해상 1차 생산
Marine primary production
해양 1차 생산은 해양에서 대기 또는 용해된 이산화탄소에서 유기 화합물을 합성하는 것이다.주로 빛을 에너지원으로 사용하는 광합성 과정을 거치지만 무기화합물의 산화나 환원을 에너지원으로 사용하는 화학합성을 통해서도 발생한다.지구상의 거의 모든 생명체는 1차 생산에 직간접적으로 의존하고 있다.1차 생산을 담당하는 유기체는 1차 생산자 또는 자가영양체라고 불린다.
대부분의 해양 1차 생산은 조류와 시아노박테리아라고 불리는 다양한 해양 미생물에 의해 생성된다.이들은 모두 해양 먹이사슬의 기초에서 주요 1차 생산국을 형성하고 전 세계 산소의 절반을 생산한다.해양 1차 생산자들은 존재하는 데 필요한 거의 모든 산소와 식용 해양동물을 만들어냄으로써 거의 모든 해양동물의 생명을 지탱한다.일부 해양 일차 생산자들은 환경을 바꾸고 다른 해양 생물들에게 서식지를 제공하는 생태계 엔지니어이기도 하다.
해양에서의 1차 생산은 육지에서의 1차 생산과 대조될 수 있다.전 세계적으로 바다와 육지는 각각 거의 같은 양의 1차 생산을 하지만, 해양에서 1차 생산은 주로 시아노박테리아와 조류에서 나오는 반면, 육지에서는 주로 혈관 식물에서 나온다.
해양 조류는 일반적으로 해조류라고 불리는 더 크고, 더 눈에 보이고 복잡한 다세포 대식조류뿐만 아니라, 시아노박테리아와 함께 해양 식물 플랑크톤을 형성하는, 주로 보이지 않는, 종종 단세포 미세조류를 포함합니다.해초는 해안 지역을 따라 발견되며 대륙붕 바닥에 서식하며 조간대에서 밀려옵니다.일부 해조류는 외양의 햇빛이 비치는 지표수(해역)에서 플랑크톤과 함께 떠돌기도 한다.실루리아기에 몇몇 식물성 플랑크톤은 적색, 갈색, 녹조로 진화했다.그리고 나서 이 조류는 육지에 침입하여 오늘날 우리가 알고 있는 육지식물로 진화하기 시작했다.백악기 후반에 이 육지 식물들 중 일부는 맹그로브와 해초로 바다로 돌아왔다.이것들은 조간 지역의 해안과 강 하구의 기수에서 발견된다.게다가, 해초와 같은 일부 해초는 대륙붕의 부드러운 바닥과 단단한 바닥 모두에서 최대 50미터 깊이에서 발견될 수 있다.
| 에 대한 일련의 개요의 일부 |
| 해양생물 |
|---|
 |
해상 1차 생산자
클릭하여 애니메이션을 작성하다
• 빨간색 = 규조류(실리카가 필요한 큰 식물 플랑크톤)
• 노란색 = 편모충류 (기타 큰 식물 플랑크톤)
• 녹색 = 프로클로로코커스(질산염을 사용할 수 없는 작은 식물성 플랑크톤)
• 시안 = Synechoccus (기타 작은 식물성 플랑크톤)
불투명도는 탄소 바이오매스의 농도를 나타낸다.특히,[2][3] 소용돌이와 필라멘트의 역할은 바다에서 높은 생물 다양성을 유지하는 데 있어 중요한 것으로 보인다.
| 의 시리즈의 일부 |
| 탄소 순환 |
|---|
 |
1차 생산자는 다른 유기체를 먹는 대신 그들만의 음식을 만드는 자가영양 생물체이다.이것은 1차 생산자들이 다른 유기체를 먹는 이단영양 유기체의 먹이사슬의 출발점이 된다는 것을 의미한다.일부 해양 1차 생산자는 전문화된 박테리아와 화학영양인 고세균으로, 열수 분출구 및 냉침출구 주변에 모여 화학합성을 이용하여 그들만의 음식을 만든다.하지만, 대부분의 해양 1차 생산은 물에 용해된 이산화탄소에 광합성을 사용하는 유기체로부터 온다.이 과정은 물과 이산화탄소를[4]: 186–187 화학 에너지원으로 사용할 [4]: 1242 수 있는 당과 세포의 구조적 구성요소에 사용되는 유기 분자로 바꾸기 위해 햇빛의 에너지를 사용합니다.해양 일차 생산자들은 다른 유기체들에게 그들이 존재하기 위해 필요한 화학 에너지를 제공하는 대부분의 산소와 음식을 생산함으로써 거의 모든 해양 동물들의 생명을 지탱하기 때문에 중요하다.
주요 해양 일차 생산지는 시아노박테리아, 조류, 해양 식물이다.광합성의 부산물로 방출되는 산소는 거의 모든 생물들이 세포호흡을 하기 위해 필요하다.또한 1차 생산자는 전 세계 탄소 및 물 순환에 영향을 미칩니다.그들은 해안 지역을 안정시키고 해양 동물들에게 서식지를 제공할 수 있다.조류, 곰팡이, 식물에 대한 국제 명명법에서는 현재 [5]이 용어를 동등하게 받아들이고 있지만, 1차 생산자에 대해 논의할 때 문 대신 분할이라는 용어를 전통적으로 사용해 왔다.
육지, 바다에서 일어나는 패턴의 반대로, 거의 모든 광합성은 조류와 시아노박테리아에 의해 이루어지며, 혈관 식물과 다른 그룹에 의해 기여되는 작은 부분들이 있다.해조류는 단일 부유 세포에서 부착된 해초까지 다양한 범위의 유기체를 포함합니다.그들은 다양한 그룹의 광자영양생물을 포함한다.유박테리아는 해양생태계와 육지생태계 모두에서 중요한 광합성물질이며, 일부 고세균은 광영양성이지만, 산소진화 [6]광합성을 이용하는 것은 알려져 있지 않다.많은 진핵생물들이 녹조, 갈조류, 홍조류, 그리고 다양한 단세포 그룹을 포함한 해양에서의 1차 생산에 중요한 기여를 하고 있다.혈관식물은 또한 해초와 같은 그룹에 의해 바다에서 나타난다.
육지 생태계와 달리, 바다에서의 1차 생산의 대부분은 식물성 플랑크톤이라고 불리는 자유 생물에 의해 이루어진다.세계 산소의 절반이 식물성 플랑크톤에 [7][8]의해 생산되는 것으로 추정되어 왔다.해초와 대조류(해초)와 같은 더 큰 자기영양동물은 일반적으로 연안 지역과 인접한 얕은 물에 한정되어 있으며, 그곳에서 기초 기질에 부착될 수 있지만 여전히 광영역 내에 있습니다.Sargassum과 같은 예외는 있지만, 자유 부유 생산의 대부분은 미세한 유기체 안에서 일어납니다.
해양의 1차 생산을 제한하는 요소들 또한 육지에서의 것과 매우 다르다.물의 가용성은 분명히 문제가 되지 않는다.마찬가지로, 온도는 대사율에 영향을 미치지만(Q 참조10), 바닷물의 열 용량이 온도를 완충하고 해빙의 형성은 낮은 온도에서 단열하기 때문에 육지보다 바다에서 덜 넓게 분포한다.그러나 광합성을 위한 에너지원인 빛의 이용과 새로운 성장을 위한 구성 요소인 미네랄 영양소의 이용은 [9]해양에서의 1차 생산을 조절하는 데 중요한 역할을 한다.이용 가능한 지구 시스템 모델은 해양 생물 화학 변화가 방출 [10]시나리오에 따라 현재 값의 3%와 10% 사이의 해양 NPP 감소를 촉발할 수 있음을 시사한다.
2020년 연구진은 북극해에서 지난 20년간 1차 생산량을 측정한 결과 식물성 플랑크톤 농도가 높아 60% 가까이 증가했다고 보고했다.그들은 새로운 영양소들이 다른 바다에서 유입되고 있다는 가설을 세우고 이것은 북극해가 [11][12]미래에 더 높은 영양 수준의 생산과 추가적인 탄소 고정을 지원할 수 있다는 것을 의미한다.
시아노박테리아속

시아노박테리아는 세균의 문(분열)으로 단세포에서 필라멘트형까지 다양하며, 무기 탄소를 유기 탄소 화합물에 고정시키는 군체 종을 포함한다.그들은 지구의 거의 모든 곳에서 발견됩니다: 습한 토양, 담수 환경, 해양 [19]환경, 그리고 심지어 남극 바위에서도 발견됩니다.특히 일부 종은 바다에 떠다니는 세포로 발생하며, 이는 최초의 식물 플랑크톤 중 하나였다.이 박테리아들은 바다에 없을 때 대기 중의 질소를 처리할 수 있다는 점에서 조류와 같은 기능을 한다.

