008). "The influence of cochlear shape on low-frequency hearing". Proc Natl Acad Sci U S A. 105 (16): 6162–6166. Bibcode:2008PNAS..105.6162M. doi:10.1073/pnas.0710037105. PMC 2299218. PMID 18413615.
- ^ Ashmore, Jonathan Felix (1987). "A fast motile response in guinea-pig outer hair cells: the cellular basis of the cochlear amplifier". The Journal of Physiology. 388 (1): 323–347. doi:10.1113/jphysiol.1987.sp016617. ISSN 1469-7793. PMC 1192551. PMID 3656195.

- ^ Ashmore, Jonathan (2008). "Cochlear Outer Hair Cell Motility". Physiological Reviews. 88 (1): 173–210. doi:10.1152/physrev.00044.2006. ISSN 0031-9333. PMID 18195086. S2CID 17722638.
- ^ Zhao, H. -B.; Kikuchi, T.; Ngezahayo, A.; White, T. W. (2006). "Gap Junctions and Cochlear Homeostasis". Journal of Membrane Biology. 209 (2–3): 177–186. doi:10.1007/s00232-005-0832-x. PMC 1609193. PMID 16773501.
- ^ Erbe, C. B.; Harris, K. C.; Runge-Samuelson, C. L.; Flanary, V. A.; Wackym, P. A. (2004). "Connexin 26 and Connexin 30 Mutations in Children with Nonsyndromic Hearing Loss". The Laryngoscope. 114 (4): 607–611. doi:10.1097/00005537-200404000-00003. PMID 15064611. S2CID 25847431.
- ^ Kikuchi, T.; Kimura, R. S.; Paul, D. L.; Takasaka, T.; Adams, J. C. (2000). "Gap junction systems in the mammalian cochlea". Brain Research. Brain Research Reviews. 32 (1): 163–166. doi:10.1016/S0165-0173(99)00076-4. PMID 10751665. S2CID 11292387.
- ^ Kikuchi, T.; Kimura, R. S.; Paul, D. L.; Adams, J. C. (1995). "Gap junctions in the rat cochlea: Immunohistochemical and ultrastructural analysis". Anatomy and Embryology. 191 (2): 101–118. doi:10.1007/BF00186783. PMID 7726389. S2CID 24900775.
- ^ Anne Trafton (June 3, 2009). "Drawing inspiration from nature to build a better radio: New radio chip mimics human ear, could enable universal radio". MIT newsoffice.
- ^ Soumyajit Mandal; Serhii M. Zhak; Rahul Sarpeshkar (June 2009). "A Bio-Inspired Active Radio-Frequency Silicon Cochlea" (PDF). IEEE Journal of Solid-State Circuits. 44 (6): 1814–1828. Bibcode:2009IJSSC..44.1814M. doi:10.1109/JSSC.2009.2020465. hdl:1721.1/59982. S2CID 10756707.
- ^ Vater M, Meng J, Fox RC청각 기관의 진화 및 전문화: 초기 및 후기 포유동물.입력: GA Manley, AN Popper, RR Fay(Eds).척추동물 청각 시스템의 진화(Springer-Verlag, New York, 2004), 페이지 256–288.
- ^ 어원 "sylleaa",
추가 정보
- Dallos, Peter; Popper, Arthur N.; Fay, Richard R. The Cochlea.
- Imbert, Michel; Kay, R. H. (1992). Audition. ISBN 9780262023313.
- Jahn, Anthony F.; Santos-Sacchi, Joseph (2001). Physiology of the Ear. ISBN 9781565939943.
- Roeser, Ross J.; Valente, Michael; Hosford-Dunn, Holly (2007). Audiology. ISBN 9781588905420.
외부 링크
- 미국 국립 의학 도서관(MeSH)의 Cochlea
- R의 "Promenade 'Around the Cochlea"몽펠리에 대학교 푸졸, S. Blatrix, T. Pujol 등
- '귀의 역사 비디오'


