해양 원핵생물
Marine prokaryotes.jpg)
| 다음에 대한 일련의 개요의 일부 |
| 해양생물 |
|---|
| |
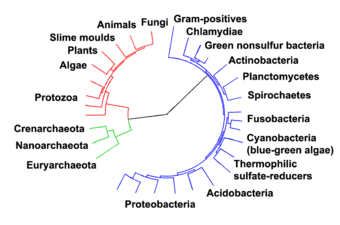
해양 원핵생물들은 해양 박테리아와 해양 고고학이다. 그들은 서식지에 의해 해양 환경, 즉 바다나 바다의 소금물이나 해안 강초의 고사리 같은 곳에서 사는 원핵생물로 정의된다. 모든 세포 생물 형태는 원핵생물과 진핵생물로 나눌 수 있다. 진핵생물은 세포가 세포막 안에 밀폐된 핵을 가지고 있는 유기체인 반면 원핵생물은 세포막 안에 밀폐된 핵을 가지고 있지 않은 유기체들이다.[1][2][3] 생물을 분류하는 3개 영역 체계는 또 다른 분열을 더한다: 원핵생물은 생물의 2개 영역으로 나뉘는데, 원핵생물은 현미경 박테리아와 현미경 고고학이고, 다른 모든 영역인 진핵생물은 제3의 영역이 된다.[4]
원핵생물은 분해자들이 영양분을 재활용하는 생태계에서 중요한 역할을 한다. 일부 원핵생물들은 병원성으로 인해 식물과 동물에서 질병과 심지어 죽음을 야기한다.[5] 해양 원핵생물들은 탄소나 다른 영양소의 중요한 순환뿐만 아니라 바다에서 일어나는 광합성의 상당한 수준에 책임이 있다.[6]
원핵생물들은 생물권 전체에 서식한다. 2018년에 지구상의 모든 원핵생물들의 총 바이오매스는 770억 톤의 탄소(77 Gt C)에 해당한다고 추정되었다. 이것은 고대의 경우 7 Gt C, 박테리아의 경우 70 Gt C로 구성되어 있다. 이러한 수치는 지구상 동물들의 총 바이오매스 추정치인 약 2 Gt C와 인간의 총 바이오매스 0.06 Gt C와 대조될 수 있다.[7] 이는 고고학이 집단적으로 인간의 100배 이상, 1000배 이상의 박테리아를 가지고 있다는 것을 의미한다.
지구상의 생명체가 존재한 첫 6억년 동안 지구상에 존재한다는 명확한 증거는 없다. 생명체가 도착했을 때, 그것은 해양 원핵생물들에 의해 3억 2천만 년 동안 지배되었다. 크라운 eukaryotes의 형태로, 더 복잡한 삶은 단지 5억년 전에 캄브리아기가 폭발하기 전까지는 나타나지 않았다.[8]
진화
−4500 — – −4000 — – −3500 — – −3000 — – −2500 — – −2000 — – −1500 — – −1000 — – −500 — – 0 — | 해양의 원핵생물 |
| ||||||||||||||||||||||||
지구는 약 45억 4천만 년 전이다.[9][10][11] 지구상에 생명체가 존재한다는 최초의 확실한 증거는 일찍이 녹아버린 하데스 연에 이어 지질학적 지각까지 굳어지기 시작한 이후 적어도 35억년 전 [12][13]어안시대 때부터 시작되었다. 미생물 돗자리 화석이 호주 서부 34억8000만년 된 사암에서 발견됐다.[14][15]

과거 종들도 진화 역사에 대한 기록을 남겼다. 화석은 오늘날의 유기체의 비교 해부학과 함께 형태학적, 즉 해부학적 기록을 구성한다.[16] 고생물학자들은 현대와 멸종된 종 모두의 해부학을 비교함으로써 그 종들의 행을 유추할 수 있다. 그러나 이 접근법은 껍질, 뼈, 치아 등 딱딱한 신체 부위를 가진 유기체에게 가장 성공적이다. 게다가, 박테리아와 고고학과 같은 원핵생물들이 제한된 공통 형태학들을 공유하기 때문에, 그들의 화석은 그들의 조상들에 대한 정보를 제공하지 않는다.
원핵생물들은 약 30억~40억년 전부터 지구에 살았다.[17][18] 이후 몇 십억 년 동안 이들 유기체에서 형태학이나 세포조직에 뚜렷한 변화가 일어나지 않았다.[19] 진핵 세포는 16억년에서 27억년 사이에 출현했다. 세포 구조의 다음 주요한 변화는 박테리아가 내시경증이라고 불리는 협동조합에서 진핵세포에 휩싸였을 때 일어났다.[20][21] 그 후, 삼킨 박테리아와 숙주 세포는 공진화를 거쳤고, 박테리아는 미토콘드리아나 수소좀으로 진화했다.[22] 또 다른 시아노박테리아 같은 유기체들이 포획되면서 해조류와 식물에서 엽록체들이 형성되었다.[23]


생명의 역사는 에디아카란 시대에 바다에 다세포 유기체가 나타나기 시작한 약 6억1000만년 전까지만 해도 단세포 원핵생물 및 진핵생물의 역사였다.[17][26] 다세포의 진화는 해면체, 갈색 해조류, 시아노박테리아, 슬라임 몰드, 멕소박테리아와 같은 다양한 유기체에서 여러 독립적인 사건에서 일어났다.[27] 2016년 과학자들은 약 8억년 전에 GK-PID라고 불리는 단일 분자의 사소한 유전적 변화가 유기체가 단일 세포 유기체에서 많은 세포 중 하나로 이동하는 것을 허용했을지도 모른다고 보고했다.[28]
이러한 최초의 다세포 유기체가 출현한 직후, 캄브리아기 폭발이라고 불리는 사건에서 약 1천만 년의 기간에 걸쳐 놀라운 양의 생물학적 다양성이 나타났다. 여기서 현대 동물의 대다수는 화석 기록에서 나타났고, 그 후에 멸종된 독특한 선들도 발견되었다.[29] 광합성에 의한 대기 중의 산소가 축적되는 등 캄브리아 폭발에 대한 다양한 촉발제가 제시되어 왔다.[30]
배경

프로카리오테와 eukaryote라는 단어는 프로가 "이전"을, eu는 "웰" 또는 "진실"을, karyon은 "넛", "커널" 또는 "뉴클레우스"를 의미하는 그리스어에서 유래되었다.[31][32][33] 그래서 어원학적으로 원핵은 "핵 앞의"을 의미하고 진핵은 "진정한 핵"을 의미한다.
원핵생물과 진핵생물의 생물형태의 분업은 미생물학자 로저 스타니어와 C. B. 반 니엘이 1962년 논문 '박테리아의 개념'에서 확고히 정립했다.[34] 이러한 분류의 한 가지 이유는 당시 흔히 청록색조류(현재의 시아노박테리아라고 불리던 것)가 식물로 분류되는 것을 중단하지만 박테리아와 함께 분류되는 것을 멈추게 하기 위해서였다.
1990년 칼 워즈 외는 3도메인 제도를 도입했다.[35][36] 원핵생물들은 고고학과 박테리아라는 두 개의 영역으로 나뉘었고 진핵생물들은 그들 자신의 권리로 영역이 되었다. 이전의 분류들과 주요한 차이점은 고대의 박테리아와 분리하는 것이다.



지구상에 생명체가 존재한다는 가장 초기 증거는 37억년 된 암석에서 발견된 생물학적 탄소 서명과 스트로마톨라이트 화석으로부터 나온다.[37][38] 2015년에는 41억년 된 암석에서도 '생물의 흔적'이 발견됐다.[39][40] 2017년, 지구에서 가장 오래된 생명체의 화석화된 미생물의 형태로 보고되었는데, 이는 해양이 44억 4천만년 전에 형성된 지 얼마 되지 않은, 그리고 지구의 45억 4천만년 후에 발견된 것이다.가거라[41][42]
공존하는 박테리아와 고대의 미생물 매트는 초기 아르칸 이온에서 지배적인 삶의 형태였으며 초기 진화의 많은 주요 단계들이 이러한 환경에서 일어난 것으로 생각된다.[43] 3.5 Ga를 중심으로 광합성이 진화하면서 대기 중에 폐제품 산소가 축적되어 2.4 Ga 전후로 시작하는 대산소화 현상이 일어났다.[44]
진핵생물의 가장 초기 증거는 1.85 Ga에서 유래했으며,[45][46] 그것들이 더 일찍 존재했을 수도 있지만, 그들이 신진대사에 산소를 사용하기 시작하면서 그들의 다양화가 가속화되었다. 이후 1.7 Ga 전후로 다세포 유기체가 나타나기 시작했으며, 분화된 세포가 전문화된 기능을 수행했다.[47]

원핵생물을 포함한 공기 중에 떠다니는 미생물들이 기상 시스템 위는 물론 상업용 항공로 아래는 지구를 돌고 있다.[51] 일부 지각 미생물은 지상 먼지 폭풍에서 쓸려 올라오지만 대부분은 해양 미생물이 바다 스프레이를 통해 생겨난다. 2018년 과학자들은 지구 전역의 1평방미터마다 매일 수억 개의 바이러스와 수천만 개의 박테리아가 축적되고 있다고 보고했다.[52][53]
바다 밑의 미세한 생명체들은 다양하지만 해양 생태계에서 바이러스의 역할과 같이 여전히 잘 이해되지 않는다.[54] 대부분의 해양바이러스는 식물과 동물에는 무해하지만 염수와 담수 생태계 규제에 필수적이다.[55] 그들은 수생 미생물 집단의 박테리아와 고고학을 감염시키고 파괴하며, 해양 환경에서 탄소를 재활용하는 가장 중요한 메커니즘이다. 죽은 박테리아 세포에서 방출된 유기 분자는 신선한 박테리아와 조류 성장을 자극한다.[56] 바이러스 활동은 또한 생물학적 펌프, 즉 탄소가 심해에 격리되는 과정에 기여할 수 있다.[57]
해양세균




박테리아는 원핵 미생물의 큰 영역을 구성한다. 전형적으로 몇 마이크로미터 길이의 박테리아는 구에서부터 막대, 나선형까지 다양한 모양을 가지고 있다. 박테리아는 지구상에 나타난 최초의 생명체 중 하나였으며, 대부분의 서식지에 존재한다. 박테리아는 토양, 물, 산성 온천, 방사성 폐기물,[61] 그리고 지각의 깊은 부분에 서식한다. 박테리아는 또한 식물과 동물과 공생하고 기생하는 관계에서 산다.
한때 정신분열증을 구성하는 식물로 여겨졌던 박테리아는 현재 원핵생물로 분류된다. 동물 세포나 다른 진핵 세포와는 달리 박테리아 세포에는 핵이 들어 있지 않고 거의 막에 묶인 오르가넬을 품고 있지 않다. 비록 박테리아라는 용어가 전통적으로 모든 원핵생물을 포함했지만, 원핵생물은 고대 공통의 조상으로부터 진화한 두 개의 매우 다른 유기체 그룹으로 이루어진다는 1990년대 발견 이후 과학적인 분류가 바뀌었다. 이러한 진화영역은 박테리아와 고세아라고 불린다.[62]
현대 박테리아의 조상은 약 40억년 전 지구상에 나타난 최초의 생명체였던 단세포 미생물이었다. 약 30억년 동안 대부분의 유기체는 미세했고, 박테리아와 고고학은 생명체의 지배적인 형태였다.[63][64] 스트로마톨라이트와 같은 박테리아 화석이 존재하지만, 이들의 독특한 형태학이 결여되어 있어 박테리아 진화의 역사를 조사하거나 특정 박테리아 종의 발생 시기를 추정하는 데 이용되는 것을 막는다. 그러나, 유전자 배열은 박테리아 혈통을 재구성하는 데 사용될 수 있으며, 이러한 연구는 박테리아가 고고학/유핵 혈통에서 먼저 분리되었다는 것을 보여준다.[65] 박테리아는 또한 고대와 진핵생물의 두 번째 큰 진화적 분리에 관여했다. 여기서 진핵생물은 고대 박테리아가 진핵세포의 조상들과 내분비생제 결합으로 들어간 데서 비롯되었는데, 그 자체가 고대와 관련이 있을 가능성이 있었다.[21][66] 이것은 아직 알려진 모든 에우카리아에서 발견되는 미토콘드리아나 수소소체를 형성하기 위해 알파프로테오박테리아 공생의 원핵세포에 의해 퍼지는 것을 포함한다. 이후 이미 미토콘드리아가 함유된 일부 진핵생물도 시아노박테리아 같은 유기체를 삼켰다. 이로 인해 해조류와 식물에서 엽록체가 형성되었다. 심지어 나중에 일어난 내분비생물학적 사건에서 유래한 조류도 있다. 여기서 진핵생물은 진핵생성 해조류를 삼켜 '제2세대' 플라스티드로 발전했다.[67][68] 이것은 2차 내분비증이라고 알려져 있다.
박테리아는 일정한 크기로 성장한 다음 무성 생식의 한 형태인 이항 핵분열을 통해 번식한다.[69] 최적의 조건 하에서 박테리아는 매우 빠르게 성장하고 분열될 수 있으며, 박테리아 개체수는 매 9.8분마다 두 배씩 증가할 수 있다.[70]
펠라지박터 우비크와 그 친척들은 바다에서 가장 풍부한 미생물일 수 있으며, 그들은 아마도 세계에서 가장 풍부한 박테리아일 것이라고 주장되어 왔다. 그들은 전체 미생물 플랑크톤 세포의 약 25%를 구성하고, 여름에는 온대 해양 표면수에 존재하는 세포의 약 절반을 차지할 수 있다. P. ubique와 친척의 총 풍부함은 약 2 × 1028 미생물로 추정된다.[71] 그러나 P. ubique를 공격하는 박테리오파지 HTVC010P가 발견되어 아마도 지구상에서 가장 흔한 유기체일 것이라는 것이 2013년 2월 네이처에 보고되었다.[72][73]
로즈박터 역시 바다에서 가장 풍부하고 다재다능한 미생물 중 하나이다. 그들은 해안에서 열린 바다, 그리고 해빙에서 해저에 이르기까지 다양한 종류의 해양 서식지에 걸쳐 다양하며, 해안 해양 박테리아의 약 25%를 차지한다. 로소박터 속 구성원들은 해양 생물 화학적 순환과 기후 변화에서 중요한 역할을 하며, 해양 환경에서 총 탄소의 상당 부분을 처리한다. 그들은 방향성 화합물을 분해하고 미량 금속을 흡수할 수 있는 공생 관계를 형성한다. 그것들은 양식업과 정족수 감지에 널리 사용된다. 녹조 기간 동안 원핵생물 집단의 20~30%가 로소박터다.[74][75]
알려진 가장 큰 박테리아인 해양 티오마르가리타 나미비엔시스는 육안으로 볼 수 있고 때로는 0.75mm(750μm)에 달하기도 한다.[76][77]