광합성을 사용한 최초의 1차 생산자는 약 23억 년 [20][21]전 해양 시아노박테리아였다.광합성의 부산물로서 시아노박테리아에 의한 분자 산소 방출은 지구 환경의 세계적인 변화를 야기했다.당시 산소는 지구상의 대부분의 생명체에게 독성이었기 때문에, 이것은 산소 불내성 유기체의 거의 멸종으로 이어졌고, 이는 주요 동식물의 [22]진화를 재연하는 극적인 변화로 이어졌다.
.jpg)
1986년에 발견된 이 작은 해양 시아노박테륨 프로클로로코쿠스는 오늘날 해양 먹이사슬의 기저부를 형성하고 있으며[23] 광합성의 절반 이상을 차지하고 있으며 지구 대기 [24]중 산소의 약 20%를 차지하고 있다.그것은 아마도 지구상에서 가장 풍부한 속일 것이다: 1밀리리터의 표면 바닷물은 10만 개 이상의 [25]세포를 포함할 수 있다.
원래 생물학자들은 시아노박테리아가 조류라고 생각했고 그것을 "청록조류"라고 불렀다.더 최근의 견해는 시아노박테리아가 박테리아이기 때문에 조류와 같은 왕국에 있지도 않다는 것이다.대부분의 권위자들은 모든 원핵생물, 즉 시아노박테리아를 [26][27]조류의 정의에서 제외한다.

생물학적 색소
생물학적 색소는 식물이나 동물 세포에 있는 착색 물질이다.모든 생물학적 색소는 다른 파장을 반사하면서 특정 파장의 [28][29]빛을 선택적으로 흡수한다.식물에서 색소의 주요 기능은 광합성인데, 이것은 녹색 색소 엽록소와 가능한 한 많은 빛 에너지를 흡수하는 여러 가지 색소를 사용합니다.엽록소는 식물의 주요 색소이다; 엽록소는 녹색을 반사하면서 노란색과 파란색 파장의 빛을 흡수하는 클로로린이다.식물에게 초록색을 주는 것은 엽록소의 존재와 상대적 풍부함이다.녹조류와 식물들은 이 색소의 두 가지 형태를 가지고 있다: 엽록소 a와 엽록소 b.켈프, 규조류, 그리고 다른 광합성 헤테로콘트들은 b 대신에 클로로필 c를 포함하고 있는 반면, 홍조는 클로로필 a만을 가지고 있다.모든 엽록소는 식물이 광합성을 촉진하기 위해 빛을 차단하는 데 사용하는 주요 수단 역할을 한다.
엽록체
.jpg)

엽록체는 광합성을 하는 세포소기관으로,[31] 광합성 색소 엽록소가 햇빛으로부터 에너지를 포착하고, 그것을 변환하고, 식물과 조류 세포의 물에서 산소를 방출하는 동안 에너지 저장 분자에 저장합니다.그리고 그들은 저장된 에너지를 캘빈 회로라고 알려진 과정에서 이산화탄소로 부터 유기 분자를 만들기 위해 사용한다.
엽록체는 플라스틱으로 알려진 유기체의 한 종류로, 두 개의 막과 고농도의 엽록소를 특징으로 합니다.이들은 매우 역동적입니다. 식물 세포 내에서 순환 및 이동하며 때로는 두 동강 나 번식하기도 합니다.그들의 행동은 밝은 색과 강도와 같은 환경적 요인에 의해 강하게 영향을 받는다.미토콘드리아와 같은 엽록체들은 그들의 조상으로부터 물려받은 것으로 생각되는 그들 자신의 DNA를 포함하고 있는데, 이것은 초기 진핵세포에 의해 [32]삼켜진 광합성 시아노박테륨이다.엽록체는 식물 세포에 의해 만들어질 수 없으며 세포 분열 동안 각 딸 세포에 의해 유전되어야 한다.
대부분의 엽록체는 아마도 시아노박테륨이 진핵생물에 의해 삼켜진 단일 내생생물학적 사건으로 거슬러 올라갈 수 있다.그럼에도 불구하고 엽록체는 매우 광범위한 유기체 집합에서 발견될 수 있으며, 일부는 서로 직접적으로 관련이 없다. 이는 많은 2차적, 심지어 3차적 내분비학적 사건의 결과이다.

미생물 로돕신

광영양대사는 세 가지 에너지 변환 색소 중 하나인 엽록소, 박테리오클로로필, 그리고 망막에 의존한다.망막은 로돕신에서 발견되는 발색단이다.빛 에너지를 변환하는 데 있어서 엽록소의 중요성은 수십 년 동안 쓰여져 왔지만, 망막 색소에 기초한 광영양화는 이제 [35]막 연구되기 시작했습니다.
.jpg)
| 외부 비디오 | |
|---|---|
2000년 에드워드 드롱이 이끄는 미생물학자 팀은 해양 탄소와 에너지 순환에 대한 이해에서 중요한 발견을 했다.그들은 이전에는 박테리아에서 볼 수 없었던 로돕신 단백질 생성에 책임이 있는 여러[37][38] 종의 박테리아에서 유전자를 발견했다.세포막에서 발견되는 이러한 단백질은 햇빛이 로돕신 분자의 형태 변화로 인해 빛 에너지를 생화학적 에너지로 변환시킬 수 있으며,[39] 이로 인해 내부에서 양성자가 펌핑되어 에너지를 발생시키는 후속 유입을 일으킨다.고기와 비슷한 로돕신은 복잡한 다세포 [40][41][42]유기체에서는 드물지만 박테리아와 고세균뿐만 아니라 다양한 분류군, 원생동물들 사이에서도 발견되었다.
2019년의 연구는 이러한 "햇볕을 쬐는 박테리아"가 이전에 생각했던 것보다 더 널리 퍼져 있고, 바다가 지구 온난화의 영향을 받는 방식을 바꿀 수 있다는 것을 보여준다."이 연구결과는 바다에 있는 거의 모든 햇빛이 녹조의 엽록소에 의해 포착된다는 교과서에서 볼 수 있는 해양생태학의 전통적인 해석에서 벗어난다.대신에, 로돕신을 탑재한 박테리아는 하이브리드 자동차와 같은 기능을 합니다.이는 대부분의 박테리아와 마찬가지로 유기물로 구동되며 영양소가 [43][35]부족할 때는 햇빛으로 구동됩니다."
지구상의 원래 생명체들이 엽록소에 기반을 둔 것이 아니라 망막에 기반을 둔 것으로 추측되는 보라색 지구 가설이라고 불리는 우주생물학적 추측이 있는데, 이것은 지구를 [44][45]녹색이 아닌 보라색으로 보이게 했을 것이다.
해조류

| 시리즈의 일부 |
| 플랑크톤 |
|---|
 |
조류는 광합성 진핵 생물의 광범위하고 다양한 집단을 뜻하는 비공식 용어이며, 이들은 반드시 밀접하게 관련되어 있지 않기 때문에 다계통이다.고등 식물과 달리, 해조류는 뿌리, 줄기, 또는 잎이 없다.
조류군
해조류는 전통적으로 녹조, 홍조, 갈조, 규조, 콕콜리소포, 쌍편모충과 같은 그룹으로 분류되어 왔다.
녹조
녹조는 대부분의 삶을 단세포로 살거나 필라멘트로 이루어져 있는 반면, 다른 것들은 긴 세포사슬로 이루어진 군락을 형성하거나 고도로 분화된 거시적 해조류이다.그들은 약 8,000종의 [47]인정된 종을 포함하는 비공식적인 그룹을 형성한다.
홍조류
현대의 홍조는 대부분 분화된 세포를 가진 다세포이고 많은 주목할 만한 [48][49]해조류를 포함하고 있다.산호조류로서, 그들은 산호초의 생태계에서 중요한 역할을 한다.그들은 약 7,000종의 [48]인정된 종을 포함하는 (논쟁의 여지가 있는) 문군을 형성한다.

