채널 차단기
Channel blocker
채널차단기는 특정 분자가 세포 내에서 생리적 반응을 일으키기 위해 이온 채널의 개방을 막기 위해 사용하는 생물학적 메커니즘이다. 채널 차단은 양이온, 음이온, 아미노산, 그리고 다른 화학물질과 같은 다른 종류의 분자에 의해 수행된다. 이러한 차단기는 이온 채널 길항제 역할을 하여 통상적으로 채널 개방에 의해 제공되는 반응을 방지한다.
이온 채널은 모공 역할을 하는 단백질을 활용해 세포막을 통한 이온의 선택적 통행을 허용해 전하가 세포 안팎으로 통할 수 있도록 했다.[1] 이러한 이온 채널은 가장 흔히 게이트로 되어 있는데, 이는 채널이 열리고 닫히도록 하기 위해 특정한 자극을 필요로 한다는 것을 의미한다. 이러한 이온 채널 유형은 전하를 띤 이온의 흐름을 막 전체에 걸쳐 조절하여 세포의 막 전위를 조정한다.
약리학 분야에서 채널 차단제 역할을 하는 분자는 약리학 분야에서 중요한데, 약물 설계의 많은 부분이 생리학적 반응을 조절하는 데 이온 채널 길항제의 사용이기 때문이다. 특정 채널의 채널 블록 분자의 특수성은 수많은 장애를 치료하는 데 귀중한 도구가 된다.[2][3]
배경
이온 채널
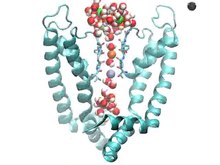
채널 차단기의 메커니즘을 이해하기 위해서는 이온 채널의 구성을 이해하는 것이 중요하다. 그들의 주요 기능은 세포막을 통한 이온의 흐름을 통해 세포의 휴식막 전위에 기여하는 것이다. 이온들은 이 임무를 수행하기 위해 불리한 과정인 지질 빌라이어 막의 소수성 영역을 횡단할 수 있어야 한다. 이온 수송을 돕기 위해 이온 채널은 막부를 통해 친수성 기공을 형성하여 일반적으로 불리한 친수성 분자의 전달을 허용한다.[4] 다양한 이온 채널은 다양한 기능 메커니즘을 가지고 있다. 여기에는 다음이 포함된다.
- 전압 게이트 이온 채널
- 멤브레인 전위 변화에 의해 활성화된 이온도
- 리간드 게이트 이온 채널
- 채널 단백질에 대한 작은 분자의 결합에 의해 매개되는 이온 채널
- 기계 감도가 높은 이온 채널
- 스트레치, 진동 또는 온도 변화에 반응하는 이온 채널
- 경량 이온 채널
- 빛에 반응하여 열거나 닫히는 이온 채널
이온 채널 차단기 역할을 하는 분자는 이러한 다양한 채널 중 어느 것과도 관련지어 사용될 수 있다. 예를 들어, 작용 전위의 생산에 필수적인 나트륨 채널은 많은 다른 독소의 영향을 받는다. 복어에서 발견된 독소인 테트로도톡신(TTX)은 채널의 선택성 필터 영역을 차단해 나트륨 이온 수송을 완전히 차단한다.[5] 이온 채널의 모공 구조 중 많은 부분이 채널 기능을 억제하기 위해 독소를 사용한 연구에서 해명되었다.[6][7][8]
아이덴티티
X선 결정학 및 전기생리학과 같은 도구는 개방 채널 블록 분자의 결합 부위를 찾는데 필수적이었다. 이온 채널의 생물학적, 화학적 구성을 연구함으로써, 연구자들은 특정 지역에 결합하는 분자의 구성을 결정할 수 있다. X선 결정학은 해당 채널과 분자의 구조적 이미지를 제공한다.[9] 또한 친수성 플롯을 통해 채널 영역의 친수성을 결정하는 것은 분자의 화학적 구성과 그것이 특정 지역에 결합하는 이유에 대한 단서를 제공한다. 예를 들어, 단백질이 채널의 소수성 영역에 결합하는 경우(따라서, 트랜스메브레인 영역이 있는 경우), 해당 분자는 아미노산 알라닌, 류신 또는 페닐알라닌으로 구성될 수 있는데, 이는 모두 그 자체로 소수성이기 때문이다.[10] 개방 채널 블록 분자의 억제 작용을 이해하는 데 채널 활성화를 이끄는 이온적 요인을 분석하는 것이 중요할 수 있기 때문에 전기생리학은 채널 구조를 파악하는 데도 중요한 도구다.[3][9]