시아노박테리아

.jpg)

시아노박테리아는 햇빛을 화학 에너지로 바꾸는 능력을 진화한 최초의 유기체였다. 그들은 단세포에서 필라멘트까지 다양한 박테리아를 형성하고 식민지 종을 포함한다. 그들은 지구의 거의 모든 곳에서 발견된다: 축축한 토양, 담수와 해양 환경, 그리고 심지어 남극 암석에서도 발견된다.[79] 특히 일부 종은 바다에 떠다니는 표류 세포로 발생하는데, 이는 식물성 플랑크톤 중 첫 번째 종이다.
광합성을 사용한 최초의 생산자는 약 23억년 전 해양 시아노박테리아였다.[80][81] 광합성의 부산물로서 시아노박테리아에 의한 분자 산소의 방출은 지구 환경의 세계적 변화를 유도했다. 산소는 그 당시 지구상의 대부분의 생명체에게 독성이 있었기 때문에, 이것은 산소 내성 유기체의 거의 멸종으로 이어졌고, 이것은 주요한 동식물 종의 진화를 재연결시킨 극적인 변화였다.[82]

박테리아와 상호작용하여 먼지로부터 철분을 획득하다.
b.트라이코드뮴은 먼지의 침적률이 높은 영양소가 부족한 해양 지역에서 대규모의 꽃을 피울 수 있는데, 부분적으로 먼지를 포착하고, 중심에 놓고, 이후에 녹이는 독특한 능력 때문이다.
c. 제안된 분진결합 Fe 획득 경로: 군집 내에 거주하는 박테리아는 군집 중심부의 먼지 입자와 반응하여 용해된 Fe(c-II)를 생성하는 시더로포어(c-I)를 생성한다. 이 용해된 Fe는 시데로포레스에 의해 복잡하게 되고, 트리코데스뮴과 그 거주 박테리아(c-III)에 의해 모두 획득되어, 컨소시엄의 양쪽 파트너에게 상호 이익을 가져다 준다.[83]



시네코코쿠스, 널리 퍼진 해양 시아노박테리움



1986년에 발견된 아주 작은 (0.6 µm) 해양 시아노박테리움 프로클로로코쿠스는 오늘날 해양 먹이 사슬의 밑부분의 중요한 부분을 형성하며, 개방된 대양의[85] 광합성의 많은 부분과 지구 대기에 있는 산소의 약 20%를 차지한다.[86] 그것은 아마도 지구상에서 가장 풍부한 속일 것이다: 1밀리리터의 표면 바닷물은 10만개 이상의 세포를 포함할 수 있다.[87]
원래 생물학자들은 시아노박테리아를 조류로 분류해 '푸른 녹조'라고 불렀다. 더 최근의 견해는 시아노박테리아가 박테리아여서 조류와 같은 왕국에 있지도 않다는 것이다. 대부분의 당국은 모든 원핵생물, 즉 시아노박테리아를 조류 정의에서 배제한다.[88][89]
| 외부 영상 | |
|---|---|
기타세균
시아노박테리아 외에 다른 해양 박테리아는 어디에나 존재하거나 바다에서 중요한 역할을 할 수 있다. 여기에는 기회주의적인 코피오트로피인 알테로모나스 마클레오디 등이 포함된다.[90][91]
해양고고대

고고학(고대의[93] 그리스어)은 단세포 미생물의 영역과 왕국을 구성한다. 이 미생물들은 세포핵이나 세포 안에 다른 막에 묶인 유기체가 없다는 것을 의미하는 원핵생물이다.
고세아는 처음에 박테리아로 분류되었지만, 이 분류는 시대에 뒤떨어져 있다.[94] 고고 세포는 다른 두 생명 영역인 박테리아와 유카리오타로부터 분리되는 독특한 성질을 가지고 있다. 고세아는 더 나아가 여러 개의 공인된 필라로 나뉜다. 대다수가 실험실에서 격리되지 않고 환경에서 추출한 샘플에서 핵산을 분석함으로써만 검출되었기 때문에 분류가 어렵다.
박테리아와 고고학은 일반적으로 크기와 모양이 비슷하지만, 몇몇 고고자는 할로카드라툼 월스비이의 평평하고 네모난 모양의 세포와 같이 매우 이상한 모양을 가지고 있다.[95] 이러한 박테리아와의 형태학적 유사성에도 불구하고, 고고아는 진핵생물의 그것, 특히 전사 및 번역에 관련된 효소와 더 밀접하게 관련된 유전자와 몇 가지 대사 경로를 가지고 있다. 고고학 생화학의 다른 측면은, 고고학처럼 세포막에 있는 에테르 지질에 의존하는 것과 같은, 독특하다. 고고학은 진핵생물보다 더 많은 에너지원을 사용한다: 이것들은 설탕과 같은 유기 화합물에서부터 암모니아, 금속 이온 또는 심지어 수소 가스에 이르기까지 다양하다. 소금강화 고고학(Haloarchaea)은 햇빛을 에너지원으로 사용하고 다른 종류의 고고학에서는 탄소를 고정하지만 식물이나 시아노박테리아와는 달리 알려진 고고학 종은 둘 다 하지 않는다. 고세아는 이항 핵분열, 분열 또는 싹트기에 의해 무성하게 번식한다; 박테리아나 진핵생물과는 달리 알려진 종은 포자를 형성하지 않는다.
고아는 특히 바다에 많고, 플랑크톤에 있는 고아는 지구상에서 가장 풍부한 생물군 중 하나일 것이다. 고세아는 지구 생활의 주요 부분이며 탄소 순환과 질소 순환 모두에서 역할을 할 수 있다. 크레나르카에오타(세포)는 해양환경에 매우 풍부하다고 생각되는 고고학의 망막으로, 탄소를 고정시키는 주요 원인 중 하나이다.[96]

에모세포는 해양 고고학 중에서 가장 풍부할 수 있다.

소금에 가까운 물에서 발견되는 할로박테리아는 현재 고고학자로 인식되고 있다.

고고학 할로카드라툼 월스비이의 평평하고 네모난 모양의 세포

메탄을 생산하는 해양 고고학자인 메탄노사르시나 바케리






나노카기움은 2002년 열수분출구에서 발견된 해양고고학의 한 종이다. 그것은 약 섭씨 80도의 온도에서 자라는 열성분이다. 나노카케움은 고고학 이그니코쿠스에 대한 의무적인 공생으로 보인다. 나노카케움 이퀴탄은 지질을 합성할 수 없지만 숙주로부터 얻어내기 때문에 생존하기 위해서는 숙주 유기체와 계속 접촉해야 한다. 이것의 세포는 지름이 400 nm에 불과해 알려진 가장 작은 세포 유기체 중 하나이며, 가장 작은 것으로 알려진 고고학자로 알려져 있다.[97][98]
해양 고고학은 다음과 같이 분류되었다.[99][100][101][102][103]
- 마린 그룹 I(MG-I 또는 MGI): Ia(일명 I.a)부터 Ia(일명 I.a)까지 하위 그룹을 가진 마린 타우마카게오타
- 마린 그룹 II(MG-II): 해양 에우리아카게오타, 부분군 IIA를 포함한 포세이도식민지를[104] 최대 IID까지 주문(IIA는 포세이도니아과, IIB는 탈라사르카과)
MGII를 기생시키는 바이러스는 마그로바이러스로 분류된다. - Marine Group III (MG-III): 또한 Marine Euriarchaeota, Marine Benthic Group[105] D
- 마린 그룹 4(MG-IV): 또한 마린 에우리카게오타[106]
영양모드
프로카리오테 신진대사는 에너지의 원천, 사용하는 전자 기증자, 성장에 사용되는 탄소의 원천 등 3대 기준에 따라 영양군으로 분류된다.[107][108]
| 영양형 | 에너지원 | 탄소의 근원 | 예 |
|---|---|---|---|
| 광전자 | 햇빛 | 유기 화합물(광체소성분) 또는 탄소 고정(광체소성분) | 시아노박테리아, 녹황세균, 클로로플렉시 또는 보라색세균 |
| 리토트로프스 | 무기 화합물 | 유기 화합물(리토헤테로트로프스) 또는 탄소 고정(리토오토트로프스) | 테르메데설포박테리아, 수소화과 또는 니트로스피레아과 |
| 유기체 | 유기 화합물 | 유기 화합물(화학체질소) 또는 탄소 고정(화학) | 바실러스, 클로스트리디움 또는 엔테로박테리아과 |
해양 원핵생물들은 오랜 기간 동안 매우 다양해졌다. 원핵생물들의 신진대사는 진핵생물들의 대사보다 훨씬 더 다양해서, 많은 구별되는 원핵생물 종류로 이어진다. 예를 들어 진핵생물처럼 광합성이나 유기화합물을 에너지로 사용하는 것 외에도 해양 원핵생물들은 황화수소 같은 무기 화합물로부터 에너지를 얻을 수 있다. 이것은 해양 원핵생물들이 남극 대륙의 얼음 표면처럼 추운 환경에서 극한 환경에서도, 극저온 생물학에서 연구되고, 해저 열수 분출물처럼 뜨겁거나, 또는 (할로파일)과 같은 높은 식염수 조건에서도 극한 동물로 번성할 수 있게 한다.[109] 몇몇 해양 원핵생물들은 다른 해양생물의 몸 안이나 몸 위에서 공생적으로 산다.

- 광생성은 박테리아 분류에서 항상 주된 역할을 해야 하는 특히 중요한 표식이다.[110]
- 에어로빅산소성 광생성 박테리아(AAPBs)는 널리 분포된 해양 플랑크톤으로, 오픈오션 미생물 집단의 10% 이상을 차지할 수 있다. 마린 AAPB는 2개의 마린(에리스로박터, 로즈박터) 제네라로 분류된다. 그들은 특히 공동체의 24%로 밝혀진 과두성 환경에서 풍부할 수 있다.[111] 이것들은 에너지를 생산하기 위해 빛을 사용하지만 이산화탄소를 주요 탄소원으로 사용할 수 없는 이질성 유기체들이다. 대부분은 필수적으로 에어로빅인데, 이것은 그들이 자라기 위해 산소를 필요로 한다는 것을 의미한다. 현재 데이터에 따르면 해양 박테리아는 수 일 동안 생성되는 시간을 가지고 있는 반면, AAPB는 훨씬 더 짧은 생성 시간을 가지고 있다는 새로운 증거가 존재한다.[112] 해안/선박 해역은 종종 더 많은 양의 AAPB를 가지고 있으며, 어떤 것은 13.51%의 AAPB를 가지고 있다. 식물성 플랑크톤도 AAPB%에 영향을 미치지만 이 분야에서는 연구가 거의 이뤄지지 않았다.[113] 그들은 또한 세계 해양의 가장 과점적인 정권을 포함한 다양한 과점 조건에서도 풍부할 수 있다.[114] 그것들은 유향 지역에 전세계적으로 분포되어 있으며 해양 미생물 집단의 지금까지 인식되지 않은 구성요소를 대표하고 있는데, 이는 해양의 유기 탄소와 무기 탄소의 순환에 중요한 것으로 보인다.[115]
- 보라색 박테리아:
- 제타프로테오박테리아: 철분 산화 중성미자 화학 비석소 자급생물로, 전 세계적으로 강초와 해양 서식지에 분포한다.
- 수소산화세균은 에어로브와 혐기성으로 나눌 수 있는 기능적 자가영양이다. 전자는 수소를 전자 공여자로, 산소를 수용자로, 후자는 황산염이나 이산화질소를 전자 수용기로 사용한다.[116]
운동성