단세포 홍조류의 일종인 시아니디오파이스아 군락


갈조류
갈조류는 대부분 다세포이고 다시마를 포함한 많은 해조류를 포함한다.그들은 약 2,000종의 [50]인정된 종을 포함하고 있다.
규조류

모두 합쳐서, 해양의 1차 생산의 약 45%가 [51]규조류에 의해 기여된다.
_Various_diatoms.jpg)
![They are a major algae group generating about 20% of world oxygen production.[52]](/wiki/File:Diatoms_through_the_microscope.jpg)
![Diatoms have glass like cell walls called frustules which are made of silica.[53]](/wiki/File:Diatom_algae_Amphora_sp.jpg)
![Diatoms linked in a colonial chain [54]](/wiki/File:Phytoplankton_in_the_form_of_a_diatom_chain.jpg)
콕콜리소포어

어디에나 있는 에밀리아 헉슬레이

에밀리아 헉슬레이는 잉글랜드 남부에서 꽃을 피운다.


콕콜리소포는 거의 전적으로 해양이고 바다의 햇빛 구역에서 다량으로 발견됩니다.그들은 중요한 미세 화석인 콕콜리스라고 불리는 기능이 불확실한 탄산칼슘 판을 가지고 있습니다.콕콜리소포어는 해양 산성도가 높아짐에 따라 [55]탄소 흡수원으로서 더욱 중요해질 수 있기 때문에 지구 기후 변화를 연구하는 사람들에게 흥미롭다.가장 풍부한 콕콜리소포어 종인 에밀리아 헉슬레이는 해양 먹이 [56]거미줄에 있는 플랑크톤 베이스의 유비쿼터스 성분입니다.부영양화 관련 콕콜리소포(coccolithophore)의 꽃은 해수면 [57]하부로의 영양소 흐름 감소로 이어지기 때문에 이를 방지하기 위한 관리전략이 강구되고 있다.
쌍편모충류

디노플라겔라테스

![Karenia brevis produces red tides highly toxic to humans[58]](/wiki/File:Karenia_brevis.jpg)
_by_Noctiluca_in_Nagasaki.jpg)
혼합영양조류
기타 그룹

디플로네미드는 세계 해양에 풍부할 수 있다

전통적으로 위에서 논의한 조류 집단과 같은 미생물의 계통 발생이 추론되었고 형태학 연구에 기초하여 그 분류법이 확립되었다.그러나 분자 계통학에서의 발전은 종의 DNA와 단백질 [59]서열을 분석함으로써 종의 진화적 관계를 확립할 수 있게 했다.위에서 설명한 조류군을 포함한 많은 분류군은 분자 계통학을 사용하여 재분류되거나 재정의되는 과정에 있다.최근 분자 배열 분석의 발달로 환경 샘플에서 직접 게놈을 회수하고 배양 필요성을 피할 수 있게 되었습니다.예를 들어, 해양 미생물의 풍부함과 다양성에 대한 지식의 급속한 확대로 이어졌다.게놈 분해 메타게노믹스나 단세포게노믹스와 같은 분자 기술이 높은 처리량 기술과 함께 사용되고 있다.
2009년과 2013년 사이에, 타라 오션스 탐험대는 플랑크톤을 채취하고 현대 분자 기술로 그것들을 분석하기 위해 세계 바다를 횡단했다.그들은 이전에는 알려지지 않았던 광합성 [60]조류와 혼합영양조류를 광범위하게 발견했다.그들의 발견들 중에는 디플로네미드가 있었다.이러한 유기체는 일반적으로 무색이고 장방형이며, 일반적으로 길이가 약 20µm이고 편모가 [61]2개 있다.DNA 바코딩의 증거는 디플로네미드가 모든 해양 진핵생물군 [62][63]중에서 가장 풍부하고 가장 풍부한 종족 중 하나일 수 있다는 것을 암시한다.
크기별
조류는 크기에 따라 미세조류 또는 거시조류로 분류될 수 있다.
미세조류
미세조류는 육안으로는 볼 수 없는 미세한 종류의 조류이다.그들은 대부분 개인 또는 사슬이나 그룹으로 존재하는 단세포 종이지만, 일부는 다세포 종이다.미세조류는 해양 식물성 플랑크톤뿐만 아니라 해양 원생 동물들의 중요한 구성요소이다.그들은 매우 다양합니다.20만~80만 종의 생물이 있는 것으로 추정되고 있으며, 그 중 약 5만 종의 생물이 [64]기술되어 있다.종에 따라, 그들의 크기는 몇 마이크로미터에서 몇 백 마이크로미터까지 다양합니다.그것들은 점성력에 의해 지배되는 환경에 특별히 적응되어 있다.
- 미세조류


_(cropped).jpg)
대조류

대조류는 일반적으로 해조류라고 불리는 더 크고, 다세포이고, 더 눈에 띄는 종류의 조류이다.미역은 보통 얕은 해안에서 자라는데, 바닷바닥에 닻을 내리고 있습니다.표류한 해초는 해변으로 떠내려갈 수 있다.다시마는 세계 해안선의 [65]약 25%를 덮고 있는 커다란 수중 숲을 형성하는 큰 갈색 해조류입니다.그들은 지구에서 [66]가장 생산적이고 역동적인 생태계 중 하나이다.일부 해조류는 플랑크톤(자유 부유성)으로 부유 [67]: 246–255 표류를 형성한다.미세조류처럼, 대식물은 진정한 식물이 아니기 때문에 기술적으로 해양 원생동물이다.


- 대조류

자이언트 다시마는 진짜 식물이 아니기 때문에 기술적으로 원생동물이지만 다세포이고 50m까지 자랄 수 있다.

살가슴 해초는 공기 방광을 가진 갈색 조류이다.

표류하는 해초 사이에서 살기 위해 위장된 물고기.



![This unicellular bubble algae lives in tidal zones. It can have a 4 cm diameter.[68]](/wiki/File:Ventricaria_ventricosa.JPG)
육지식물의 진화

오른쪽 그림은 줄생식물에 [69]의한 토지 정복에 대한 진화 시나리오를 보여준다.스트렙토식물 조류는 모든 녹조를 포함하며, 육안 육지 식물군이 진화한 유일한 광합성 진핵생물입니다.즉, 진화의 과정 내내, 다양한 다른 계통에서 온 조류가 땅을 식민지로 만들었습니다(노란색 선). 그러나 또한 연쇄식물 조류는 습한 상태에서 건조한 상태로 지속적으로 독립적으로 전환되었습니다(빨간색과 노란색의 융합).역사를 통틀어 수많은 혈통이 멸종되었다(X라벨).다양한 분류학적 연관성을 가진 육생 조류들은 암석 표면에 살고 생물학적 토양 지각들을 형성한다.그러나 측문성 줄생식물 조류의 다양성으로부터 후손들이 마침내 전지구적 규모로 땅을 정복한 유기체, 즉 뿌리꼴 구조를 형성하고 때때로 건조증을 경험하는 가지 모양의 필라멘트 또는 심지어 실질질의 유기체가 나타났다.이 "가상의 수생 육생 조류"에서 지그네마토피체아류와 태생식물(육지식물)의 혈통이 생겨났다.[69]초기 단계에서, 배아식물로 이어지는 궤적은 현재 멸종된 최초의 [71]육지식물로 표현되었다.
최초의 육지 식물은 아마도 유익한 기질 미생물군과 상호작용하여 기질로부터 영양분을 얻는 데 도움을 주었다.또한, 초기 육상 식물은 지상 스트레스 요인(자외선과 광합성 활성 방사선 강도, 가뭄, 급격한 온도 변화 등)을 성공적으로 극복해야 했다.그들이 성공한 이유는 그들이 올바른 특성 세트를 가지고 있었기 때문입니다. 즉, 그들의 수생-지상 조류 조상에서 선택된 적응의 혼합, 박멸, 그리고 우연한 일련의 유전자와 [69]경로의 공동 선택 가능성입니다.진화 과정에서 초기 육지 식물 집단의 일부 구성원은 육상 환경(예를 들어 어떤 형태의 물 전도, 기공과 같은 구조, 배아 등)에 적응 가능한 특성을 얻었다. 결국 "육지 식물의 가상의 마지막 공통 조상"이 나타났다.이 조상으로부터 현존하는 이끼식물들과 기관식물들이 진화했다.육지식물의 가상의 마지막 공통 조상의 정확한 특성 레퍼토리는 불확실하지만, 그것은 분명히 혈관 식물과 비혈관 식물의 특성을 수반할 것이다.또한 확실한 것은 육지 식물의 마지막 공통 조상이 조류 [69]조상의 특징을 가지고 있었다는 것이다.
해양 식물