생리학

수용체 길항제
채널 차단기는 각 이온 채널의 길항제다. 많은 채널은 세포와 유기체 내의 요건에 따라 정상 기능을 촉진하거나 억제할 수 있는 규제 요소에 대한 결합 지점을 가지고 있다. 작용제 결합의 정상적인 기능은 다양한 다운스트림 효과로 이어지는 세포 변화의 발생이다. 이러한 영향은 막 전위를 바꾸는 것에서부터 신호 계단식 시작에 이르기까지 다양하다.[11] 반대로 개방 채널 차단기가 셀에 결합할 때 작용제 결합의 정상적인 기능을 방지한다. 예를 들어, 전압 게이트 채널은 멤브레인 전위에 기초하여 개폐되며, 이온의 허용량에 의한 작용 전위 생성에 있어 확립된 구배 아래로 흐른다. 그러나 개방 채널 차단기는 이온이 흐르지 않도록 이들 채널에 결합할 수 있으므로 작용 전위의 시작을 억제한다.[12]
분자의 특수성
많은 다른 유기 화합물들이 채널의 특수성에도 불구하고 채널 차단제의 역할을 할 수 있다. 채널은 지역에 걸쳐 있는 막 때문에 다양한 이온이나 화합물을 구별할 수 있는 구조로 진화했다. 예를 들어, 일부 물체는 나트륨 채널에 맞추려고 시도하는 칼륨 이온과 같이 작은 물체를 운반하기 위해 구조적으로 지정된 채널에 맞추기에는 너무 크다. 반대로, 어떤 물체는 너무 작아서 칼륨 채널을 통과하려고 하는 나트륨 이온과 같이 특정 채널 모공에 의해 적절하게 안정화되지 않는다.[11][13] 두 경우 모두 채널 플럭스는 허용되지 않는다. 그러나 특정 화합물이 채널에 대한 적절한 화학적 친화력을 갖는 한, 해당 화합물은 채널 모공을 결합하고 차단할 수 있을 것이다. 예를 들어 TTX가 나트륨 이온과 화학적으로 훨씬 크고 다르다는 사실에도 불구하고 TTX는 전압 게이트 나트륨 채널을 결합하고 비활성화할 수 있다. TTX와 나트륨 이온 사이의 크기와 화학적 성질의 차이를 고려할 때, 이것은 구조물이 보통 특정 채널을 차단하기 위해 사용되는 예다. [14]
키네틱스
채널 블록은 대상 채널의 모공 일부에 결합할 수 있는 한 많은 다른 유형의 유기 화합물에 의해 유도될 수 있다. 채널 차단기의 동역학은 주로 마취제로 사용되지만 이해된다. 국소마취제는 대상 뉴런에서 페이식 블록 상태를 유도하여 작용한다.[13] 처음에는 개방 채널 차단기가 차단되는 채널이 거의 없고 그 특성에 따라 차단기 자체를 채널에서 빠르게 또는 천천히 방출할 수 있기 때문에 액션 전위를 효과적으로 차단하지 못한다. 그러나 파상블록은 반복적인 탈분극화가 뉴런의 채널에 대한 차단기의 친화력을 증가시키면서 발생한다. 이용 가능한 채널의 증가와 차단기 결합 친화력을 높이기 위한 채널 순응의 변경의 조합이 이 작용에 책임이 있다.[13][15][16]
임상적 유의성
치료 용도
칼슘 의존성 신경독성을 중재하기 위한 NMDA 수용체 활성화가 과도한 것과 다양한 신경퇴행성 질환이 연관되어 왔다. 연구원들은 많은 다양한 NMDA 길항제들과 그들의 치료 효능을 조사했는데, 그 중 어느 것도 안전하면서도 효과적이라고 결론지지는 않았다.[17] 수년 동안 연구원들은 신경독성의 치료 옵션으로 개방 채널 블록인 메만틴의 효과를 연구해 왔다. 그들은 메만틴의 더 빠른 차단과 차단 해제율, 그리고 전반적인 운동학적 특성이 임상적 허용오차의 근본적인 이유가 될 수 있다고 가설을 세웠다.[17][3] 경쟁력 없는 대항마로써, memantine은 높은 글루탐산염 농도에도 불구하고 NMDA 수준을 정상 수준에 가깝게 해야 한다. 연구자들은 이 정보를 바탕으로 언젠가 메망틴이 다른 치료법에 비해 부작용이 거의 없는 신경독성 관련 글루탐산염 수치가 증가하는 것을 막기 위해 개방 채널 블록으로 사용될 수 있을 것으로 추측했다.[17]