운동성은 신진대사에너지를 이용하여 독립적으로 움직이는 유기체의 능력이다.
플라겔라 운동성


박테리아와 고고학 둘 다인 원핵생물들은 주로 이동에 플라겔라를 사용한다.
- 박테리아 플라겔라는 나선형 필라멘트인데, 각각 기저에 시계 방향 또는 시계 반대 방향으로 회전할 수 있는 회전 모터가 있다.[118][119][120] 그것들은 여러 종류의 박테리아 운동성 중 두 가지를 제공한다.[121][122]
- 고대의 플라겔라는 아르카엘라라고 불리며, 박테리아 플라겔라와 거의 같은 방식으로 기능한다. 구조적으로 대퇴골은 표면적으로는 세균성 편모세포와 비슷하지만, 여러 세부적인 면에서는 차이가 있어 비호모성으로 간주된다.[123][117]
박테리아가 사용하는 회전 운동 모델은 플라겔라를 이동시키기 위해 전기 화학적 경사의 양자를 사용한다. 박테리아의 플라겔라에서 토크는 플라겔럼의 밑부분 주위에 양자를 전도하는 입자에 의해 생성된다. 박테리아에서 플라겔라의 회전 방향은 플라겔라 모터의 둘레를 따라 양성자 채널을 점유하는 것에서 나온다.[124]
몇몇 진핵 세포들은 또한 플라겔라를 사용한다 - 그리고 그것들은 동물 세포뿐만 아니라 일부 양성자와 식물에서도 발견될 수 있다. 진핵 플라겔라는 순환운동이 아니라 앞뒤로 부딪치는 복잡한 세포투영이다. 원핵 플라겔라는 회전식 모터를 사용하고 진핵 플라겔라는 복잡한 슬라이딩 필라멘트 시스템을 사용한다. 진핵 플라겔라는 ATP가 주도하고, 원핵 플라겔라는 ATP가 주도하거나(아카에아) 양성자가 주도할 수 있다.[125]
| 외부 영상 | |
|---|---|
경련 운동성
경련 운동성은 표면 위를 기어다니는 박테리아 운동성의 일종이다. 경련은 세포의 외부로부터 확장되어 주변의 고체 기판과 결합하고 수축하면서 그러플링 후크의 작용과 유사한 방식으로 세포를 앞으로 당기는 타입 IV 필라멘트라고 불리는 머리카락 같은 필라멘트의 활동으로 매개된다.[126][127][128] 트위칭 운동성은 현미경으로 볼 때 개별 세포의 특징적인 육포와 불규칙한 움직임에서 유래한다.[129]
글라이딩 운동성
글라이딩 운동성은 플라젤라나 필리와 같은 추진 구조와는 독립된 번역의 일종이다.[130] 글라이딩은 미생물이 낮은 수용성 필름의 표면을 따라 이동할 수 있게 해준다. 이 운동성의 메커니즘은 부분적으로만 알려져 있다. 글라이딩 속도는 유기체마다 다르며, 방향의 반전은 겉보기에는 일종의 내부 시계에 의해 조절된다.[131] 예를 들어, 아피콤플렉산들은 1-10μm/s 사이의 빠른 속도로 이동할 수 있다. 이와는 대조적으로 Myxococcus xantus 박테리아는 5 μm/min의 속도로 활공한다.[132][133]
군더더기 운동성
군집 운동성은 고형 또는 반고형 표면에 걸쳐 박테리아 집단의 신속한(2~10μm/s) 및 조정된 변환이며,[134] 박테리아 다세포성 및 군집행동의 예다. 크라우밍 운동성은 조르겐 헨리히센에 의해 1972년에 처음 보고되었다.[135]
비동기
비운동성 종은 그들이 그들 자신의 힘 아래, 환경을 통해 그들 자신을 추진하게 할 수 있는 능력과 구조가 부족하다. 비운동성 박테리아가 찌르는 관에서 배양되면 찌르는 선을 따라서만 자란다. 박테리아가 움직이면 선이 확산되어 매질로 확장될 것이다.[136]
택시: 방향 이동
자석축


자석성 박테리아는 지구 자기장의 자기장 선을 따라 방향을 정한다.[138] 이 정렬은 이러한 유기체가 최적의 산소 농도의 영역에 도달하는 데 도움을 준다고 믿어진다.[139] 이 임무를 수행하기 위해, 이 박테리아들은 자석 결정체를 포함하고 있는 자석이라고 불리는 생물학적 유기체를 가지고 있다. 미생물이 환경의 자기 특성에 반응하여 움직이는 생물학적 현상을 자석축이라고 한다. 그러나 이 용어는 택시의 다른 모든 적용은 자극-반응 메커니즘을 수반한다는 점에서 오해의 소지가 있다. 동물들의 자석감각과는 대조적으로, 박테리아는 박테리아를 정렬 상태로 만드는 고정 자석을 포함하고 있다. 심지어 죽은 세포도 나침반 바늘처럼 정렬 상태로 끌려간다.[139]
해양 환경은 일반적으로 저농도의 영양소가 조류와 난류에 의해 안정적이거나 간헐적으로 움직인다는 특징이 있다. 해양세균은 수영과 방향감지-반응 시스템을 이용하여 영양분 구배에서 유리한 곳으로 이동하기 위한 전략을 개발했다. 자석성 박테리아는 지구의 자기장을 활용하여 화학적으로 층화된 퇴적물이나 물기둥에서 지속성과 증식에 가장 유리한 장소인 난독성-난독성 인터페이스로 하향 수영을 용이하게 한다.[140]

그들의 위도와 박테리아가 적도의 북쪽에 있는지 남쪽에 있는지에 따라, 지구의 자기장은 가능한 두 개의 극성 중 하나를 가지고 있으며, 해양 깊이로 다양한 각도로 가리키며, 일반적으로 더 많은 산소가 풍부한 표면으로부터 떨어져 있는 방향을 가지고 있다. 에어로트축은 박테리아가 산소 구배에서 최적의 산소 농도로 이동하는 반응이다. 다양한 실험에서 자석축과 에어로트축이 자석성 박테리아와 결합하여 작용한다는 것을 분명히 밝혀냈다. 물방울에서 단방향 수영 자석성 박테리아는 난독성 조건(최적 산소 농도보다 큼)과는 반대로 감소 조건(최적 산소 농도 미만)에서 수영 방향을 반대로 되돌리고 뒤로 헤엄칠 수 있는 것으로 나타났다.
형태학에 관계없이 지금까지 연구된 모든 자석성 박테리아는 플라겔라를 이용하여 운동성이 있다.[141] 특히 해양 자석성 박테리아는 수만 개의 플라겔라를 포함할 수 있는 정교한 플라겔라 기구를 보유하는 경향이 있다. 그러나 최근 몇 년간 광범위한 연구에도 불구하고 자기장에서의 정렬에 대응하여 자기성 박테리아가 플라겔라 모터를 조종하는지는 아직 밝혀지지 않았다.[140] 자석 박테리아와 공생하는 것은 일부 해양 원생들에게 자석감각에 대한 설명으로 제안되었다.[142] 유사한 관계가 척추동물에게도 자기관찰의 근거가 될 수 있는지에 대한 연구가 진행 중이다.[143] 가장 오래되고 모호하지 않은 자성 광석은 영국 남부의 백악기 분필 침대에서 나온 것이지만,[144] 자성 광석은 19억년 된 군플린트 체르트로 확대되는 확실한 보고는 더 적다.[145]
가스 바쿠올
| 다음에 대한 시리즈 일부 |
| 플랑크톤 |
|---|
 |
몇몇 해양 원핵생물들은 가스 vacuole을 가지고 있다. 가스 vacuole은 해양 박테리아와 고고학이 부력을 조절할 수 있도록 가스에 자유롭게 침투할 수 있는 나노 물질이다. 그들은 스핀들 모양의 막으로 묶인 베시클의 형태를 취하며, 몇몇 시아노박테리아를 포함한 몇몇 플랑크톤 원핵에서 발견된다.[146] 세포가 계속 광합성을 할 수 있도록 물기둥 상류에 유지하려면 양성 부력이 필요하다. 가스 vacuole은 내부 표면이 높은 단백질 껍질로 이루어져 있어 물에는 불침투성이지만 대부분의 가스에는 침투할 수 있다. 기체는 속이 빈 원통이기 때문에 주변 압력이 높아지면 무너지기 쉽다. 자연선택은 땋은 호스파이프의 녹색 실처럼 외부 강화 단백질인 GvpC를 포함하여 기체 방광의 구조를 미세하게 조정하여 버클에 대한 저항을 극대화했다. 가스 음낭의 직경과 그것이 붕괴할 압력 사이에는 간단한 관계가 있다 – 가스 음낭이 넓을수록 약해진다. 그러나 넓은 가스 베시클은 좁은 가스 베시클보다 단백질 단위당 부력이 더 높아 효율적이다. 서로 다른 종은 서로 다른 직경의 가스 배실체를 만들어 물기둥의 다른 깊이를 식민지화할 수 있게 한다(급성장, 가장 위층에 넓은 가스 배실체를 가진 경쟁이 치열한 종, 더 깊은 층에 강한 좁은 가스 배실체를 가진 종).
그 세포는 가스 vesicle을 합성하여 물기둥의 높이를 달성한다. 세포가 상승함에 따라 광합성을 증가시킴으로써 탄수화물 부하를 증가시킬 수 있다. 너무 높으면 세포가 광피해를 입게 되고 죽을 수도 있지만 광합성 과정에서 생성되는 탄수화물은 세포의 밀도를 높여 세포가 가라앉게 한다. 어두운 시간 동안 광합성, 탄수화물 포물선염에 의한 탄수화물 증식의 하루 주기는 물기둥에서 세포의 위치를 미세하게 조정하여 탄수화물 수치가 낮고 광합성이 필요할 때 표면으로 끌어올려 세포의 자동차에 의한 유해한 자외선에 의해 가라앉게 하는 데 충분하다.보이드레이트 수치가 보충되었다. 탄수화물이 극도로 과잉되면 세포 내부 압력에 상당한 변화가 일어나 가스 베실체가 버클링되고 붕괴되고 세포가 빠져 나간다.
큰 바쿠올은 티오플로카, 베기아토아, 티오마르가리타 등 세 종류의 황세균에서 발견된다. 시토솔은 이러한 제네랄에서 극도로 감소하고 바쿠올은 세포의 40~98%를 차지할 수 있다.[147] 바쿠올은 고농도의 질산 이온을 함유하고 있어 저장용 오르가넬로 생각된다.[148]
생물 발광


생물 발광 박테리아는 주로 바닷물, 해양 퇴적물, 부패한 물고기의 표면, 해양 동물의 내장에 존재하는 빛을 내는 박테리아다. 흔한 것은 아니지만, 박테리아 생물 발광은 지상이나 담수 박테리아에서도 발견된다.[126] 이 박테리아는 자유 생활일 수도 있고(비브리오 하베이와 같은) 하와이 밥테일 오징어(Alivibrio fischeri) 또는 육상 네마토드(Photorhabdus luminescens)와 같은 동물과 공생할 수도 있다. 숙주 유기체는 이 박테리아들에게 안전한 집과 충분한 영양분을 제공한다. 대신, 호스트들은 위장, 먹이 그리고/또는 짝짓기를 위해 박테리아에 의해 생성된 빛을 사용한다. 생물 발광 박테리아는 두 참가자가 거의 동등하게 이익을 얻는 다른 유기체와의 공생 관계를 진화시켰다.[150] 박테리아가 발광 반응을 사용하는 또 다른 가능한 이유는 박테리아 세포 밀도에 반응하여 유전자 발현을 조절할 수 있는 능력인 정족수 감지 때문이다.[151]
하와이 단발 오징어는 오징어 맨틀의 특별한 빛 기관에 서식하는 생물 발광 박테리아 알리비브리오 피쉐리와 공생하며 산다. 이 박테리아는 오징어에 의해 설탕과 아미노산을 먹이고, 그 대가로 오징어의 실루엣을 아래에서 볼 때 숨겨주고, 맨틀 꼭대기에 부딪히는 빛의 양과 일치시켜 반감시킨다.[152] 오징어는 동물-박테리아 공생의 모범적인 유기체 역할을 하며 박테리아와의 관계가 널리 연구되어 왔다.
비브리오 하베이는 막대 모양의 모틸(폴라플라겔라를 통해) 생물 발광성 박테리아로, 30~35℃ 사이에서 최적으로 자란다. 그것은 열대 해양에서 자유 수영하는 것을 발견할 수 있고, 일반적으로 해양 동물의 내장 미세플로라에서 그리고 많은 해양 동물의 일차적, 기회주의적 병원체 둘 다에서 발견될 수 있다.[153] 밤 동안 바닷물에서 균일한 푸른 빛이 뿜어져 나오는 은하수 효과의 원인으로 생각된다. 어떤 빛은 거의 6,000 평방 미 (16,000 km2)를 덮을 수 있다.
미생물로돕신