실루리아기에 몇몇 식물성 플랑크톤은 적색, 갈색, 녹조로 진화했다.녹조가 육지에 침입하여 오늘날 우리가 알고 있는 육지식물로 진화하기 시작했다.나중에 백악기에 이 육지 식물들 중 일부는 맹그로브와 [72]해초로 바다로 돌아왔다.
맹그로브나 탯줄풀이나 해변의 풀이 자랄 수 있는 강어귀의 염수에서 식물 생물이 번성할 수 있다.개화식물은 해초지 형태의 [73]모래 여울에서 자라며, 맹그로브는 열대 및 아열대[74] 지역의 해안가에 늘어서 있으며, 염분에 강한 식물은 정기적으로 침수되는 [75]염습지에서 잘 자란다.이 모든 서식지는 많은 양의 탄소를 격리시킬 수 있고 점점 더 크고 작은 [76]동물들의 생물다양한 범위를 지탱할 수 있습니다.해양식물은 조간대나 얕은 물에서 볼 수 있는데, 예를 들어 장어풀이나 거북풀과 같은 해초와 살라시아가 그것이다.이 식물들은 해양 환경의 높은 염도에 적응했다.
빛은 꼭대기 200미터(660피트)까지만 투과할 수 있기 때문에 식물이 [77]자랄 수 있는 바다의 유일한 부분입니다.표면층은 종종 생물학적으로 활성 질소 화합물이 부족하다.해양 질소 사이클은 질소 고정, 질소 동화, 질화,[78] 아나목스 및 탈질소를 포함하는 복잡한 미생물 변형으로 구성됩니다.이러한 과정 중 일부는 깊은 물에서 일어나 차가운 물이 솟아오르는 곳과 육지 영양소가 있는 강 하구 부근에서 식물이 더 많이 자랄 수 있습니다.이것은 플랑크톤이 풍부하고 따라서 어류가 풍부한 가장 생산적인 지역이 주로 [79]: 160–163 해안 지역이라는 것을 의미합니다.
맹그로브
맹그로브는 대형 어류와 무척추동물의 유충과 어린 형태를 감추고 먹이를 찾는 장소로서 해양 생물에게 중요한 요람 서식지를 제공합니다.위성 자료에 근거하여, 맹그로브 숲의 총 세계 면적은 2010년에 134,257 [80][81]평방 킬로미터로 추정되었다.



- Spalding, M. (2010) 루트리지 맹그로브 세계 지도책. ISBN978184976608.doi:10.4324/978184976608.
해초류
맹그로브 숲처럼, 해초는 더 큰 물고기와 무척추동물의 유충과 어린 형태를 위한 중요한 요람 서식지를 제공합니다.해초 초원의 총 세계 면적은 맹그로브 숲보다 더 어렵지만, 2003년에는 177,000 평방 킬로미터(68,000 평방 mi)[82]로 보수적으로 추정되었다.

![Sea dragons camouflaged to look like floating seaweed live in kelp forests and seagrass meadows[83]](/wiki/File:Leafy_Sea_Dragon_SA.jpg)
| 외부 비디오 | |
|---|---|
스토이치메트리
지표면 해양에서의 1차 생산의 화학량 측정(화학반응물질과 생성물의 측정)은 지구 해양의 원소 순환에 중요한 역할을 한다.C:N:P 비율로 표현되는 수출 유기물의 탄소(C), 질소(N), 인(P) 요소 간의 비율은 제한 [84]영양소의 가용성과 관련하여 심해에서 격리된 대기 탄소의 양을 결정하는 데 도움이 됩니다.지질학적 타임스케일에서 N:P 비율은 인산염에 대한 질산염의 상대적 가용성을 반영하며, 둘 다 대기 중 질소 고정 및/또는 하천 공급을 통해 대륙에서 외부로 공급되며 탈질 및 [85][86][87][88]매립으로 손실된다.짧은 시간 척도에서 수출된 벌크 입자 유기물의 평균 화학량계는 식물성 플랑크톤의 [89][90][91]원소 화학량계를 반영하며, 동물성 플랑크톤과 이종 영양균에 의한 유기물의 생물학적 다양성과 2차 처리의 영향을 추가로 받는다.전지구적 변화에 직면하여 미래의 [84]기후변화를 정확하게 예측하기 위해서는 C:N:P 비율의 변동을 초래하는 메커니즘을 이해하고 정량화하는 것이 중요하다.
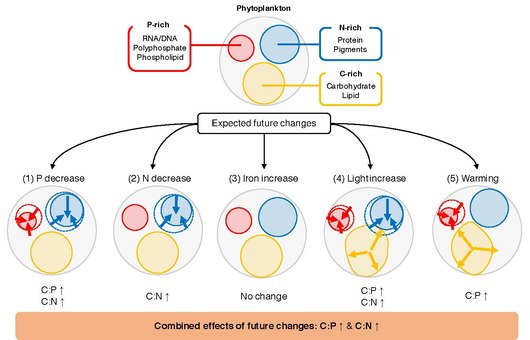
주요 환경 운동가들에게
주요 미해결 질문은 개별 식물성 플랑크톤의 C:N:P를 결정하는 것이다.식물성 플랑크톤은 무기 영양소, 빛, 그리고 온도가 공간적으로 그리고 [84]시간적으로 변화하는 바다의 밝은 윗층에서 자랍니다.실험실 연구에 따르면 이러한 변동은 세포 수준에서 반응을 유발하며, 세포는 주변 [92]환경에 최적으로 적응하기 위해 자원 할당을 수정한다.예를 들어 식물성 플랑크톤은 P농축생합성장치, N농축광장치 및 C농축에너지 [93]비축량 간의 자원 할당을 변경할 수 있다.전형적인 미래 온난화 시나리오에서 지구 바다는 영양소 가용성, 온도 및 방사선 [94]강도의 변화를 겪을 것으로 예상된다.이러한 변화는 식물성 플랑크톤의 [95][96]생리에 깊은 영향을 미칠 수 있으며, 관찰 결과 경쟁적인 식물성 플랑크톤 종은 10대 시간 [97]척도의 온도, 방사선 강도 및 영양소의 변화에 적응하고 적응할 수 있는 것으로 나타났습니다.식물성 플랑크톤의 C:N:P 비율과 환경 드라이버 사이의 관계를 연구하는 수많은 실험실 및 현장 실험이 수행되었다.그러나 이러한 연구를 합성하고 식물성 플랑크톤 C:N의 반응을 일반화하기는 어렵다.환경 [84]동인의 변화에 대한 P.개별 연구는 단순 t 테스트에서 더 복잡한 혼합 모델에 이르기까지 요소 비율에 대한 환경 동인의 영향을 특징짓기 위해 서로 다른 통계 분석 세트를 사용한다. 이는 상호 연구 비교를 어렵게 한다.또한 환경유발 특성변화는 가소성(승화), 적응성 및 [98][99]생활역사의 조합에 의해 이루어지기 때문에 식물성플랑크톤의 화학량학적 반응은 밀접한 관련이 있는 [84]종들 사이에서도 가변적일 수 있다.
메타 분석/체계적 검토는 독립적인 연구에서 얻은 연구 결과를 종합하고 통합하고 일반적인 [100]추세를 파악하기 위한 강력한 통계 프레임워크이다.2002년 [101]Geider와 La Roche에 의한 정합성 [102]및 2010년 Persson 등의 보다 최근의 연구에서 C:P와 N:P는 영양소 보충 세포와 영양소 제한 세포 사이에서 최대 20배까지 차이가 날 수 있는 것으로 나타났다.이러한 연구들은 또한 영양소 제한으로 인해 C:N 비율이 약간 가소성이 될 수 있다는 것을 보여주었다.2013년 Hillebrand 등의 메타 분석 연구는 원소 화학량 측정에서 성장률의 중요성을 강조했으며 [103]C:P와 N:P 비율이 증가율에 따라 감소하는 것으로 나타났다.2015년 Yvon-Durocher 등은 C:N:[104]P 조절에 있어 온도의 역할을 조사하였다.이들의 데이터 세트는 1996년 이전에 수행된 연구로 제한되었지만 C:P와 온도 상승 사이에 통계적으로 유의한 관계가 있음을 보여주었다.MacIntyre 등(2002)[105] 및 Thrane et al. (2016)[106]는 방사조도가 최적의 세포 C:N 및 N:P 비율을 제어하는데 중요한 역할을 한다는 것을 보여주었다.가장 최근에, Moreno와 Martiny(2018)는 환경 조건이 생리학적 [107][84]관점에서 세포 화학량 측정을 어떻게 규제하는지에 대한 포괄적인 요약을 제공했다.
해양 식물성 플랑크톤의 원소 화학 측정법은 영양 순환, 2차 생산, 탄소 수출에 대한 영향을 통해 전세계 생물 지구 화학적 순환에 중요한 역할을 한다.식물성 플랑크톤의 원소 구성에 대한 다양한 환경 동인의 영향을 평가하기 위해 수년간 광범위한 실험실 실험이 수행되었지만, 프로세스에 대한 종합적인 정량적 평가는 여전히 부족하다.여기서 해양식물플랑크톤의 P:C 및 N:C 비율의 반응은 104개 저널 기사의 366개 실험 데이터에 대한 메타분석에 의해 5대 드라이버(무기인, 무기질소, 무기철, 무기철, 방사강도, 온도)에 대해 합성되었다.이러한 결과는 영양소 이용률의 증가가 P:C 및 N:C 비율의 변화와 긍정적인 관련이 있는 모든 연구에서 이러한 비율의 반응이 일치함을 보여준다.그 결과는 진핵생물인 식물성 플랑크톤이 아마도 더 큰 세포 크기와 유전자 발현 패턴을 빠르게 조절하는 능력 때문에 원핵생물들에 비해 마크롱 영양소의 변화에 더 민감하다는 것을 보여준다.방사선 강도의 영향은 모든 연구에서 유의하고 일정했으며, 방사선 강도의 증가는 P:C와 N:C를 모두 감소시켰다.온난화와 함께 P:C 비율은 크게 감소하였으나, 재배 생육 모드와 수확 시 생육 단계에 따라 온도 변화에 대한 반응이 엇갈렸다.아열대 회전의 다른 해양학적 조건(예: 낮은 마크롱 영양소 가용성)과 함께, 높아진 온도는 왜 P:C가 아열대 바다에서 지속적으로 낮은지를 설명할 수 있다.철 첨가는 P:C 또는 N:C [84]중 어느 쪽도 체계적으로 변경되지 않았습니다.
진화 연표
.png)