알츠하이머병
특정 신경퇴행성 장애인 알츠하이머병은 알츠하이머병의 주요 인지증상을 초래한다고 여겨지는 글루타민성 신경전달 방해와 관련이 있다.[18][2][3] 연구원들은 비경쟁 NMDA 수용체 작용제를 사용하여 심각한 부작용을 일으키지 않고 이러한 증상들을 관리하는 데 도움을 줄 수 있다고 제안한다.[18] 알츠하이머 치료를 위해 승인된 유일한 약품 중 하나로, 메망틴은 흥분성 후 시냅스 전류가 영향을 받지 않고 유지되도록 하는 동시에 억제 후 시냅스 전류의 발생률과 진폭을 감소시키는 것으로 나타났다.[19] 증거는 강한 전압 의존성과 메만틴의 빠른 운동 모두 부작용 감소와 인지 진행의 원인이 될 수 있다는 가설을 뒷받침한다.[20]
낭포성 섬유증
낭포성 섬유증은 진행성 유전성 질환으로 CFTR(Convertembrane Regulator, CFTR) 기능장애와 관련이 있다.[21] 특정 세포질, 음전하 물질에 의해 이 채널이 막히면 염화 이온과 중탄산염 음이온 수송이 감소하고 유체와 염분 분비가 감소한다. 이것은 낭포성 섬유증의 특징인 두꺼운 점액질의 축적을 초래한다.[21]
약리학
마취제
마취제 분야에서는 채널 차단기가 필수적이다. 나트륨 채널 억제제는 환자의 고출혈성 조직을 억제할 수 있기 때문에 항피질제와 부정맥 모두로 사용된다.[22] 특정 나트륨 채널 차단기를 조직에 도입하면 나트륨 채널에 대한 차단기의 우선 결합이 가능해져 조직으로 유입되는 나트륨의 흐름을 궁극적으로 억제할 수 있다. 시간이 지남에 따라, 이 메커니즘은 전체적인 조직 흥분 감소로 이어진다. 장시간 과극화는 정상적인 채널 복구를 방해하고 지속적인 억제를 허용하여 주어진 설정에서 마취제의 동적 제어를 제공한다.[22]
알츠하이머병
글루탐산염에 과도하게 노출되면 알츠하이머병 환자에게 신경독성이 생긴다. 구체적으로는 NMDA형 글루탐산염 수용체의 과잉 활성화가 신경세포의 흥분성 및 세포사멸과 연계되어 왔다.[18][2] 이에 대한 잠재적 해결책은 임상 부작용을 일으킬 정도로 급격한 간섭 없이 NMDA 수용체 활성의 감소다.[23]
더 이상의 신경퇴행 발생을 막기 위한 시도로 연구자들은 개방형 채널 블록인 메만틴을 치료의 한 형태로 사용해 왔다. 지금까지 알츠하이머 병에 걸린 환자에게 메만틴을 사용하는 것은 많은 다른 증상들에 걸쳐 임상적인 진보를 빠르게 가져온다. 메만틴은 채널 내 축적을 방지하고 정상적인 시냅스 전송이 가능한 키네틱스를 빠르게 수정하는 능력이 있어 효과가 있다고 판단된다. 다른 채널 차단기가 모든 NMDA 수용체 활동을 차단하는 것으로 밝혀져 임상적 부작용을 낳고 있다.[3]
CFTR 채널 기능 저하
염화물이온, 중탄산염 음이온 및 유체 수송에서 낭포성 섬유증 투과 조절기(CFTRs)[24] 기능. 그것들은 주로 호흡기, 췌장, 위장, 생식 조직에서 상피 세포의 세포막에서 표현된다.[21][24] 비정상적으로 격리된 CFTR 기능은 과도한 유체 분비를 초래한다. CFTR-172와inh 글리H-101과 같은 고선호도 CFTR 억제제는 분비성 설사 꿩의 치료에 효율적인 것으로 나타났다.[25][26] 이론적으로 CFTR 채널 차단기는 남성 피임약으로도 유용할 수 있다. CFTR 채널은 정자 용량 할당에 필수적인 중탄산염 음이온 입력을 중재한다.[27]