광섬유대사 작용은 세 가지 에너지 변환 색소 중 하나인 엽록소, 박테리오클로로필, 레티날 중 하나에 의존한다. 레티날은 로돕신에서 발견되는 색소동물이다. 빛 에너지를 변환하는 데 있어서 엽록소의 중요성은 수십 년 동안 쓰여져 왔지만, 망막 색소를 기반으로 한 광자생식은 이제 막 연구되기 시작하고 있다.[155]
.jpg)
2000년에 에드워드 드롱이 이끄는 미생물학자 팀은 해양 탄소 및 에너지 사이클의 이해에서 중요한 발견을 했다. 그들은 이전에 박테리아에서 들어본 적이 없었던 단백질 로돕신의 생산을 담당하는 여러 종의 박테리아에서[157][158] 유전자를 발견했다. 세포막에서 발견되는 이러한 단백질은 햇빛에 의해 로도신 분자의 구성이 변화하여 내부에서 양성자가 펌핑되어 에너지를 발생시키는 후속 유입으로 인해 빛에너지를 생화학적 에너지로 변환시킬 수 있다.[159] 고고학처럼 생긴 이 로돕신들은 비록 복잡한 다세포 유기체에서는 드물지만 박테리아와 고고학뿐만 아니라 서로 다른 세아, 양성자들 사이에서 나중에 발견되었다.[160][161][162]
2019년의 연구는 이러한 "태양 관측 박테리아"가 이전에 생각했던 것보다 더 널리 퍼져 있고, 해양이 지구 온난화의 영향을 받는 방식을 바꿀 수 있다는 것을 보여준다. 그는 "이번 연구 결과는 해조류에서 엽록소에 의해 바다의 거의 모든 햇빛을 포착한다는 교과서에서 발견된 해양생태학의 전통적인 해석에서 벗어나게 된다"고 말했다. 대신에, 로돕신 장착 박테리아는 하이브리드 자동차와 같은 기능을 하는데, 대부분의 박테리아가 그러하듯이, 유기 물질과 영양소가 부족할 때 햇빛에 의해 작동된다."[163][155]
지구상의 원래 생명체는 엽록소가 아닌 망막 기반이었다고 추측하는 보라색 지구 가설이라고 불리는 천체 생물학적 추측이 있는데, 이것은 지구를 녹색 대신 보라색으로 보이게 만들었을 것이다.[164][165]
공생
어떤 해양 생물들은 박테리아나 고고학과 공생한다. 폼페이 벌레는 최고 80℃의 온도에서 열수 분출구에 의해 매우 깊은 곳에서 산다. 그들은 등에 털이 많은 것으로 보이는 것을 가지고 있지만, 이 "해어"는 사실 노틸리아 프로푼디콜라와 같은 박테리아의 군집인데, 이 박테리아는 이 벌레에게 어느 정도 절연 효과를 줄 것으로 생각된다. 벌레 등의 분비샘은 공생의 한 형태인 박테리아가 먹이를 주는 점액을 분비한다.

폼페이 벌레의 "요정" 등은 공생 박테리아의 군집이다.

헤시오카에카 메탄니콜라는 메탄 빙하 위에서 매우 깊은 곳에서 살고 있으며 종괴를 대사하는 박테리아와 공생하여 생존하는 것으로 보인다.[166]

Olavius algarvensis는 그 영양을 위해 5종의 공생균에 의존한다.





내시경 박테리아는 다른 유기체의 몸이나 세포 안에 사는 박테리아다. 시아노박테리아의 일부 종류는 내시경비온이며, 시아노박테리아는 질소고정을 할 수 있는 유전자를 가지고 있는 것으로 밝혀졌다.[168]
유기체는 일반적으로 서식지에서 자원의 이용이 제한되거나 식량의 한계로 인해 공생관계를 형성한다. 열수 분출구 근처에 위치한 홍합(바스티모디올루스)과 연관된 공생, 화학합성 박테리아는 수소를 에너지원으로 활용할 수 있게 하는 유전자를 가지고 있는데, 이는 황이나 메탄보다 에너지를 생산하는 에너지원이다.[169]
올라비우스 알가르벤시스는 지중해 연안 퇴적물에 서식하며 영양을 위해 공생균에 의존하는 벌레다. 그것은 큐티클 아래에 두 개의 황화물 산화, 두 개의 황산염 감소, 그리고 한 개의 스피로차테의 다섯 가지 다른 종류의 박테리아와 함께 살고 있다. 공생균은 또한 지렁이가 수소와 일산화탄소를 에너지원으로 사용하고, 말라이트와 아세테이트와 같은 유기 화합물을 대사할 수 있도록 한다.[170][171]
북쪽의 별 산호인 아스트랑기아 포쿨라타는 미국 동부 해안을 따라 널리 기록되어 있는 온대성 돌로니 산호다. 산호는 동물원탄첼레(algal symbellae)와 함께 살 수 있고 없어도 살 수 있어 공생 상태와 연관된 미생물 공동체 상호작용을 연구하는 이상적인 모델 유기체가 된다. 그러나 Illumina 플랫폼에 의해 생산된 시퀀스의 길이가 프라이머와 프로브의 설계에 불충분하기 때문에(약 250 base pairs) 전체 길이 16S rRNA 시퀀스가 부족하여 프라이머와 프로브를 보다 구체적으로 타깃팅하는 키 미생물 그룹에 대한 프라이머와 프로브를 개발할 수 있는 능력이 방해받고 있다.[172] Goldsmith 외 연구진은 2019년에 Sanger 시퀀싱이 차세대 심층 시퀀싱에 의해 검출된 생물학적 관련 다양성을 재현하는 동시에 프로브 및 프라이머 설계에 유용한 더 긴 시퀀스를 만들 수 있음을 시연했다(오른쪽 다이어그램 참조).[173]
해양식품망에서의 역할

세계 대양의 대부분은 어둠 속에 있다. 얇은 조명이 들어오는 표면층(표면에서 50~170m 사이의 광층) 내에서 발생하는 프로세스는 지구 생물권에 중요한 의미를 갖는다. 예를 들어, 이 태양빛 층에 도달하는 태양 스펙트럼의 가시 영역(일명 광합성 이용 가능한 방사선 또는 PAR)은 행성의 일차 생산성의 약 절반을 연료로 하며, 지구상의 대부분의 생명에 필요한 대기 산소의 약 절반을 담당한다.[175][176]
헤테로토스트리아성 박테리오플랑크톤은 태양빛을 발광하는 해양 먹이 그물에서 용해된 유기물(DOM)의 주요 소비국이다. 자외선에 대한 그들의 민감성(UVR)과 최근 발견된 몇몇 메카니즘이 광합성 이용 가능한 방사선(PAR)의 혜택을 받기 위해 진화했다는 사실은 자연 햇빛이 해양의 박테리아 생물화학 기능을 조절하는 데 관련되기는 하지만 역할을 예측하기는 어렵다는 것을 시사한다.[174]
바다 표면 서식지는 대기와 바다 사이의 접점에 있다. 바다 표면의 생물필름과 같은 서식지는 흔히 네우스턴이라고 불리는 표면적 질식 미생물을 서식한다. 이 거대한 공기-물 인터페이스는 지구 표면적의 70% 이상을 차지하는 주요 공기-물 교환과정의 교차점에 위치한다. 박테리오네우스턴이라 불리는 해양 표면 미세층 박테리아는 온실가스의 공기-해상 가스 교환, 기후-활성 마린 생산과 같은 실용적인 응용으로 인해 관심을 끌고 있다.에어로졸과 바다의 원격감각.[177] 특정 관심사는 미생물 생화학 공정을 통한 계면활성제(표면 활성물질)의 생산과 분해다. 공해상에 존재하는 계면활성제의 주요 공급원으로는 식물성 플랑크톤,[178] 지상유출, 대기권 침적 등이 있다.[177]

색조류와는 달리 계면활성제 관련 박테리아는 해양 색채 이미지에서 보이지 않을 수 있다. 합성개구경 레이더를 사용하여 이러한 "보이지 않는" 계면활성제 관련 박테리아를 탐지할 수 있는 능력을 갖추는 것은 구름, 안개, 대낮에 관계없이 전천후 조건에서 엄청난 이점을 가진다.[177] 이것은 매우 강한 바람에서 특히 중요하다. 왜냐하면 이것들은 가장 격렬한 공해 가스 교환과 해양 에어로졸 생산이 일어나는 조건이기 때문이다. 따라서, 컬러 위성 사진 외에도, SAR 위성 사진은 해양과 대기의 경계에서 생물물리학적 과정, 공해 온실가스 교환 및 기후 활성 해양 에어로졸 생산에 대한 글로벌 그림에 대한 추가적인 통찰력을 제공할 수 있다.[177]