「 」를 참조해 주세요.
레퍼런스
- ^ 클로로필 NASA 지구 관측소입니다.2019년 11월 30일 접속.
- ^ 지구 대양의 식물성 플랑크톤 군집 모델링NASA Hyperwall, 2015년 9월 30일
 이 문서에는 퍼블릭 도메인에 있는 이 소스로부터의 텍스트가 포함되어 있습니다..
이 문서에는 퍼블릭 도메인에 있는 이 소스로부터의 텍스트가 포함되어 있습니다.. - ^ 다윈 프로젝트 매사추세츠 공과대학.
- ^ a b Campbell, Neil A.; Reece, Jane B.; Urry, Lisa Andrea; Cain, Michael L.; Wasserman, Steven Alexander; Minorsky, Peter V.; Jackson, Robert Bradley (2008). Biology (8 ed.). San Francisco: Pearson – Benjamin Cummings. ISBN 978-0-321-54325-7.
- ^ McNeill, J.; et al., eds. (2012). International Code of Nomenclature for algae, fungi, and plants (Melbourne Code), Adopted by the Eighteenth International Botanical Congress Melbourne, Australia, July 2011 (electronic ed.). International Association for Plant Taxonomy. Retrieved 14 May 2017.
- ^ Schäfer G, Engelhard M, Müller V (1 September 1999). "Bioenergetics of the Archaea". Microbiol. Mol. Biol. Rev. 63 (3): 570–620. doi:10.1128/MMBR.63.3.570-620.1999. PMC 103747. PMID 10477309.
- ^ Roach, John (7 June 2004). "Source of Half Earth's Oxygen Gets Little Credit". National Geographic News. Retrieved 4 April 2016.
- ^ Lin, I.; Liu, W. Timothy; Wu, Chun-Chieh; Wong, George T. F.; Hu, Chuanmin; Chen, Zhiqiang; Wen-Der, Liang; Yang, Yih; Liu, Kon-Kee (2003). "New evidence for enhanced ocean primary production triggered by tropical cyclone". Geophysical Research Letters. 30 (13): 1718. Bibcode:2003GeoRL..30.1718L. doi:10.1029/2003GL017141. S2CID 10267488.
- ^ Sigman, D.M.; Hain, M.P. (2012). "The Biological Productivity of the Ocean" (PDF). Nature Education Knowledge. 3 (6): 1–16. Retrieved 1 June 2015.
The deep chlorophyll maximum (DCM) occurs at the contact where there is adequate light for photosynthesis and yet significant nutrient supply from below.
- ^ Mora, C.; et al. (2013). "Biotic and Human Vulnerability to Projected Changes in Ocean Biogeochemistry over the 21st Century". PLOS Biology. 11 (10): e1001682. doi:10.1371/journal.pbio.1001682. PMC 3797030. PMID 24143135.
- ^ "A 'regime shift' is happening in the Arctic Ocean, scientists say". phys.org. Retrieved 16 August 2020.
- ^ Lewis, K. M.; Dijken, G. L. van; Arrigo, K. R. (10 July 2020). "Changes in phytoplankton concentration now drive increased Arctic Ocean primary production". Science. 369 (6500): 198–202. doi:10.1126/science.aay8380. ISSN 0036-8075. PMID 32647002. S2CID 220433818. Retrieved 16 August 2020.
- ^ Olson, J. M. and Blankenship, R. E. (2005) "광합성의 진화에 대해 생각한다."인: 광합성에 관한 발견, 1073-1086쪽, 스프링거.ISBN 9781402033247.doi:10.1007/1-4020-3324-9_95.
- ^ Blankenship, R. E., Sadekar, S. 및 Raymond, J. (2007) "비산소 광합성에서 산소 광합성으로의 진화적 전환"인: 수생 광자동영양생물 진화, eds P. G. Falkowski 및 A.N. Knoll, 뉴욕: 학술 프레스, 21-35페이지. doi:10.1016/B978-012370518-1/50004-7.
- ^ Hohmann-Marriott, M.F.와 Blankenship, R.E. (2011) "광합성의 진화"식물생물학 연간 리뷰, 62:515-548.doi:10.1146/anurev-arplant-042110-103811.
- ^ Kim, E., Harrison, J.W., Sudek, S., Jones, M.D., Wilcox, H.M., Richards, T.A., Worden, A.Z. 및 J.M.(2011) "진핵 플라스티드에서 새롭게 식별되고 다양한 플라스틱을 가진"미국 국립과학원회보, 108(4): 1496–1500. doi:10.1073/pnas.10337108.
- ^ Garcia-Mendoza, E. 및 Ocampo-Alvarez, H. (2011년) "갈색 조류 대식세포의 사진 보호: 진화적 의미"광화학 및 광생물학 B: 생물학, 104(1-2): 377-385. doi:10.1016/j.photobiol.2011.04.004.
- ^ Shevela, D. (2011) "시아노박테리아를 이용한 모험: 개인적인 관점"식물과학 분야 프런티어, 2:28.doi:10.3389/fpls.2011.00028.
- ^ Walsh PJ, Smith S, Fleming L, Solo-Gabriele H, Gerwick WH, eds. (2 September 2011). "Cyanobacteria and cyanobacterial toxins". Oceans and Human Health: Risks and Remedies from the Seas. Academic Press. pp. 271–296. ISBN 978-0-08-087782-2.
- ^ "The Rise of Oxygen - Astrobiology Magazine". Astrobiology Magazine. 30 July 2003. Retrieved 6 April 2016.
- ^ Flannery, D. T.; R.M. Walter (2012). "Archean tufted microbial mats and the Great Oxidation Event: new insights into an ancient problem". Australian Journal of Earth Sciences. 59 (1): 1–11. Bibcode:2012AuJES..59....1F. doi:10.1080/08120099.2011.607849. S2CID 53618061.
- ^ Rothschild, Lynn (September 2003). "Understand the evolutionary mechanisms and environmental limits of life". NASA. Archived from the original on 29 March 2012. Retrieved 13 July 2009.
- ^ Nadis S (December 2003). "The cells that rule the seas" (PDF). Scientific American. 289 (6): 52–3. Bibcode:2003SciAm.289f..52N. doi:10.1038/scientificamerican1203-52. PMID 14631732. Archived from the original (PDF) on 19 April 2014. Retrieved 11 July 2019.
- ^ "The Most Important Microbe You've Never Heard Of". npr.org.
- ^ Flombaum, P.; Gallegos, J. L.; Gordillo, R. A.; Rincon, J.; Zabala, L. L.; Jiao, N.; Karl, D. M.; Li, W. K. W.; Lomas, M. W.; Veneziano, D.; Vera, C. S.; Vrugt, J. A.; Martiny, A. C. (2013). "Present and future global distributions of the marine Cyanobacteria Prochlorococcus and Synechococcus". Proceedings of the National Academy of Sciences. 110 (24): 9824–9829. Bibcode:2013PNAS..110.9824F. doi:10.1073/pnas.1307701110. PMC 3683724. PMID 23703908.
- ^ Nabors, Murray W. (2004). Introduction to Botany. San Francisco, CA: Pearson Education, Inc. ISBN 978-0-8053-4416-5.
- ^ Allaby, M., ed. (1992). "Algae". The Concise Dictionary of Botany. Oxford: Oxford University Press.
- ^ Grotewold, E. (2006). "The Genetics and Biochemistry of Floral Pigments". Annual Review of Plant Biology. 57: 761–780. doi:10.1146/annurev.arplant.57.032905.105248. PMID 16669781.
- ^ Lee, DW (2007) 자연의 팔레트 - 식물 색채의 과학.시카고 대학교 출판부
- ^ 생물학 개념: 진핵생물 기원OpenStax CNX.2020년 7월 16일 회수.
 수정된 텍스트는 Creative Commons Attribution 4.0 International License에 따라 제공되는 이 원본에서 복사되었습니다.
수정된 텍스트는 Creative Commons Attribution 4.0 International License에 따라 제공되는 이 원본에서 복사되었습니다. - ^ "chloroplast". Online Etymology Dictionary.
- ^ Basic Biology (18 March 2016). "Bacteria".