다양한 종류의 물질이 CFTR 염화물이온 채널을 차단하는 것으로 알려져 왔다. 가장 잘 알려져 있고 연구된 물질로는 설포닐루레아, 아릴라미노벤제노아테스, 이황성 스틸베네스가 있다.[28][29][30] 이러한 차단제는 세포질 쪽에서 독점적으로 모공에 들어갈 때 측면에 의존하며, 극지방 막 전위로서 전압에 의존하며, 세포질 쪽에서 모공으로의 부정충전 물질 진입을 선호하며, 염화물이온 농도에 의존하여 높은 세포외 염화물이온을 정전기적으로 밀어낸다.차단제를 다시 세포질 속으로 쑤셔넣었다.[31]
종류들
채널 차단기의 주요 등급은 다음과 같이 몇 가지가 있다.
모공 결합을 통해 리간드 게이트 이온 채널(LGICs)에 작용하는 다음과 같은 유형도 존재한다.
채널 차단기는 또한 AMPA 수용체, 글리신 수용체, 카이네이트 수용체, P2X 수용체, 아연(Zn2+) 활성 채널에서도 작용하는 것으로 알려져 있다. 채널 차단기에 의해 매개되는 억제 유형을 비경쟁적 또는 비경쟁적이라 할 수 있다.
참고 항목
참조
- ^ "Medical Definition of Ion channel". MedicineNet. Retrieved 2017-03-20.
- ^ a b c Kocahan S, Doğan Z (February 2017). "Mechanisms of Alzheimer's Disease Pathogenesis and Prevention: The Brain, Neural Pathology, N-methyl-D-aspartate Receptors, Tau Protein and Other Risk Factors". Clinical Psychopharmacology and Neuroscience. 15 (1): 1–8. doi:10.9758/cpn.2017.15.1.1. PMC 5290713. PMID 28138104.
- ^ a b c d e Lipton SA (January 2004). "Failures and successes of NMDA receptor antagonists: molecular basis for the use of open-channel blockers like memantine in the treatment of acute and chronic neurologic insults". NeuroRx. Neuroprotection. 1 (1): 101–10. doi:10.1602/neurorx.1.1.101. PMC 534915. PMID 15717010.
- ^ Ahern CA, Payandeh J, Bosmans F, Chanda B (January 2016). "The hitchhiker's guide to the voltage-gated sodium channel galaxy". The Journal of General Physiology. 147 (1): 1–24. doi:10.1085/jgp.201511492. PMC 4692491. PMID 26712848.
- ^ Moore JW, Blaustein MP, Anderson NC, Narahashi T (May 1967). "Basis of tetrodotoxin's selectivity in blockage of squid axons". The Journal of General Physiology. 50 (5): 1401–11. doi:10.1085/jgp.50.5.1401. PMC 2225715. PMID 6033592.
- ^ Stevens M, Peigneur S, Tytgat J (2011-11-09). "Neurotoxins and their binding areas on voltage-gated sodium channels". Frontiers in Pharmacology. 2: 71. doi:10.3389/fphar.2011.00071. PMC 3210964. PMID 22084632.