오른쪽의 도표는 해양 생물학 펌프와 펠로직 먹이 그물 사이의 연관성과 선박, 위성, 자율 차량으로부터 원격으로 이러한 구성품을 샘플링할 수 있는 능력을 보여준다. 연한 푸른 물이 유향적인 반면, 더 진한 푸른 물은 황혼의 영역을 나타낸다.[179]
생체화학 사이클링에서의 역할
고세아는 탄소, 질소, 황과 같은 원소를 그들의 다양한 서식지를 통해 재활용한다.[180] 고세아는 질소 사이클에서 많은 단계를 수행한다. 여기에는 생태계에서 질소를 제거하는 반응(질산염 기반 호흡, 탈리피화 등)뿐만 아니라 질소를 도입하는 과정(질산염 동화 및 질소 고정 등)도 포함된다.[181][182]
연구원들은 최근 암모니아 산화 반응에 고고학적 관여를 발견했다. 이러한 반응은 바다에서 특히 중요하다.[183][184] 유황순환에서 황화합물을 산화시켜 성장시키는 고고학은 이 원소를 암석으로부터 방출하여 다른 유기체들이 이용할 수 있게 하지만, 설포오부스와 같이 이렇게 하는 고고학은 폐생물로 황산을 생산하며, 폐광산에서의 이러한 유기체들의 증식은 산성 광산배수 및 기타 환경 d에 기여할 수 있다.아마게.[185] 탄소 순환에서 메탄노겐 고아는 수소를 제거하고 퇴적물이나 습지와 같은 혐기성 생태계에서 분해자 역할을 하는 미생물의 개체수에 의해 유기물질의 부패에 중요한 역할을 한다.[186]
참고 항목
- 박테리오플랑크톤 계수법
- 생물 발광 박테리아
- 철분산화세균
- 펠라기박테랄레스 – 이론 능률화에 있어 유기체 모델
- 합리화 이론
참조
- ^ Youngson RM (2006). Collins Dictionary of Human Biology. Glasgow: HarperCollins. ISBN 978-0-00-722134-9.
- ^ Nelson DL, Cox MM (2005). Lehninger Principles of Biochemistry (4th ed.). New York: W.H. Freeman. ISBN 978-0-7167-4339-2.
- ^ Martin EA, ed. (1983). Macmillan Dictionary of Life Sciences (2nd ed.). London: Macmillan Press. ISBN 978-0-333-34867-3.
- ^ Fuerst JA (2010). "Beyond Prokaryotes and Eukaryotes: Planctomycetes and Cell Organization". Nature Education. 3 (9): 44.
- ^ 2002년 WHO 사망률 데이터 2007년 1월 20일 액세스
- ^ University of Georgia (10 December 2015). "Functions of global ocean microbiome key to understanding environmental changes". www.sciencedaily.com. Retrieved 11 December 2015.
- ^ Bar-On YM, Phillips R, Milo R (2018). "The biomass distribution on Earth" (PDF). Proceedings of the National Academy of Sciences of the United States of America. 115 (25): 6506–6511. Bibcode:1998PNAS...95.6578W. doi:10.1073/pnas.1711842115. PMC 6016768. PMID 29784790.
- ^ a b Doglioni C, Pignatti J, Coleman M (2016). "Why did life develop on the surface of the Earth in the Cambrian?". Geoscience Frontiers. 7 (6): 865–873. doi:10.1016/j.gsf.2016.02.001.
- ^ "Age of the Earth". United States Geological Survey. 9 July 2007. Retrieved 31 May 2015.
- ^ 달림플 2001, 페이지 205–221
- ^ Manhesa G, Allègre CJ, Dupréa B, Hamelin B (May 1980). "Lead isotope study of basic-ultrabasic layered complexes: Speculations about the age of the earth and primitive mantle characteristics". Earth and Planetary Science Letters. 47 (3): 370–382. Bibcode:1980E&PSL..47..370M. doi:10.1016/0012-821X(80)90024-2.
- ^ Schopf JW, Kudryavtsev AB, Czaja AD, Tripathi AB (5 October 2007). "Evidence of Archean life: Stromatolites and microfossils". Precambrian Research. 158 (3–4): 141–155. Bibcode:2007PreR..158..141S. doi:10.1016/j.precamres.2007.04.009.
- ^ 라벤 & 존슨 2002, 페이지 68
- ^ Baumgartner RJ, et al. (2019). "Nano−porous pyrite and organic matter in 3.5-billion-year-old stromatolites record primordial life" (PDF). Geology. 47 (11): 1039–1043. Bibcode:2019Geo....47.1039B. doi:10.1130/G46365.1. S2CID 204258554.
- ^ 생명의 초기 징후: 과학자들은 고대의 암석 Phys.org에서 미생물 유적을 발견한다. 2019년 9월 26일.
- ^ Jablonski D (25 June 1999). "The Future of the Fossil Record". Science. 284 (5423): 2114–2116. doi:10.1126/science.284.5423.2114. PMID 10381868. S2CID 43388925.
- ^ a b Cavalier-Smith T (29 June 2006). "Cell evolution and Earth history: stasis and revolution". Philosophical Transactions of the Royal Society B: Biological Sciences. 361 (1470): 969–1006. doi:10.1098/rstb.2006.1842. PMC 1578732. PMID 16754610.
- ^ Schopf JW (29 June 2006). "Fossil evidence of Archaean life". Philosophical Transactions of the Royal Society B: Biological Sciences. 361 (1470): 869–885. doi:10.1098/rstb.2006.1834. PMC 1578735. PMID 16754604.
- Altermann W, Kazmierczak J (November 2003). "Archean microfossils: a reappraisal of early life on Earth". Research in Microbiology. 154 (9): 611–617. doi:10.1016/j.resmic.2003.08.006. PMID 14596897.
- ^ Schopf JW (19 July 1994). "Disparate rates, differing fates: tempo and mode of evolution changed from the Precambrian to the Phanerozoic". Proceedings of the National Academy of Sciences of the United States of America. 91 (15): 6735–6742. Bibcode:1994PNAS...91.6735S. doi:10.1073/pnas.91.15.6735. PMC 44277. PMID 8041691.
- ^ Poole AM, Penny D (January 2007). "Evaluating hypotheses for the origin of eukaryotes". BioEssays. 29 (1): 74–84. doi:10.1002/bies.20516. PMID 17187354.
- ^ a b Dyall SD, Brown MT, Johnson PJ (9 April 2004). "Ancient Invasions: From Endosymbionts to Organelles". Science. 304 (5668): 253–257. Bibcode:2004Sci...304..253D. doi:10.1126/science.1094884. PMID 15073369. S2CID 19424594.
- ^ Martin W (October 2005). "The missing link between hydrogenosomes and mitochondria". Trends in Microbiology. 13 (10): 457–459. doi:10.1016/j.tim.2005.08.005. PMID 16109488.
- ^ Lang BF, Gray MW, Burger G (December 1999). "Mitochondrial genome evolution and the origin of eukaryotes". Annual Review of Genetics. 33: 351–397. doi:10.1146/annurev.genet.33.1.351. PMID 10690412.
- McFadden GI (1 December 1999). "Endosymbiosis and evolution of the plant cell". Current Opinion in Plant Biology. 2 (6): 513–519. doi:10.1016/S1369-5266(99)00025-4. PMID 10607659.
- ^ Hug, Laura A.; Baker, Brett J.; Anantharaman, Karthik; Brown, Christopher T.; Probst, Alexander J.; Castelle, Cindy J.; Butterfield, Cristina N.; Hernsdorf, Alex W.; Amano, Yuki; Ise, Kotaro; Suzuki, Yohey (11 April 2016). "A new view of the tree of life". Nature Microbiology. 1 (5): 16048. doi:10.1038/nmicrobiol.2016.48. ISSN 2058-5276. PMID 27572647.
- ^ Ciccarelli FD, Doerks T, von Mering C, et al. (3 March 2006). "Toward Automatic Reconstruction of a Highly Resolved Tree of Life". Science. 311 (5765): 1283–1287. Bibcode:2006Sci...311.1283C. CiteSeerX 10.1.1.381.9514. doi:10.1126/science.1123061. PMID 16513982. S2CID 1615592.
- ^ DeLong EF, Pace NR (1 August 2001). "Environmental Diversity of Bacteria and Archaea". Systematic Biology. 50 (4): 470–478. CiteSeerX 10.1.1.321.8828. doi:10.1080/106351501750435040. PMID 12116647.
- ^ Kaiser D (December 2001). "Building a multicellular organism". Annual Review of Genetics. 35: 103–123. doi:10.1146/annurev.genet.35.102401.090145. PMID 11700279. S2CID 18276422.
- ^ Zimmer C (7 January 2016). "Genetic Flip Helped Organisms Go From One Cell to Many". The New York Times. Retrieved 7 January 2016.
- ^ Valentine JW, Jablonski D, Erwin DH (1 March 1999). "Fossils, molecules and embryos: new perspectives on the Cambrian explosion". Development. 126 (5): 851–859. doi:10.1242/dev.126.5.851. PMID 9927587. Retrieved 30 December 2014.
- ^ Ohno S (January 1997). "The reason for as well as the consequence of the Cambrian explosion in animal evolution". Journal of Molecular Evolution. 44 (Suppl. 1): S23–S27. Bibcode:1997JMolE..44S..23O. doi:10.1007/PL00000055. PMID 9071008. S2CID 21879320.
- Valentine JW, Jablonski D (2003). "Morphological and developmental macroevolution: a paleontological perspective". The International Journal of Developmental Biology. 47 (7–8): 517–522. PMID 14756327. Retrieved 30 December 2014.
- ^ 캠벨, N. "생물학:개념 & 연결". 피어슨 교육. 샌프란시스코: 2003.
- ^ Harper, Douglas. "prokaryote". Online Etymology Dictionary.
- ^ Harper, Douglas. "eukaryotic". Online Etymology Dictionary.
- ^ Stanier RY, Van Niel CB (1962). "The concept of a bacterium". Archiv für Mikrobiologie. 42: 17–35. doi:10.1007/BF00425185. PMID 13916221. S2CID 29859498.
- ^ Woese CR, Fox GE (November 1977). "Phylogenetic structure of the prokaryotic domain: the primary kingdoms". Proceedings of the National Academy of Sciences of the United States of America. 74 (11): 5088–90. Bibcode:1977PNAS...74.5088W. doi:10.1073/pnas.74.11.5088. PMC 432104. PMID 270744.
- ^ Woese CR, Kandler O, Wheelis ML (June 1990). "Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya". Proceedings of the National Academy of Sciences of the United States of America. 87 (12): 4576–9. Bibcode:1990PNAS...87.4576W. doi:10.1073/pnas.87.12.4576. PMC 54159. PMID 2112744.
- ^ Ohtomo, Yoko; Kakegawa, Takeshi; Ishida, Akizumi; Nagase, Toshiro; Rosing, Minik T. (January 2014). "Evidence for biogenic graphite in early Archaean Isua metasedimentary rocks". Nature Geoscience. 7 (1): 25–28. Bibcode:2014NatGe...7...25O. doi:10.1038/ngeo2025.
- ^ Nutman, Allen P.; Bennett, Vickie C.; Friend, Clark R. L.; Kranendonk, Martin J. Van; Chivas, Allan R. (September 2016). "Rapid emergence of life shown by discovery of 3,700-million-year-old microbial structures". Nature. 537 (7621): 535–538. Bibcode:2016Natur.537..535N. doi:10.1038/nature19355. PMID 27580034. S2CID 205250494.
- ^ Borenstein, Seth (19 October 2015). "Hints of life on what was thought to be desolate early Earth". Excite. Yonkers, NY: Mindspark Interactive Network. Associated Press. Archived from the original on 23 October 2015. Retrieved 8 October 2018.
- ^ Bell, Elizabeth A.; Boehnike, Patrick; Harrison, T. Mark; et al. (19 October 2015). "Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon". Proc. Natl. Acad. Sci. U.S.A. 112 (47): 14518–21. Bibcode:2015PNAS..11214518B. doi:10.1073/pnas.1517557112. PMC 4664351. PMID 26483481.
- ^ Dodd, Matthew S.; Papineau, Dominic; Grenne, Tor; slack, John F.; Rittner, Martin; Pirajno, Franco; O'Neil, Jonathan; Little, Crispin T. S. (2 March 2017). "Evidence for early life in Earth's oldest hydrothermal vent precipitates" (PDF). Nature. 543 (7643): 60–64. Bibcode:2017Natur.543...60D. doi:10.1038/nature21377. PMID 28252057. S2CID 2420384.
- ^ Zimmer, Carl (1 March 2017). "Scientists Say Canadian Bacteria Fossils May Be Earth's Oldest". The New York Times. Retrieved 2 March 2017.
- ^ Nisbet, Euan G.; Fowler, C. M. R. (7 December 1999). "Archaean metabolic evolution of microbial mats". Proceedings of the Royal Society of London B. 266 (1436): 2375–2382. doi:10.1098/rspb.1999.0934. PMC 1690475.
- ^ Anbar, Ariel D.; Yun Duan; Lyons, Timothy W.; et al. (28 September 2007). "A Whiff of Oxygen Before the Great Oxidation Event?". Science. 317 (5846): 1903–1906. Bibcode:2007Sci...317.1903A. doi:10.1126/science.1140325. PMID 17901330. S2CID 25260892.
- ^ Knoll, Andrew H.; Javaux, Emmanuelle J.; Hewitt, David; Cohen, Phoebe (29 June 2006). "Eukaryotic organisms in Proterozoic oceans". Philosophical Transactions of the Royal Society B. 361 (1470): 1023–1038. doi:10.1098/rstb.2006.1843. PMC 1578724. PMID 16754612.
- ^ Fedonkin, Mikhail A. (31 March 2003). "The origin of the Metazoa in the light of the Proterozoic fossil record" (PDF). Paleontological Research. 7 (1): 9–41. doi:10.2517/prpsj.7.9. S2CID 55178329. Archived from the original (PDF) on 26 February 2009. Retrieved 2 September 2008.
- ^ Bonner, John Tyler (1998). "The origins of multicellularity". Integrative Biology. 1 (1): 27–36. doi:10.1002/(SICI)1520-6602(1998)1:1<27::AID-INBI4>3.0.CO;2-6.
- ^ 5월, R.M. (1988) "지상에는 몇 종이나 있을까?" 과학, 241 (4872): 1441–1449. doi:10.1126/과학.241.4872.1441
- ^ K.J. Locey와 J.T. Lennon은 "스케일링 법률은 전세계 미생물 다양성을 예측한다"고 말했다. 국립과학원 절차, 113(21): 5970–5975. doi:10.1073/pnas.1521291113.
- ^ 비토리노, L.C., 베사, L.A. (2018) "미생물 다양성: 추정치와 알려진 것의 차이" 다양성, 10(2): 46. doi:10.3390/d10020046.
 자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다.
자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다. - ^ 살아있는 박테리아는 지구의 기류를 타고 있다. Smithsonian Magazine, 2016년 1월 11일.
- ^ Robbins, Jim (13 April 2018). "Trillions Upon Trillions of Viruses Fall From the Sky Each Day". The New York Times. Retrieved 14 April 2018.
- ^ Reche, Isabel; D’Orta, Gaetano; Mladenov, Natalie; Winget, Danielle M; Suttle, Curtis A (29 January 2018). "Deposition rates of viruses and bacteria above the atmospheric boundary layer". ISME Journal. 12 (4): 1154–1162. doi:10.1038/s41396-017-0042-4. PMC 5864199. PMID 29379178.
- ^ Suttle, C.A. (2005). "Viruses in the Sea". Nature. 437 (9): 356–361. Bibcode:2005Natur.437..356S. doi:10.1038/nature04160. PMID 16163346. S2CID 4370363.
- ^ 2017년 8월 5일 자
- ^ 2017년 쇼르스, 페이지 593
- ^ Suttle CA (2007). "Marine viruses—major players in the global ecosystem". Nature Reviews Microbiology. 5 (10): 801–12. doi:10.1038/nrmicro1750. PMID 17853907. S2CID 4658457.
- ^ Schulz, Kestin; Smit, Mariya W.; Herfort, Lydie; Simon, Holly M. (2018). "The Unseen World in the River". Frontiers for Young Minds. 6. doi:10.3389/frym.2018.00004. S2CID 3344238. 자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다.
- ^ Stoeckenius W (October 1981). "Walsby's square bacterium: fine structure of an orthogonal procaryote". Journal of Bacteriology. 148 (1): 352–60. doi:10.1128/JB.148.1.352-360.1981. PMC 216199. PMID 7287626.