- ^ Patrick J. Keeling (2004). "Diversity and evolutionary history of plastids and their hosts". American Journal of Botany. 91 (10): 1481–1493. doi:10.3732/ajb.91.10.1481. PMID 21652304.
- ^ DeLong, E.F.; Beja, O. (2010). "The light-driven proton pump proteorhodopsin enhances bacterial survival during tough times". PLOS Biology. 8 (4): e1000359. doi:10.1371/journal.pbio.1000359. PMC 2860490. PMID 20436957. e1000359.
- ^ a b Gómez-Consarnau, L.; Raven, J.A.; Levine, N.M.; Cutter, L.S.; Wang, D.; Seegers, B.; Arístegui, J.; Fuhrman, J.A.; Gasol, J.M.; Sañudo-Wilhelmy, S.A. (2019). "Microbial rhodopsins are major contributors to the solar energy captured in the sea". Science Advances. 5 (8): eaaw8855. Bibcode:2019SciA....5.8855G. doi:10.1126/sciadv.aaw8855. PMC 6685716. PMID 31457093.
- ^ Oren, Aharon (2002). "Molecular ecology of extremely halophilic Archaea and Bacteria". FEMS Microbiology Ecology. 39 (1): 1–7. doi:10.1111/j.1574-6941.2002.tb00900.x. ISSN 0168-6496. PMID 19709178.
- ^ Béja, O.; Aravind, L.; Koonin, E.V.; Suzuki, M.T.; Hadd, A.; Nguyen, L.P.; Jovanovich, S.B.; Gates, C.M.; Feldman, R.A.; Spudich, J.L.; Spudich, E.N. (2000). "Bacterial rhodopsin: evidence for a new type of phototrophy in the sea". Science. 289 (5486): 1902–1906. Bibcode:2000Sci...289.1902B. doi:10.1126/science.289.5486.1902. PMID 10988064. S2CID 1461255.
- ^ "Interviews with Fellows: Ed Delong". American Academy of Microbiology. Archived from the original on 7 August 2016. Retrieved 2 July 2016.
- ^ 배터리 달린 박테리아, Popular Science, 2001년 1월, 55페이지.
- ^ Béja, O.; Aravind, L.; Koonin, E.V.; Suzuki, M.T.; Hadd, A.; Nguyen, L.P.; Jovanovich, S.B.; Gates, C.M.; Feldman, R.A.; Spudich, J.L.; Spudich, E.N. (2000). "Bacterial rhodopsin: evidence for a new type of phototrophy in the sea". Science. 289 (5486): 1902–1906. Bibcode:2000Sci...289.1902B. doi:10.1126/science.289.5486.1902. PMID 10988064.
- ^ Boeuf, Dominique; Audic, Stéphane; Brillet-Guéguen, Loraine; Caron, Christophe; Jeanthon, Christian (2015). "MicRhoDE: a curated database for the analysis of microbial rhodopsin diversity and evolution". Database. 2015: bav080. doi:10.1093/database/bav080. ISSN 1758-0463. PMC 4539915. PMID 26286928.
- ^ Yawo, Hiromu; Kandori, Hideki; Koizumi, Amane (5 June 2015). Optogenetics: Light-Sensing Proteins and Their Applications. Springer. pp. 3–4. ISBN 978-4-431-55516-2. Retrieved 30 September 2015.
- ^ 작은 해양 미생물은 2019년 8월 8일 캘리포니아 남부 대학 프레스룸에서 큰 역할을 할 수 있다.
- ^ DasSarma, Shiladitya; Schwieterman, Edward W. (11 October 2018). "Early evolution of purple retinal pigments on Earth and implications for exoplanet biosignatures". International Journal of Astrobiology. 20 (3): 241–250. arXiv:1810.05150. Bibcode:2018arXiv181005150D. doi:10.1017/S1473550418000423. ISSN 1473-5504. S2CID 119341330.
- ^ Sparks, William B.; DasSarma, S.; Reid, I. N. (December 2006). "Evolutionary Competition Between Primitive Photosynthetic Systems: Existence of an early purple Earth?". American Astronomical Society Meeting Abstracts. 38: 901. Bibcode:2006AAS...209.0605S.
- ^ Javed, M.R., Bilal, M.J., Ashraf, M.U.F, Waqar, A., Mehmood, M.A., Saed, M. 및 N. N. N. Nashat.(2019) "바이오 연료 생산을 위한 원료로서의 마이크로 조류: 「현황과 장래의 전망」인: 에너지 연구 개발에 있어서의 공헌도 톱 5, 제3판, 제2장, Avid Science.ISBN 978-93-88170-77-2. 수정된 텍스트는 Creative Commons Attribution 4.0 International License에 따라 제공되는 이 원본에서 복사되었습니다.
- ^ Guiry MD (October 2012). "How many species of algae are there?". Journal of Phycology. 48 (5): 1057–63. doi:10.1111/j.1529-8817.2012.01222.x. PMID 27011267. S2CID 30911529.
- ^ a b Guiry, M.D.; Guiry, G.M. (2016). "Algaebase". www.algaebase.org. Retrieved 20 November 2016.
- ^ D. Thomas (2002). Seaweeds. Life Series. Natural History Museum, London. ISBN 978-0-565-09175-0.
- ^ Hoek, Christiaan; den Hoeck, Hoeck Van; Mann, David; Jahns, H.M. (1995). Algae : an introduction to phycology. Cambridge University Press. p. 166. ISBN 9780521316873. OCLC 443576944.
- ^ Yool, A.; Tyrrell, T. (2003). "Role of diatoms in regulating the ocean's silicon cycle". Global Biogeochemical Cycles. 17 (4): n/a. Bibcode:2003GBioC..17.1103Y. CiteSeerX 10.1.1.394.3912. doi:10.1029/2002GB002018. S2CID 16849373.
- ^ 당신이 숨쉬는 공기?그것을 만든 규조류
- ^ "More on Diatoms". University of California Museum of Paleontology. Archived from the original on 4 October 2012. Retrieved 11 July 2019.
- ^ 이 황혼지대는 어둡고, 물이 많고, 그렇다.또한 2018년 8월 21일, NASA 블로그의 흥미로운 내용도 가득하다.
- ^ Smith, H.E.K.; et al. (2012), "Predominance of heavily calcified coccolithophores at low CaCO3 saturation during winter in the Bay of Biscay", Proceedings of the National Academy of Sciences, 109 (23): 8845–8849, Bibcode:2012PNAS..109.8845S, doi:10.1073/pnas.1117508109, PMC 3384182, PMID 22615387
- ^ "Biogeography and dispersal of micro-organisms: a review emphasizing protists", Acta Protozoologica, 45 (2): 111–136, 2005
- ^ Yunev, O.A.; et al. (2007), "Nutrient and phytoplankton trends on the western Black Sea shelf in response to cultural eutrophication and climate changes", Estuarine, Coastal and Shelf Science, 74 (1–2): 63–67, Bibcode:2007ECSS...74...63Y, doi:10.1016/j.ecss.2007.03.030
- ^ Brand, Larry E.; Campbell, Lisa; Bresnan, Eileen (2012). "Karenia: The biology and ecology of a toxic genus". Harmful Algae. 14: 156–178. doi:10.1016/j.hal.2011.10.020.
- ^ Olsen GJ, Woese CR, Overbeek R (1994). "The winds of (evolutionary) change: breathing new life into microbiology". Journal of Bacteriology. 176 (1): 1–6. doi:10.2172/205047. PMC 205007. PMID 8282683.
- ^ Bork, P., Bowler, C., De Vargas, C., Gorsky, G., Karsenti, E. 및 Winker, P. (2015) "타라 오션은 행성 규모로 플랑크톤을 연구한다." doi:10.1126/science.