- ^ Miller C (December 1988). "Competition for block of a Ca2(+)-activated K+ channel by charybdotoxin and tetraethylammonium". Neuron. 1 (10): 1003–6. doi:10.1016/0896-6273(88)90157-2. PMID 2483092.
- ^ Aiyar J, Withka JM, Rizzi JP, Singleton DH, Andrews GC, Lin W, Boyd J, Hanson DC, Simon M, Dethlefs B (November 1995). "Topology of the pore-region of a K+ channel revealed by the NMR-derived structures of scorpion toxins". Neuron. 15 (5): 1169–81. doi:10.1016/0896-6273(95)90104-3. PMID 7576659.
- ^ a b Findeisen F, Campiglio M, Jo H, Abderemane-Ali F, Rumpf CH, Pope L, Rossen ND, Flucher BE, DeGrado WF, Minor DL (March 2017). "Stapled Voltage-Gated Calcium Channel (CaV) α-Interaction Domain (AID) Peptides Act As Selective Protein-Protein Interaction Inhibitors of CaV Function". ACS Chemical Neuroscience. 8: 1313–1326. doi:10.1021/acschemneuro.6b00454. PMC 5481814. PMID 28278376.
- ^ Phoenix DA, Harris F (2002-01-01). "The hydrophobic moment and its use in the classification of amphiphilic structures (review)". Molecular Membrane Biology. 19 (1): 1–10. doi:10.1080/09687680110103631. PMID 11989818.
- ^ a b Jackson MB (February 2010). "Open channel block and beyond". The Journal of Physiology. 588 (Pt 4): 553–4. doi:10.1113/jphysiol.2009.183210. PMC 2828128. PMID 20173077.
- ^ Ahern CA, Payandeh J, Bosmans F, Chanda B (January 2016). "The hitchhiker's guide to the voltage-gated sodium channel galaxy". The Journal of General Physiology. 147 (1): 1–24. doi:10.1085/jgp.201511492. PMC 4692491. PMID 26712848.
- ^ a b c Butterworth JF, Strichartz GR (April 1990). "Molecular mechanisms of local anesthesia: a review". Anesthesiology. 72 (4): 711–34. doi:10.1097/00000542-199004000-00022. PMID 2157353.
- ^ Evans MH (September 1969). "Mechanism of saxitoxin and tetrodotoxin poisoning". British Medical Bulletin. 25 (3): 263–7. doi:10.1093/oxfordjournals.bmb.a070715. PMID 5812102.
- ^ Mert T, Gunes Y, Guven M, Gunay I, Ozcengiz D (March 2002). "Comparison of nerve conduction blocks by an opioid and a local anesthetic". European Journal of Pharmacology. 439 (1–3): 77–81. doi:10.1016/S0014-2999(02)01368-7. PMID 11937095.
- ^ Mitolo-Chieppa D, Carratu MR (May 1983). "Anaesthetic drugs: electrophysiological bases of their conduction blocking effect". Pharmacological Research Communications. 15 (5): 439–50. doi:10.1016/s0031-6989(83)80064-2. PMID 6351107.
- ^ a b c Chen HS, Pellegrini JW, Aggarwal SK, Lei SZ, Warach S, Jensen FE, Lipton SA (November 1992). "Open-channel block of N-methyl-D-aspartate (NMDA) responses by memantine: therapeutic advantage against NMDA receptor-mediated neurotoxicity". The Journal of Neuroscience. 12 (11): 4427–36. PMID 1432103.
- ^ a b c Müller WE, Mutschler E, Riederer P (July 1995). "Noncompetitive NMDA receptor antagonists with fast open-channel blocking kinetics and strong voltage-dependency as potential therapeutic agents for Alzheimer's dementia". Pharmacopsychiatry. 28 (4): 113–24. doi:10.1055/s-2007-979603. PMID 7491365.
- ^ Povysheva NV, Johnson JW (December 2016). "Effects of memantine on the excitation-inhibition balance in prefrontal cortex". Neurobiology of Disease. 96: 75–83. doi:10.1016/j.nbd.2016.08.006. PMC 5102806. PMID 27546057.