- ^ Durham, Bryndan P.; Grote, Jana; Whittaker, Kerry A.; Bender, Sara J.; Luo, Haiwei; Grim, Sharon L.; Brown, Julia M.; Casey, John R.; Dron, Antony; Florez-Leiva, Lennin; Krupke, Andreas; Luria, Catherine M.; Mine, Aric H.; Nigro, Olivia D.; Pather, Santhiska; Talarmin, Agathe; Wear, Emma K.; Weber, Thomas S.; Wilson, Jesse M.; Church, Matthew J.; Delong, Edward F.; Karl, David M.; Steward, Grieg F.; Eppley, John M.; Kyrpides, Nikos C.; Schuster, Stephan; Rappé, Michael S. (2014). "Draft genome sequence of marine alphaproteobacterial strain HIMB11, the first cultivated representative of a unique lineage within the Roseobacter clade possessing an unusually small genome". Standards in Genomic Sciences. 9 (3): 632–645. doi:10.4056/sigs.4998989. PMC 4148974. PMID 25197450.
- ^ Fredrickson JK, Zachara JM, Balkwill DL, Kennedy D, Li SM, Kostandarithes HM, Daly MJ, Romine MF, Brockman FJ (2004). "Geomicrobiology of high-level nuclear waste-contaminated vadose sediments at the Hanford site, Washington state". Applied and Environmental Microbiology. 70 (7): 4230–41. Bibcode:2004ApEnM..70.4230F. doi:10.1128/AEM.70.7.4230-4241.2004. PMC 444790. PMID 15240306.
- ^ Woese CR, Kandler O, Wheelis ML (1990). "Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya". Proceedings of the National Academy of Sciences of the United States of America. 87 (12): 4576–9. Bibcode:1990PNAS...87.4576W. doi:10.1073/pnas.87.12.4576. PMC 54159. PMID 2112744.
- ^ Schopf JW (1994). "Disparate rates, differing fates: tempo and mode of evolution changed from the Precambrian to the Phanerozoic". Proceedings of the National Academy of Sciences of the United States of America. 91 (15): 6735–42. Bibcode:1994PNAS...91.6735S. doi:10.1073/pnas.91.15.6735. PMC 44277. PMID 8041691.
- ^ DeLong EF, Pace NR (2001). "Environmental diversity of bacteria and archaea". Systematic Biology. 50 (4): 470–8. CiteSeerX 10.1.1.321.8828. doi:10.1080/106351501750435040. PMID 12116647.
- ^ Brown JR, Doolittle WF (1997). "Archaea and the prokaryote-to-eukaryote transition". Microbiology and Molecular Biology Reviews. 61 (4): 456–502. doi:10.1128/.61.4.456-502.1997. PMC 232621. PMID 9409149.
- ^ Poole AM, Penny D (2007). "Evaluating hypotheses for the origin of eukaryotes". BioEssays. 29 (1): 74–84. doi:10.1002/bies.20516. PMID 17187354. S2CID 36026766.
- ^ Lang BF, Gray MW, Burger G (1999). "Mitochondrial genome evolution and the origin of eukaryotes". Annual Review of Genetics. 33: 351–97. doi:10.1146/annurev.genet.33.1.351. PMID 10690412.
- ^ McFadden GI (1999). "Endosymbiosis and evolution of the plant cell". Current Opinion in Plant Biology. 2 (6): 513–9. doi:10.1016/S1369-5266(99)00025-4. PMID 10607659.
- ^ Koch AL (2002). "Control of the bacterial cell cycle by cytoplasmic growth". Critical Reviews in Microbiology. 28 (1): 61–77. doi:10.1080/1040-840291046696. PMID 12003041. S2CID 11624182.
- ^ Eagon RG (April 1962). "Pseudomonas natriegens, a marine bacterium with a generation time of less than 10 minutes". Journal of Bacteriology. 83 (4): 736–37. doi:10.1128/jb.83.4.736-737.1962. PMC 279347. PMID 13888946.
- ^ "캔디다투스 펠라지박터 우비크." 유럽 생물정보연구소. 2011년 유럽 생물정보학 연구소. 웹. 2012년 1월 8일. http://www.ebi.ac.uk/2can/genomes/bacteria/Candidatus_Pelagibacter_ubique.html 2008년 12월 1일 웨이백 머신에 보관
- ^ "Flea market: A newly discovered virus may be the most abundant organism on the planet". The Economist. 16 February 2013. Retrieved 16 February 2013.
- ^ Zhao, Y.; Temperton, B.; Thrash, J. C.; Schwalbach, M. S.; Vergin, K. L.; Landry, Z. C.; Ellisman, M.; Deerinck, T.; Sullivan, M. B.; Giovannoni, S. J. (2013). "Abundant SAR11 viruses in the ocean". Nature. 494 (7437): 357–360. Bibcode:2013Natur.494..357Z. doi:10.1038/nature11921. PMID 23407494. S2CID 4348619.
- ^ Bentzon-Tilia M, Gram L (2017). Bioprospecting. Topics in Biodiversity and Conservation. Springer, Cham. pp. 137–166. doi:10.1007/978-3-319-47935-4_7. ISBN 978-3-319-47933-0.
- ^ NCBI Taxonomy Browser: 로소박터 국립 생명공학 정보 센터 접근 가능: 2020년 5월 8일.
- ^ "The largest Bacterium: Scientist discovers new bacterial life form off the African coast", Max Planck Institute for Marine Microbiology, 8 April 1999, archived from the original on 20 January 2010
- ^ List of Prokaryotic names with Standing in Nomenclature - Genus Thiomargarita
- ^ 우리 바다의 산소 농도의 변화는 근본적인 생물학적 순환을 방해할 수 있다. Phys.org, 2019년 11월 25일.
- ^ Walsh PJ, Smith S, Fleming L, Solo-Gabriele H, Gerwick WH, eds. (2 September 2011). "Cyanobacteria and cyanobacterial toxins". Oceans and Human Health: Risks and Remedies from the Seas. Academic Press. pp. 271–296. ISBN 978-0-08-087782-2.
- ^ "The Rise of Oxygen - Astrobiology Magazine". Astrobiology Magazine. 30 July 2003. Retrieved 6 April 2016.
- ^ Flannery, D. T.; R.M. Walter (2012). "Archean tufted microbial mats and the Great Oxidation Event: new insights into an ancient problem". Australian Journal of Earth Sciences. 59 (1): 1–11. Bibcode:2012AuJES..59....1F. doi:10.1080/08120099.2011.607849. S2CID 53618061.
- ^ Rothschild, Lynn (September 2003). "Understand the evolutionary mechanisms and environmental limits of life". NASA. Archived from the original on 11 March 2012. Retrieved 13 July 2009.
- ^ 바수, Subhajit, Gledhill, 마사, 드 비어, 디르크;PrabhuMatondkar, S.G.;Shaked, Yeala(2019년)."관련된 박테리아로 해양 cyanobacteria Trichodesmium 식민지가 상호 작용하여 먼지에서 철을 획득하기 위해".통신 생물학.2:284.doi:10.1038/s42003-019-0534-z.PMC6677733.PMID 31396564.재료는 창조적 공용 귀인 4.0국제 라이센스 하에 가능하다 이 원본에서 복사되었다.
- ^ Patrick J. Keeling (2004). "Diversity and evolutionary history of plastids and their hosts". American Journal of Botany. 91 (10): 1481–1493. doi:10.3732/ajb.91.10.1481. PMID 21652304.
- ^ Nadis S (December 2003). "The cells that rule the seas" (PDF). Scientific American. 289 (6): 52–3. Bibcode:2003SciAm.289f..52N. doi:10.1038/scientificamerican1203-52. PMID 14631732. Archived from the original (PDF) on 19 April 2014. Retrieved 2 June 2019.
- ^ "The Most Important Microbe You've Never Heard Of". npr.org.
- ^ Flombaum, P.; Gallegos, J. L.; Gordillo, R. A.; Rincon, J.; Zabala, L. L.; Jiao, N.; Karl, D. M.; Li, W. K. W.; Lomas, M. W.; Veneziano, D.; Vera, C. S.; Vrugt, J. A.; Martiny, A. C. (2013). "Present and future global distributions of the marine Cyanobacteria Prochlorococcus and Synechococcus". Proceedings of the National Academy of Sciences. 110 (24): 9824–9829. Bibcode:2013PNAS..110.9824F. doi:10.1073/pnas.1307701110. PMC 3683724. PMID 23703908.
- ^ Nabors, Murray W. (2004). Introduction to Botany. San Francisco, CA: Pearson Education, Inc. ISBN 978-0-8053-4416-5.
- ^ Allaby, M., ed. (1992). "Algae". The Concise Dictionary of Botany. Oxford: Oxford University Press.
- ^ Ivars-Martínez, Elena; d'Auria, Giuseppe; Rodríguez-Valera, Francisco; Sánchez-Porro, Cristina; Ventosa, Antonio; Joint, IAN; Mühling, Martin (2008). "Biogeography of the ubiquitous marine bacterium Alteromonas macleodiidetermined by multilocus sequence analysis". Molecular Ecology. 17 (18): 4092–4106. doi:10.1111/j.1365-294x.2008.03883.x. PMID 19238708. S2CID 38830049.
- ^ López-Pérez, Mario; Gonzaga, Aitor; Martin-Cuadrado, Ana-Belen; Onyshchenko, Olga; Ghavidel, Akbar; Ghai, Rohit; Rodriguez-Valera, Francisco (2012). "Genomes of surface isolates of Alteromonas macleodii: The life of a widespread marine opportunistic copiotroph". Scientific Reports. 2: 696. Bibcode:2012NatSR...2E.696L. doi:10.1038/srep00696. PMC 3458243. PMID 23019517.
- ^ Bang C, Schmitz RA (2015). "Archaea associated with human surfaces: not to be underestimated". FEMS Microbiology Reviews. 39 (5): 631–48. doi:10.1093/femsre/fuv010. PMID 25907112.
- ^ 고고학 온라인 어원 사전. 2016년 8월 17일 회수
- ^ Pace NR (May 2006). "Time for a change". Nature. 441 (7091): 289. Bibcode:2006Natur.441..289P. doi:10.1038/441289a. PMID 16710401. S2CID 4431143.
- ^ Stoeckenius W (1 October 1981). "Walsby's square bacterium: fine structure of an orthogonal procaryote". Journal of Bacteriology. 148 (1): 352–60. doi:10.1128/JB.148.1.352-360.1981. PMC 216199. PMID 7287626.
- ^ Madigan M, Martinko J, eds. (2005). Brock Biology of Microorganisms (11th ed.). Prentice Hall. ISBN 978-0-13-144329-7.
- ^ Huber, Harald; et al. (2002). "A new phylum of Archaea represented by a nanosized hyperthermophilic symbiont". Nature. 417 (6884): 63–67. Bibcode:2002Natur.417...63H. doi:10.1038/417063a. PMID 11986665. S2CID 4395094.
- ^ Brochier, Celine; Gribaldo, S; Zivanovic, Y; Confalonieri, F; et al. (2005). "Nanoarchaea: representatives of a novel archaeal phylum or a fast-evolving euryarchaeal lineage related to Thermococcales?". Genome Biology. 6 (5): R42. doi:10.1186/gb-2005-6-5-r42. PMC 1175954. PMID 15892870.
- ^ Orellana, Luis H.; Ben Francis, T.; Krüger, Karen; Teeling, Hanno; Müller, Marie-Caroline; Fuchs, Bernhard M.; Konstantinidis, Konstantinos T.; Amann, Rudolf I. (2019). "Niche differentiation among annually recurrent coastal Marine Group II Euryarchaeota". The ISME Journal. 13 (12): 3024–3036. doi:10.1038/s41396-019-0491-z. PMC 6864105. PMID 31447484.
- ^ . 4니시무라, 요수 케;Watai, 히로야스, 혼다, 다카시, 미하라, 도모코, Omae, Kimiho, 루, 사이먼, Blanc-Mathieu, 로맹, 야마모토 Keigo, Hingamp, 파스칼, Sako, 요시히코. Sullivan, 매튜 B에, 특히 그림 참조하십시오.고토, 스스무, 오가타, 히로유키, 요시다, 다카시(2017년)."환경 바이러스 유전체 비 자유 주의파 Virus-Host 상호 작용에 해양에". mSphere.2(2). doi:10.1128/mSphere.00359-16.PMC5332604.PMID 28261669.
- ^ Philosof, Alon; Yutin, Natalya; Flores-Uribe, José; Sharon, Itai; Koonin, Eugene V.; Béjà, Oded (2017). "Novel Abundant Oceanic Viruses of Uncultured Marine Group II Euryarchaeota". Current Biology. 27 (9): 1362–1368. doi:10.1016/j.cub.2017.03.052. PMC 5434244. PMID 28457865.
- ^ Xia, Xiaomin; Guo, Wang; Liu, Hongbin (2017). "Basin Scale Variation on the Composition and Diversity of Archaea in the Pacific Ocean". Frontiers in Microbiology. 8: 2057. doi:10.3389/fmicb.2017.02057. PMC 5660102. PMID 29109713.
- ^ Martin-Cuadrado, Ana-Belen; Garcia-Heredia, Inmaculada; Moltó, Aitor Gonzaga; López-Úbeda, Rebeca; Kimes, Nikole; López-García, Purificación; Moreira, David; Rodriguez-Valera, Francisco (2015). "A new class of marine Euryarchaeota group II from the mediterranean deep chlorophyll maximum". The ISME Journal. 9 (7): 1619–1634. doi:10.1038/ismej.2014.249. PMC 4478702. PMID 25535935.
- ^ 엔씨비: 칸디다투스 포세이디데인(주문)
- ^ 엔씨비: 마린 그룹 3
- ^ NCBI: 마린 그룹 4
- ^ Zillig W (December 1991). "Comparative biochemistry of Archaea and Bacteria". Current Opinion in Genetics & Development. 1 (4): 544–51. doi:10.1016/S0959-437X(05)80206-0. PMID 1822288.
- ^ Slonczewski JL, Foster JW. Microbiology: An Evolving Science (3 ed.). WW Norton & Company. pp. 491–44.
- ^ Hogan CM (2010). "Extremophile". In Monosson E, Cleveland C (eds.). Encyclopedia of Earth. National Council of Science & the Environment.
- ^ Yurkov, V. V.; Beatty, J. T. (1998). "Aerobic anoxygenic phototrophic bacteria". Microbiology and Molecular Biology Reviews. 62 (3): 695–724. doi:10.1128/MMBR.62.3.695-724.1998. PMC 98932. PMID 9729607.
- ^ Lami, R.; Cottrell, M. T.; Ras, J.; Ulloa, O.; Obernosterer, I.; Claustre, H.; Kirchman, D. L.; Lebaron, P. (2007). "High Abundances of Aerobic Anoxygenic Photosynthetic Bacteria in the South Pacific Ocean". Applied and Environmental Microbiology. 73 (13): 4198–205. Bibcode:2007ApEnM..73.4198L. doi:10.1128/AEM.02652-06. PMC 1932784. PMID 17496136.
- ^ 생명과학 주간. (2012). 박테리아; 스페인 국립연구위원회(CSIC)의 보고 박테리아의 최근 진보를 설명한다. ISSN 1552-2466. 페이지 4582.
- ^ Jiao, Nianzhi; Zhang, Yao; Zeng, Yonghui; Hong, Ning; Liu, Rulong; Chen, Feng; Wang, Pinxian (2007). "Distinct distribution pattern of abundance and diversity of aerobic anoxygenic phototrophic bacteria in the global ocean". Environmental Microbiology. 9 (12): 3091–3099. doi:10.1111/j.1462-2920.2007.01419.x. PMID 17991036.
- ^ Lami, Raphaël; Cottrell, Matthew T.; Ras, JoséPhine; Ulloa, Osvaldo; Obernosterer, Ingrid; Claustre, Hervé; Kirchman, David L.; Lebaron, Philippe (2007). "High Abundances of Aerobic Anoxygenic Photosynthetic Bacteria in the South Pacific Ocean". Applied and Environmental Microbiology. 73 (13): 4198–4205. Bibcode:2007ApEnM..73.4198L. doi:10.1128/AEM.02652-06. PMC 1932784. PMID 17496136.
- ^ Kolber, Z. S. (2001). "Contribution of Aerobic Photoheterotrophic Bacteria to the Carbon Cycle in the Ocean". Science. 292 (5526): 2492–2495. doi:10.1126/science.1059707. PMID 11431568. S2CID 1970984.
- ^ Aragno M, Schlegel HG (1981). "The Hydrogen-Oxidizing Bacteria". In Starr MP, Stolp H, Trüper HG, Balows A, Schlegel HG (eds.). The Prokaryotes. Berlin, Heidelberg: Springer. pp. 865–893. doi:10.1007/978-3-662-13187-9_70. ISBN 978-3-662-13187-9.
- ^ a b Albers SV, Jarrell KF (27 January 2015). "The archaellum: how Archaea swim". Frontiers in Microbiology. 6: 23. doi:10.3389/fmicb.2015.00023. PMC 4307647. PMID 25699024.