- ^ Gawryluk, Ryan M.R.; Del Campo, Javier; Okamoto, Noriko; Strassert, Jürgen F.H.; Lukeš, Julius; Richards, Thomas A.; Worden, Alexandra Z.; Santoro, Alyson E.; Keeling, Patrick J. (2016). "Morphological Identification and Single-Cell Genomics of Marine Diplonemids". Current Biology. 26 (22): 3053–3059. doi:10.1016/j.cub.2016.09.013. PMID 27875688.
- ^ 박토로바, D., 도바코바, E., 페냐-디아즈, P. 및 루크시, J., 2016.단순한 것부터 초복잡한 것까지: 우글레노존 원생들의 미토콘드리아 게놈.F1000 Research, 5. doi:10.12688/f1000 Research.8040.1. 수정된 텍스트는 Creative Commons Attribution 4.0 International License에 따라 제공되는 이 원본에서 복사되었습니다.
- ^ De Vargas, C., Audic, S., Henry, N., Delayle, J., Mahé, F., Logares, R., Lara, E., Berney, C., Le Bescot, N., Probert, I., Carmichael, Mary 및 기타 44. (2015)과학", 348(6237): 1261605.doi: 10.1126/science.1265.
- ^ Starckx, Senne (2012년 10월 31일) 태양 아래 있는 곳 - 조류는 미래의 작물이라고 Geel Flanders Today, Retrieved 2012년 12월 8일
- ^ Wernberg, T., K., K., Filbee-Dexter, K. 및 M.F. Pedersen.(2019) "세계 다시마 숲의 현황과 동향"In: World sea: 환경 평가, 57~78페이지).학술용 프레스: 10.1016/B978-0-12-805052-1.00003-6.
- ^ 1973년 K.H. 만미역: 생산성과 성장 전략.과학 182: 975-981.
- ^ Kindersley, Dorling (2011). Illustrated Encyclopedia of the Ocean. Dorling Kindersley. ISBN 978-1-4053-3308-5.
- ^ Tunnell, John Wesley; Chávez, Ernesto A.; Withers, Kim (2007). Coral reefs of the southern Gulf of Mexico. Texas A&M University Press. p. 91. ISBN 978-1-58544-617-9.
- ^ a b c d e 드프리스:네덜란드의, 얀, Hugo, 소피, Fürst-Jansen, 재닌 MR.(2020년)."Evo-physio:스트레스 반응 및 초기 땅 plants".실험 식물학 신문. 71(11):3254–3269. doi:10.1093/jxb/eraa007.PMC 7289718.PMID 31922568.수정 텍스트는 창조적 공용 귀인 4.0국제 라이센스 하에 가능하다 이 원본에서 복사되었다.
- ^ Morris, Jennifer L.; Puttick, Mark N.; Clark, James W.; Edwards, Dianne; Kenrick, Paul; Pressel, Silvia; Wellman, Charles H.; Yang, Ziheng; Schneider, Harald; Donoghue, Philip C. J. (2018). "The timescale of early land plant evolution". Proceedings of the National Academy of Sciences. 115 (10): E2274–E2283. Bibcode:2018PNAS..115E2274M. doi:10.1073/pnas.1719588115. PMC 5877938. PMID 29463716.
- ^ Delaux, Pierre-Marc; Hetherington, Alexander J.; Coudert, Yoan; Delwiche, Charles; Dunand, Christophe; Gould, Sven; Kenrick, Paul; Li, Fay-Wei; Philippe, Hervé; Rensing, Stefan A.; Rich, Mélanie; Strullu-Derrien, Christine; De Vries, Jan (2019). "Reconstructing trait evolution in plant evo–devo studies". Current Biology. 29 (21): R1110–R1118. doi:10.1016/j.cub.2019.09.044. PMID 31689391. S2CID 207844920.
- ^ Orth, R.J., Caruthers, T.J., Dennison, W.C., Duarte, C.M., Fourqueran, J.W., Heck, K.L., Hughes, A.R., Kendrick, Kenworth, W.바이오사이언스, 56(12): 987~996페이지.doi:10.1641/0006-3568(2006)56[987:AGCFSE2.0.CO;2]
- ^ van der Heide, T.; van Nes, E. H.; van Katwijk, M. M.; Olff, H.; Smolders, A. J. P. (2011). Romanuk, Tamara (ed.). "Positive feedbacks in seagrass ecosystems: evidence from large-scale empirical data". PLOS ONE. 6 (1): e16504. Bibcode:2011PLoSO...616504V. doi:10.1371/journal.pone.0016504. PMC 3025983. PMID 21283684.
- ^ "Mangal (Mangrove)". Mildred E. Mathias Botanical Garden. Retrieved 11 July 2013.
- ^ "Coastal Salt Marsh". Mildred E. Mathias Botanical Garden. Retrieved 11 July 2013.
- ^ "Facts and figures on marine biodiversity". Marine biodiversity. UNESCO. 2012. Retrieved 11 July 2013.
- ^ Russell, F. S.; Yonge, C. M. (1928). The Seas. Frederick Warne. pp. 225–227.
- ^ Voss, Maren; Bange, Hermann W.; Dippner, Joachim W.; Middelburg, Jack J.; Montoya, Joseph P.; Ward, Bess (2013). "The marine nitrogen cycle: recent discoveries, uncertainties and the potential relevance of climate change". Philosophical Transactions of the Royal Society B. 368 (1621): 20130121. doi:10.1098/rstb.2013.0121. PMC 3682741. PMID 23713119.
- ^ Stow, Dorrik (2004). Encyclopedia of the Oceans. Oxford University Press. ISBN 978-0-19-860687-1.
- ^ Giri C, Ochien E, Tieszen LL, Zhu Z, Singh A, Loveland T, et al. (2011) "지구 관측 위성 데이터를 이용한 세계 맹그로브 숲의 현황과 분포"지구생태학과 생물지리학, 20(1):154~159.doi:10.1111/j.1466-8238.00584.x
- ^ Thomas, N., Lucas, R., Bunting, P., Hardy, A., Rosenqvist, A. 및 Simard, M.(2017) "글로벌 맹그로브 숲 변화의 분포와 추진 요인, 1996-2010"PLOS ONE, 12(6): e0179302.doi:10.1371/journal.pone.0179302
- ^ 줄여서, F.T.와 Frederick, T. (2003) 캘리포니아 대학 출판부, 24페이지.ISBN 9780520240476
- ^ Froise, Rainer; Pauly, Daniel (ed.) (2009).FishBase의 "Phycodurus eques"입니다.2009년 7월판
- ^ a b c d e f g h Tanioka, 타츠로;마쓰모토, Katsumi(2020년)."해양 식물성 플랑크톤 C의 환경 운전에 대한 메타 분석:N:P".Biogeosciences.17(11):2939–2954.Bibcode:2020BGeo...17.2939T. doi:10.5194/bg-17-2939-2020.S2CID 226197209.수정 텍스트는 창조적 공용 귀인 4.0국제 라이센스 하에 가능하다 이 원본에서 복사되었다.
- ^ Broecker, Wallace S. (1982). "Ocean chemistry during glacial time". Geochimica et Cosmochimica Acta. 46 (10): 1689–1705. Bibcode:1982GeCoA..46.1689B. doi:10.1016/0016-7037(82)90110-7.
- ^ Lenton, Timothy M.; Watson, Andrew J. (2000). "Redfield revisited: 1. Regulation of nitrate, phosphate, and oxygen in the ocean". Global Biogeochemical Cycles. 14 (1): 225–248. Bibcode:2000GBioC..14..225L. doi:10.1029/1999GB900065. S2CID 95940597.
- ^ 레드필드 A. C.(1958) "환경 내 화학적 인자의 생물학적 방제", 미국 과학자, 46(3) 230A–221.
- ^ Tyrrell, Toby (1999). "The relative influences of nitrogen and phosphorus on oceanic primary production". Nature. 400 (6744): 525–531. Bibcode:1999Natur.400..525T. doi:10.1038/22941. S2CID 4325136.
- ^ Bonachela, Juan A.; Klausmeier, Christopher A.; Edwards, Kyle F.; Litchman, Elena; Levin, Simon A. (2016). "The role of phytoplankton diversity in the emergent oceanic stoichiometry". Journal of Plankton Research. 38 (4): 1021–1035. doi:10.1093/plankt/fbv087.
- ^ Garcia, Catherine A.; Baer, Steven E.; Garcia, Nathan S.; Rauschenberg, Sara; Twining, Benjamin S.; Lomas, Michael W.; Martiny, Adam C. (2018). "Nutrient supply controls particulate elemental concentrations and ratios in the low latitude eastern Indian Ocean". Nature Communications. 9 (1): 4868. Bibcode:2018NatCo...9.4868G. doi:10.1038/s41467-018-06892-w. PMC 6242840. PMID 30451846.