- ^ Dominguez, Evangelyn; Chin, Ting-Yu; Chen, Chih-Ping; Wu, Tzong-Yuan (2011-12-01). "Management of moderate to severe Alzheimer's disease: Focus on memantine". Taiwanese Journal of Obstetrics and Gynecology. 50 (4): 415–423. doi:10.1016/j.tjog.2011.10.004. ISSN 1028-4559. PMID 22212311.
- ^ a b c Lubamba B, Dhooghe B, Noel S, Leal T (October 2012). "Cystic fibrosis: insight into CFTR pathophysiology and pharmacotherapy". Clinical Biochemistry. 45 (15): 1132–44. doi:10.1016/j.clinbiochem.2012.05.034. PMID 22698459.
- ^ a b Ramos E, O'leary ME (October 2004). "State-dependent trapping of flecainide in the cardiac sodium channel". The Journal of Physiology. 560 (Pt 1): 37–49. doi:10.1113/jphysiol.2004.065003. PMC 1665201. PMID 15272045.
- ^ Lipton SA (May 2007). "Pathologically-activated therapeutics for neuroprotection: mechanism of NMDA receptor block by memantine and S-nitrosylation". Current Drug Targets. 8 (5): 621–32. doi:10.2174/138945007780618472. PMID 17504105.
- ^ a b Frizzell RA, Hanrahan JW (June 2012). "Physiology of epithelial chloride and fluid secretion". Cold Spring Harbor Perspectives in Medicine. 2 (6): a009563. doi:10.1101/cshperspect.a009563. PMC 3367533. PMID 22675668.
- ^ Muanprasat C, Sonawane ND, Salinas D, Taddei A, Galietta LJ, Verkman AS (August 2004). "Discovery of glycine hydrazide pore-occluding CFTR inhibitors: mechanism, structure-activity analysis, and in vivo efficacy". The Journal of General Physiology. 124 (2): 125–37. doi:10.1085/jgp.200409059. PMC 2229623. PMID 15277574.
- ^ Ma T, Thiagarajah JR, Yang H, Sonawane ND, Folli C, Galietta LJ, Verkman AS (December 2002). "Thiazolidinone CFTR inhibitor identified by high-throughput screening blocks cholera toxin-induced intestinal fluid secretion". The Journal of Clinical Investigation. 110 (11): 1651–8. doi:10.1172/JCI16112. PMC 151633. PMID 12464670.
- ^ Chen H, Ruan YC, Xu WM, Chen J, Chan HC (2012-11-01). "Regulation of male fertility by CFTR and implications in male infertility". Human Reproduction Update. 18 (6): 703–13. doi:10.1093/humupd/dms027. PMID 22709980.
- ^ Schultz BD, DeRoos AD, Venglarik CJ, Singh AK, Frizzell RA, Bridges RJ (August 1996). "Glibenclamide blockade of CFTR chloride channels". The American Journal of Physiology. 271 (2 Pt 1): L192-200. doi:10.1152/ajplung.1996.271.2.L192. PMID 8770056.
- ^ Zhang ZR, Zeltwanger S, McCarty NA (May 2000). "Direct comparison of NPPB and DPC as probes of CFTR expressed in Xenopus oocytes". The Journal of Membrane Biology. 175 (1): 35–52. doi:10.1007/s002320001053. PMID 10811966.
- ^ Linsdell P, Hanrahan JW (November 1996). "Disulphonic stilbene block of cystic fibrosis transmembrane conductance regulator Cl- channels expressed in a mammalian cell line and its regulation by a critical pore residue". The Journal of Physiology. 496 (Pt 3): 687–93. doi:10.1113/jphysiol.1996.sp021719. PMC 1160856. PMID 8930836.
- ^ Linsdell P (February 2014). "Cystic fibrosis transmembrane conductance regulator chloride channel blockers: Pharmacological, biophysical and physiological relevance". World Journal of Biological Chemistry. 5 (1): 26–39. doi:10.4331/wjbc.v5.i1.26. PMC 3942540. PMID 24600512.