- ^ Silverman M, Simon M (May 1974). "Flagellar rotation and the mechanism of bacterial motility". Nature. 249 (452): 73–4. Bibcode:1974Natur.249...73S. doi:10.1038/249073a0. PMID 4598030. S2CID 10370084.
- ^ Meister GL, Berg HC (1987). "Rapid rotation of flagellar bundles in swimming bacteria". Nature. 325 (6105): 637–640. Bibcode:1987Natur.325..637L. doi:10.1038/325637a0. S2CID 4242129.
- ^ Berg HC, Anderson RA (October 1973). "Bacteria swim by rotating their flagellar filaments". Nature. 245 (5425): 380–2. Bibcode:1973Natur.245..380B. doi:10.1038/245380a0. PMID 4593496. S2CID 4173914.
- ^ Jahn TL, Bovee EC (1965). "Movement and locomotion of microorganisms". Annual Review of Microbiology. 19: 21–58. doi:10.1146/annurev.mi.19.100165.000321. PMID 5318439.
- ^ Harshey RM (2003). "Bacterial motility on a surface: many ways to a common goal". Annual Review of Microbiology. 57: 249–73. doi:10.1146/annurev.micro.57.030502.091014. PMID 14527279.
- ^ Jarrell K (2009). "Archaeal Flagella and Pili". Pili and Flagella: Current Research and Future Trends. Caister Academic Press. ISBN 978-1-904455-48-6.
- ^ Brady, Richard M. (1993). "Torque and switching in the bacterial flagellar motor. An electrostatic model". Biophysical Journal. 64 (4): 961–973. Bibcode:1993BpJ....64..961B. doi:10.1016/S0006-3495(93)81462-0. PMC 1262414. PMID 7684268.
- ^ Streif S, Staudinger WF, Marwan W, Oesterhelt D (2008). "Flagellar rotation in the archaeon Halobacterium salinarum depends on ATP". Journal of Molecular Biology. 384 (1): 1–8. doi:10.1016/j.jmb.2008.08.057. PMID 18786541.
- ^ a b Skerker, J. M.; Berg, H. C. (5 June 2001). "Direct observation of extension and retraction of type IV pili". Proceedings of the National Academy of Sciences of the United States of America. 98 (12): 6901–6904. Bibcode:2001PNAS...98.6901S. doi:10.1073/pnas.121171698. ISSN 0027-8424. PMC 34450. PMID 11381130.
- ^ Mattick, John S. (2002). "Type IV pili and twitching motility". Annual Review of Microbiology. 56: 289–314. doi:10.1146/annurev.micro.56.012302.160938. ISSN 0066-4227. PMID 12142488.
- ^ Merz, A. J.; So, M.; Sheetz, M. P. (7 September 2000). "Pilus retraction powers bacterial twitching motility". Nature. 407 (6800): 98–102. Bibcode:2000Natur.407...98M. doi:10.1038/35024105. ISSN 0028-0836. PMID 10993081. S2CID 4425775.
- ^ Henrichsen, J. (December 1972). "Bacterial surface translocation: a survey and a classification". Bacteriological Reviews. 36 (4): 478–503. doi:10.1128/BR.36.4.478-503.1972. ISSN 0005-3678. PMC 408329. PMID 4631369.
- ^ Nan, Beiyan (February 2017). "Bacterial Gliding Motility: Rolling Out a Consensus Model". Current Biology. 27 (4): R154–R156. doi:10.1016/j.cub.2016.12.035. PMID 28222296.
- ^ Nan, Beiyan; McBride, Mark J.; Chen, Jing; Zusman, David R.; Oster, George (February 2014). "Bacteria that Glide with Helical Tracks". Current Biology. 24 (4): 169–174. doi:10.1016/j.cub.2013.12.034. PMC 3964879. PMID 24556443.
- ^ Sibley, L.David; Håkansson, Sebastian; Carruthers, Vern B (1 January 1998). "Gliding motility: An efficient mechanism for cell penetration". Current Biology. 8 (1): R12–R14. doi:10.1016/S0960-9822(98)70008-9. PMID 9427622. S2CID 17555804.
- ^ Sibley, LDI (October 2010). "How apicomplexan parasites move in and out of cells". Curr Opin Biotechnol. 21 (5): 592–8. doi:10.1016/j.copbio.2010.05.009. PMC 2947570. PMID 20580218.
- ^ Harshey, Rasika M. (1 January 2003). "Bacterial Motility on a Surface: Many Ways to a Common Goal". Annual Review of Microbiology. 57 (1): 249–73. doi:10.1146/annurev.micro.57.030502.091014. PMID 14527279.
- ^ Henrichsen, J (1972). "Bacterial surface translocation: a survey and a classification". Bacteriological Reviews. 36 (4): 478–503. doi:10.1128/BR.36.4.478-503.1972. PMC 408329. PMID 4631369.
- ^ "BIOL 230 Lab Manual: Nonmotile Bacteria in Motility Medium". faculty.ccbcmd.edu.
- ^ Possfai, M, Lefére, C, Trubitsyn, D, Bazylinski, D.A., Frankel, R.(2013) "자석 광물의 구성과 결정 형태학의 유전학적 중요성" 미생물학의 프론티어, 4: 344. doi:10.3389/fmicb.2013.00344. 자료는 이 출처에서 복사되었으며, Creative Commons Attribution 3.0 International License에 따라 이용할 수 있다.
- ^ Lin, Wei; Zhang, Wensi; Zhao, Xiang; Roberts, Andrew; Paterson, Greig; Bazylinski, Dennis; Pan, Yongxin (March 2018). "Genomic expansion of magnetotactic bacteria reveals an early common origin of magnetotaxis with lineage-specific evolution". The ISME Journal. 12 (6): 1508–1519. doi:10.1038/s41396-018-0098-9. PMC 5955933. PMID 29581530.
- ^ a b Dusenbery, David B. (2009). Living at micro scale : the unexpected physics of being small. Cambridge, Mass.: Harvard University Press. pp. 100–101. ISBN 978-0-674-03116-6.
- ^ a b W.J. 장과 L.F. 우, 2020년 해양자석세균의 플라젤라와 수영행태 생체 분자, 10(3), p.460. doi:10.3390/biom10030460. 자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다.
- ^ Jogler, C.; Wanner, G.; Kolinko, S.; Niebler, M.; Amann, R.; Petersen, N.; Kube, M.; Reinhardt, R.; Schuler, D. (2010). "Conservation of proteobacterial magnetosome genes and structures in an uncultivated member of the deep-branching Nitrospira phylum". Proceedings of the National Academy of Sciences. 108 (3): 1134–1139. Bibcode:2011PNAS..108.1134J. doi:10.1073/pnas.1012694108. PMC 3024689. PMID 21191098.
- ^ Monteil, Caroline L.; Vallenet, David; Menguy, Nicolas; Benzerara, Karim; Barbe, Valérie; Fouteau, Stéphanie; Cruaud, Corinne; Floriani, Magali; Viollier, Eric; Adryanczyk, Géraldine; Leonhardt, Nathalie (July 2019). "Ectosymbiotic bacteria at the origin of magnetoreception in a marine protist". Nature Microbiology. 4 (7): 1088–1095. doi:10.1038/s41564-019-0432-7. ISSN 2058-5276. PMC 6697534. PMID 31036911.
- ^ Natan, Eviatar; Fitak, Robert Rodgers; Werber, Yuval; Vortman, Yoni (28 September 2020). "Symbiotic magnetic sensing: raising evidence and beyond". Philosophical Transactions of the Royal Society B: Biological Sciences. 375 (1808): 20190595. doi:10.1098/rstb.2019.0595. PMC 7435164. PMID 32772668.
- ^ Kopp, R. E. & Kirschvink, J. L. (2007). "The identification and biogeochemical interpretation of fossil magnetotactic bacteria" (PDF). Earth-Science Reviews. 86 (1–4): 42–61. Bibcode:2008ESRv...86...42K. doi:10.1016/j.earscirev.2007.08.001.
- ^ Chang, S. R. & J. L. Kirschvink (1989). "Magnetofossils, the magnetization of sediments, and the evolution of magnetite biomineralization". Annual Review of Earth and Planetary Sciences. 17: 169–195. Bibcode:1989AREPS..17..169C. doi:10.1146/annurev.ea.17.050189.001125.
- ^ Walsby AE (1969). "The Permeability of Blue-Green Algal Gas-Vacuole Membranes to Gas". Proceedings of the Royal Society of London. Series B, Biological Sciences. 173 (1031): 235–255. Bibcode:1969RSPSB.173..235W. doi:10.1098/rspb.1969.0049. JSTOR 75817. OCLC 479422015. S2CID 95321956.
- ^ Kalanetra KM, Huston SL, Nelson DC (December 2004). "Novel, attached, sulfur-oxidizing bacteria at shallow hydrothermal vents possess vacuoles not involved in respiratory nitrate accumulation". Applied and Environmental Microbiology. 70 (12): 7487–96. Bibcode:2004ApEnM..70.7487K. doi:10.1128/AEM.70.12.7487-7496.2004. PMC 535177. PMID 15574952.
- ^ Schulz-Vogt HN (2006). "Vacuoles". Inclusions in Prokaryotes. Microbiology Monographs. 1. pp. 295–298. doi:10.1007/3-540-33774-1_10. ISBN 978-3-540-26205-3.
- ^ Montánchez, Itxaso; Ogayar, Elixabet; Plágaro, Ander Hernández; Esteve-Codina, Anna; Gómez-Garrido, Jèssica; Orruño, Maite; Arana, Inés; Kaberdin, Vladimir R. (2019). "Analysis of Vibrio harveyi adaptation in sea water microcosms at elevated temperature provides insights into the putative mechanisms of its persistence and spread in the time of global warming". Scientific Reports. 9 (1): 289. Bibcode:2019NatSR...9..289M. doi:10.1038/s41598-018-36483-0. PMC 6343004. PMID 30670759. S2CID 58950215.
- ^ McFall-Ngai, Margaret; Heath-Heckman, Elizabeth A.C.; Gillette, Amani A.; Peyer, Suzanne M.; Harvie, Elizabeth A. (2012). "The secret languages of coevolved symbioses: Insights from the Euprymna scolopes–Vibrio fischeri symbiosis". Seminars in Immunology. 24 (1): 3–8. doi:10.1016/j.smim.2011.11.006. PMC 3288948. PMID 22154556.
- ^ Waters, Christopher M.; Bassler, Bonnie L. (7 October 2005). "QUORUM SENSING: Cell-to-Cell Communication in Bacteria". Annual Review of Cell and Developmental Biology. 21 (1): 319–346. doi:10.1146/annurev.cellbio.21.012704.131001. PMID 16212498.
- ^ Young, R.; Roper, C. (1976). "Bioluminescent countershading in midwater animals: Evidence from living squid". Science. 191 (4231): 1046–1048. Bibcode:1976Sci...191.1046Y. doi:10.1126/science.1251214. PMID 1251214.
- ^ Owens, Leigh; Busico-Salcedo, Nancy (2006). "Vibrio harveyi: Pretty Problems in Paradise (Chapter 19)". In Thompson, Fabiano; Austin, Brian; Swings, Jean (eds.). The Biology of Vibrios. ASM Press.
- ^ DeLong, E.F.; Beja, O. (2010). "The light-driven proton pump proteorhodopsin enhances bacterial survival during tough times". PLOS Biology. 8 (4): e1000359. doi:10.1371/journal.pbio.1000359. PMC 2860490. PMID 20436957. e1000359.
- ^ a b Gómez-Consarnau, L.; Raven, J.A.; Levine, N.M.; Cutter, L.S.; Wang, D.; Seegers, B.; Arístegui, J.; Fuhrman, J.A.; Gasol, J.M.; Sañudo-Wilhelmy, S.A. (2019). "Microbial rhodopsins are major contributors to the solar energy captured in the sea". Science Advances. 5 (8): eaaw8855. Bibcode:2019SciA....5.8855G. doi:10.1126/sciadv.aaw8855. PMC 6685716. PMID 31457093.
- ^ Oren, Aharon (2002). "Molecular ecology of extremely halophilic Archaea and Bacteria". FEMS Microbiology Ecology. 39 (1): 1–7. doi:10.1111/j.1574-6941.2002.tb00900.x. PMID 19709178.
- ^ Béja, O.; Aravind, L.; Koonin, E.V.; Suzuki, M.T.; Hadd, A.; Nguyen, L.P.; Jovanovich, S.B.; Gates, C.M.; Feldman, R.A.; Spudich, J.L.; Spudich, E.N. (2000). "Bacterial rhodopsin: evidence for a new type of phototrophy in the sea". Science. 289 (5486): 1902–1906. Bibcode:2000Sci...289.1902B. doi:10.1126/science.289.5486.1902. PMID 10988064. S2CID 1461255.
- ^ "Interviews with Fellows: Ed Delong". American Academy of Microbiology. Archived from the original on 7 August 2016. Retrieved 2 July 2016.
- ^ 배터리와 함께 하는 박테리아, 대중 과학, 2001년 1월, 55페이지.
- ^ Béja, O.; Aravind, L.; Koonin, E.V.; Suzuki, M.T.; Hadd, A.; Nguyen, L.P.; Jovanovich, S.B.; Gates, C.M.; Feldman, R.A.; Spudich, J.L.; Spudich, E.N. (2000). "Bacterial rhodopsin: evidence for a new type of phototrophy in the sea". Science. 289 (5486): 1902–1906. Bibcode:2000Sci...289.1902B. doi:10.1126/science.289.5486.1902. PMID 10988064.
- ^ Boeuf, Dominique; Audic, Stéphane; Brillet-Guéguen, Loraine; Caron, Christophe; Jeanthon, Christian (2015). "MicRhoDE: a curated database for the analysis of microbial rhodopsin diversity and evolution". Database. 2015: bav080. doi:10.1093/database/bav080. PMC 4539915. PMID 26286928.
- ^ Yawo, Hiromu; Kandori, Hideki; Koizumi, Amane (5 June 2015). Optogenetics: Light-Sensing Proteins and Their Applications. Springer. pp. 3–4. ISBN 978-4-431-55516-2. Retrieved 30 September 2015.
- ^ 2019년 8월 8일 남캘리포니아대학 프레스룸에서 작은 해양 마이크로베가 기후변화 대학에서 큰 역할을 할 수 있을 것이다.
- ^ DasSarma, Shiladitya; Schwieterman, Edward W. (11 October 2018). "Early evolution of purple retinal pigments on Earth and implications for exoplanet biosignatures". International Journal of Astrobiology. 20 (3): 241–250. arXiv:1810.05150. doi:10.1017/S1473550418000423. S2CID 119341330.
- ^ Sparks, William B.; DasSarma, S.; Reid, I. N. (December 2006). "Evolutionary Competition Between Primitive Photosynthetic Systems: Existence of an early purple Earth?". American Astronomical Society Meeting Abstracts. 38: 901. Bibcode:2006AAS...209.0605S.
- ^ Dane Konop (29 July 1997). "Scientists discover methane ice worms on Gulf of Mexico sea floor". National Oceanic and Atmospheric Administration. Archived from the original on 9 June 2010. Retrieved 22 January 2010.
- ^ Sharp, Koty H.; Pratte, Zoe A.; Kerwin, Allison H.; Rotjan, Randi D.; Stewart, Frank J. (2017). "Season, but not symbiont state, drives microbiome structure in the temperate coral Astrangia poculata". Microbiome. 5 (1): 120. doi:10.1186/s40168-017-0329-8. PMC 5603060. PMID 28915923.
- ^ K.A. 레마, 윌리스, B.L., 본, D.G. (2012) "산호는 공생 질소 고정 박테리아와 특징적인 연관성을 형성한다." 적용 및 환경 미생물학, 78(9): 3136-3144. doi:10.1128/AEM.07800-11.
- ^ Petersen, Jillian M.; Frank U. Zielinski; Thomas Pape; Richard Seifert; Cristina Moraru; et al. (11 August 2011). "Hydrogen is an energy source for hydrothermal vent symbioses". Nature. 476 (7359): 176–180. Bibcode:2011Natur.476..176P. doi:10.1038/nature10325. PMID 21833083. S2CID 25578.