- ^ Martiny, Adam C.; Pham, Chau T. A.; Primeau, Francois W.; Vrugt, Jasper A.; Moore, J. Keith; Levin, Simon A.; Lomas, Michael W. (2013). "Strong latitudinal patterns in the elemental ratios of marine plankton and organic matter". Nature Geoscience. 6 (4): 279–283. Bibcode:2013NatGe...6..279M. doi:10.1038/ngeo1757. S2CID 5677709.
- ^ Geider, Richard; La Roche, Julie (2002). "Redfield revisited: Variability of C:N:P in marine microalgae and its biochemical basis". European Journal of Phycology. 37: 1–17. doi:10.1017/S0967026201003456. S2CID 13747201.
- ^ Moreno, Allison R.; Martiny, Adam C. (2018). "Ecological Stoichiometry of Ocean Plankton". Annual Review of Marine Science. 10: 43–69. Bibcode:2018ARMS...10...43M. doi:10.1146/annurev-marine-121916-063126. PMID 28853998.
- ^ Boyd, Philip W.; Strzepek, Robert; Fu, Feixue; Hutchins, David A. (2010). "Environmental control of open-ocean phytoplankton groups: Now and in the future". Limnology and Oceanography. 55 (3): 1353–1376. Bibcode:2010LimOc..55.1353B. doi:10.4319/lo.2010.55.3.1353. S2CID 15511444.
- ^ Finkel, Z. V.; Beardall, J.; Flynn, K. J.; Quigg, A.; Rees, T. A. V.; Raven, J. A. (2010). "Phytoplankton in a changing world: Cell size and elemental stoichiometry". Journal of Plankton Research. 32: 119–137. doi:10.1093/plankt/fbp098.
- ^ Van De Waal, Dedmer B.; Verschoor, Antonie M.; Verspagen, Jolanda MH; Van Donk, Ellen; Huisman, Jef (2010). "Climate-driven changes in the ecological stoichiometry of aquatic ecosystems". Frontiers in Ecology and the Environment. 8 (3): 145–152. doi:10.1890/080178. hdl:20.500.11755/c74d7e45-762a-4e66-aa31-844e96e69fa2.
- ^ Irwin, Andrew J.; Finkel, Zoe V.; Müller-Karger, Frank E.; Troccoli Ghinaglia, Luis (2015). "Phytoplankton adapt to changing ocean environments". Proceedings of the National Academy of Sciences. 112 (18): 5762–5766. Bibcode:2015PNAS..112.5762I. doi:10.1073/pnas.1414752112. PMC 4426419. PMID 25902497.
- ^ Collins, Sinéad; Boyd, Philip W.; Doblin, Martina A. (2020). "Evolution, Microbes, and Changing Ocean Conditions". Annual Review of Marine Science. 12: 181–208. Bibcode:2020ARMS...12..181C. doi:10.1146/annurev-marine-010318-095311. PMID 31451085. S2CID 201730744.
- ^ Ward, B. A.; Collins, S.; Dutkiewicz, S.; Gibbs, S.; Bown, P.; Ridgwell, A.; Sauterey, B.; Wilson, J. D.; Oschlies, A. (2019). "Considering the Role of Adaptive Evolution in Models of the Ocean and Climate System". Journal of Advances in Modeling Earth Systems. 11 (11): 3343–3361. Bibcode:2019JAMES..11.3343W. doi:10.1029/2018MS001452. PMC 6988444. PMID 32025278.
- ^ Gurevitch, Jessica; Koricheva, Julia; Nakagawa, Shinichi; Stewart, Gavin (2018). "Meta-analysis and the science of research synthesis". Nature. 555 (7695): 175–182. Bibcode:2018Natur.555..175G. doi:10.1038/nature25753. PMID 29517004. S2CID 3761687.
- ^ Geider, Richard; La Roche, Julie (2002). "Redfield revisited: Variability of C:N:P in marine microalgae and its biochemical basis". European Journal of Phycology. 37: 1–17. doi:10.1017/S0967026201003456. S2CID 13747201.
- ^ Persson, Jonas; Fink, Patrick; Goto, Akira; Hood, James M.; Jonas, Jayne; Kato, Satoshi (2010). "To be or not to be what you eat: Regulation of stoichiometric homeostasis among autotrophs and heterotrophs". Oikos. 119 (5): 741–751. doi:10.1111/j.1600-0706.2009.18545.x.
- ^ Hillebrand, Helmut; Steinert, Georg; Boersma, Maarten; Malzahn, Arne; Meunier, Cédric Léo; Plum, Christoph; Ptacnik, Robert (2013). "Goldman revisited: Faster-growing phytoplankton has lower N : P and lower stoichiometric flexibility". Limnology and Oceanography. 58 (6): 2076–2088. Bibcode:2013LimOc..58.2076H. doi:10.4319/lo.2013.58.6.2076. S2CID 55496787.
- ^ Yvon-Durocher, Gabriel; Dossena, Matteo; Trimmer, Mark; Woodward, Guy; Allen, Andrew P. (2015). "Temperature and the biogeography of algal stoichiometry". Global Ecology and Biogeography. 24 (5): 562–570. doi:10.1111/geb.12280.
- ^ MacIntyre, Hugh L.; Kana, Todd M.; Anning, Tracy; Geider, Richard J. (2002). "Photoacclimation of Photosynthesis Irradiance Response Curves and Photosynthetic Pigments in Microalgae and Cyanobacteria1". Journal of Phycology. 38: 17–38. doi:10.1046/j.1529-8817.2002.00094.x. S2CID 29301640.
- ^ Thrane, Jan‐Erik; Hessen, Dag O.; Andersen, Tom (2016). "The impact of irradiance on optimal and cellular nitrogen to phosphorus ratios in phytoplankton". Ecology Letters. 19 (8): 880–888. doi:10.1111/ele.12623. PMID 27250733.
- ^ Moreno, Allison R.; Martiny, Adam C. (2018). "Ecological Stoichiometry of Ocean Plankton". Annual Review of Marine Science. 10: 43–69. Bibcode:2018ARMS...10...43M. doi:10.1146/annurev-marine-121916-063126. PMID 28853998.
- ^ Hassani, M.A., Duran, P. 및 Hacquard, S. (2018) "발전소 홀로비온트 내 미생물 상호작용"마이크로바이옴, 6 (1): 58.doi:10.118/s40168-0445-0. 수정된 텍스트는 Creative Commons Attribution 4.0 International License에 따라 제공되는 이 원본에서 복사되었습니다.
- ^ Lucking, R., Huhndorf, S., Pister, D.H., Plata, E.R. 및 H.T. Lumbsch, H.T.(2009) "풍기는 제대로 진화했다."마이콜로지아, 101(6): 810-822.도이:10.3852/09-016.
- ^ Heckman, D.S., Geiser, D.M., Eidell, B.R., Stauffer, R.L., Kardos, N.L. 및 Hedges, S.B.(2001) "균류와 식물에 의한 토지 조기 식민지화에 대한 분자적 증거"과학, 293 (5532) : 1129 – 1133 . doi : 10 . 1126 / science . 1061457 .
추가 정보
- Falkowski, Paul (Ed.) (2013) Sea Springer의 1차 생산성.ISBN 9781468438901.
- Falkowski, Paul and Raven, John A. (2013) 수생 광합성 제2판 개정, 프린스턴 대학 출판부.ISBN 9781400849727.
- Falkowski P와 Knoll AH(2011) 바다 학술 출판의 주요 생산자의 진화.ISBN 9780080550510.
- Kirk, John T. O. (2010) 빛과 광합성 in Aquitical Ecosystems 제3판 개정, 케임브리지 대학 출판부.ISBN 9781139493918.