- ^ Kleiner, Manuel; Wentrup, Cecilia; Lott, Christian; Teeling, Hanno; Wetzel, Silke; Young, Jacque; Chang, Yun-Juan; Shah, Manesh; VerBerkmoes, Nathan C. (8 May 2012). "Metaproteomics of a gutless marine worm and its symbiotic microbial community reveal unusual pathways for carbon and energy use". Proceedings of the National Academy of Sciences of the United States of America. 109 (19): E1173–E1182. doi:10.1073/pnas.1121198109. PMC 3358896. PMID 22517752.
- ^ Woyke, Tanja; Teeling, Hanno; Ivanova, Natalia N.; Huntemann, Marcel; Richter, Michael; Gloeckner, Frank Oliver; Boffelli, Dario; Anderson, Iain J.; Barry, Kerrie W. (26 October 2006). "Symbiosis insights through metagenomic analysis of a microbial consortium" (PDF). Nature. 443 (7114): 950–955. Bibcode:2006Natur.443..950W. doi:10.1038/nature05192. PMID 16980956. S2CID 140106758.
- ^ USGS 과학자들은 프로브와 프라이머 디자인을 위한 지역사회 자원을 제공하는 온대 산호로부터 긴 시간 동안 읽혀진 마이크로바이옴 시퀀스를 발간했다. 미국 지질조사국, 2019년 3월 6일. 자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다.
- ^ B. 골드 스미스 Dawn;aPratte, 조이는;a켈로그, 크리스티나 약혼녀와의Snader, 새라, h.샤프 Koty(2019년)."온대 산호 천해 poculata microbiome의 안정성 다른 염기 서열 분석 방법론을 가로질러 반영된다".AIMS미생물학. 5(1):62–76. doi:10.3934/microbiol.2019.1.62. 1.6646935.PMID 31384703.재료는 창조적 공용 귀인 4.0국제 라이센스 하에 가능하다 이 원본에서 복사되었다.
- ^ a b 루이즈곤잘레스, C, 시모, R, 소마루가, R, 가솔, J.M. (2013) "어둠에서 멀어짐: 태양 방사선이 헤테로테틱 박테리오플랑크톤 활동에 미치는 영향에 대한 검토" 미생물학의 프론티어, 4: 131. doi:10.3389/fmicb.2013.00131. 자료는 이 출처에서 복사되었으며, Creative Commons Attribution 3.0 International License에 따라 이용할 수 있다.
- ^ 워커, J. C. G. (1980). 자연환경에서의 산소순환과 생물화학순환. 베를린: 스프링거-베를라크.
- ^ Longhurst, A, Sathyendranath, S, Platt, T, Caverhill, C. (1995년) 위성 방사선계 데이터를 통한 해양에서의 세계 1차 생산 추정치. J. 플랑크톤 레스 17, 1245–1271.
- ^ a b c d e 쿠라타, N, 벨라, K, 해밀턴, B, 시브지, M, 솔로비예프, A, 맷, S, 타르타르, A., 그리고 W. (2016) "대양의 거의 표면층에 있는 항산화 박테리아" 자연: 과학 보고서, 6(1): 1–8. doi:10.1038/srep19123. 자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다.
- ^ 슈티치, 브이, 조소비치, B, 마르첸코, E, 비하리, N., 크르시니치, F.(1981) "해양 식물성 플랑크톤에 의한 계면활성제 생산" 해양화학, 10(6): 505–520. doi:10.1016/0304-4203(81)90004-9.
- ^ a b 시겔이 만들어 낸 데이비드 A.;Buesseler, 켄 O;Behrenfeld, 마이클 J.;Benitez-Nelson, 클라우디아 R., 간수장님, 엠마뉴엘, 브레진스키,인 MarkA.;Burd, 아드리안, 칼슨, 크레이그 a.;d'Asaro, 에릭 A.,.Doney, 스콧 C;페리, 메리 j가, 스탠리, Rachel은 H.R.;스타인 버그, 데보라 K.(2016년)."예측의 수출과 운명은 글로벌 해양 네트워크 총생산:.그 EXPORTS 과학 Plan".해양 과학에 국경. 3.doi:10.3389/fmars.2016.00022.재료는 창조적 공용 귀인 4.0국제 라이센스 하에 가능하다 이 원본에서 복사되었다.
- ^ Liu X, Pan J, Liu Y, Li M, Gu JD (October 2018). "Diversity and distribution of Archaea in global estuarine ecosystems". The Science of the Total Environment. 637–638: 349–358. Bibcode:2018ScTEn.637..349L. doi:10.1016/j.scitotenv.2018.05.016. PMID 29753224.
- ^ Cabello P, Roldán MD, Moreno-Vivián C (November 2004). "Nitrate reduction and the nitrogen cycle in archaea". Microbiology. 150 (Pt 11): 3527–46. doi:10.1099/mic.0.27303-0. PMID 15528644.
- ^ Mehta MP, Baross JA (December 2006). "Nitrogen fixation at 92 degrees C by a hydrothermal vent archaeon". Science. 314 (5806): 1783–86. Bibcode:2006Sci...314.1783M. doi:10.1126/science.1134772. PMID 17170307. S2CID 84362603.
- ^ Francis CA, Beman JM, Kuypers MM (May 2007). "New processes and players in the nitrogen cycle: the microbial ecology of anaerobic and archaeal ammonia oxidation". The ISME Journal. 1 (1): 19–27. doi:10.1038/ismej.2007.8. PMID 18043610.
- ^ Coolen MJ, Abbas B, van Bleijswijk J, Hopmans EC, Kuypers MM, Wakeham SG, Sinninghe Damsté JS, et al. (April 2007). "Putative ammonia-oxidizing Crenarchaeota in suboxic waters of the Black Sea: a basin-wide ecological study using 16S ribosomal and functional genes and membrane lipids". Environmental Microbiology. 9 (4): 1001–16. doi:10.1111/j.1462-2920.2006.01227.x. hdl:1912/2034. PMID 17359272.
- ^ Baker BJ, Banfield JF (May 2003). "Microbial communities in acid mine drainage". FEMS Microbiology Ecology. 44 (2): 139–52. doi:10.1016/S0168-6496(03)00028-X. PMID 19719632.
- ^ Schimel J (August 2004). "Playing scales in the methane cycle: from microbial ecology to the globe". Proceedings of the National Academy of Sciences of the United States of America. 101 (34): 12400–01. Bibcode:2004PNAS..10112400S. doi:10.1073/pnas.0405075101. PMC 515073. PMID 15314221.
참고 문헌 목록
- Dalrymple, G. Brent (2001). "The age of the Earth in the twentieth century: a problem (mostly) solved". Special Publications, Geological Society of London. 190 (1): 205–221. Bibcode:2001GSLSP.190..205D. doi:10.1144/GSL.SP.2001.190.01.14. S2CID 130092094.
- Raven, Peter Hamilton; Johnson, George Brooks (2002). Biology. McGraw-Hill Education. ISBN 978-0-07-112261-0.
- Shors, Teri (2017). Understanding viruses. Jones and Bartlett Publishers. ISBN 978-1284025927.




{kind=link}