해양식품 거미줄
Marine food web
| 다음에 대한 일련의 개요의 일부 |
| 해양생물 |
|---|
| |
지구환경과 비교했을 때, 해양환경은 기초에서 뒤집혀진 바이오매스 피라미드를 가지고 있다. 특히 1차 생산자의 바이오매스보다 소비자(복사기, 크릴, 새우, 포리지 어류)의 바이오매스가 더 크다. 이것은 바다의 1차 생산자들이 빠르게 성장하고 번식하는 작은 식물성 플랑크톤이기 때문에 작은 질량이 1차 생산의 빠른 속도를 가질 수 있기 때문이다. 이와는 대조적으로, 성숙한 숲과 같은 많은 중요한 지상 1차 생산자들은 천천히 성장하고 번식하기 때문에 동일한 1차 생산 비율을 달성하려면 훨씬 더 큰 질량이 필요하다.
이러한 반대로 해양 동물 바이오매스의 대부분을 차지하는 것은 동물성 플랑크톤이다. 1차 소비자로서, 동물성 플랑크톤은 1차 생산자(주로 식물성 플랑크톤)와 나머지 해양 식품 웹 (2차 소비자) 사이의 중요한 연결고리다.[1]
만약 식물성 플랑크톤이 먹기 전에 죽는다면, 그것은 해양 눈의 일부로서 유향적 영역을 통해 내려와서 바다 속 깊은 곳으로 정착한다. 이렇게 해서 식물성 플랑크톤은 매년 약 20억 톤의 이산화탄소를 바다로 내보내고, 해양은 모든 격리된 이산화탄소의 약 90%를 차지하는 이산화탄소의 싱크대가 된다.[2] 바다는 세계 산소의 약 절반을 생산하고 대기보다 50배 많은 이산화탄소를 저장한다.[3]
생태계는 그것의 먹이 그물이 물질과 에너지의 흐름을 어떻게 결정하는지 알지 않고는 이해할 수 없다. 식물성 플랑크톤은 무기 화합물을 유기 화합물로 전환하여 생물총량을 자동 생산한다. 이렇게 해서 식물성 플랑크톤은 해양의 다른 모든 생명체를 지원함으로써 해양 먹이 그물의 기초 역할을 한다. 해양 먹이 거미줄의 두 번째 중심 과정은 미생물 루프다. 이 고리는 해양세균과 고고학을 분해하고 유기물과 무기질을 추억하며, 펠라릭 푸드 그물 안에 있거나 해저에 해양 침전물로 침전시켜 제품을 재활용한다.[4]
먹이 사슬과 영양 수준
| 해양 먹이사슬 (iii) |
|---|
| |
| ↓ |
| 식물성 플랑크톤 |
| ↓ |
| 초식 동물성 플랑크톤 |
| ↓ |
| 육식 동물성 플랑크톤 |
| ↓ |
| |
| ↓ |
| |
먹이사슬은 먹이사슬로 만들어진다. 바다의 모든 형태의 생명체는 다른 생명체의 먹이가 될 잠재력을 가지고 있다. 바다에서 먹이 사슬은 일반적으로 식물성 플랑크톤에 동력을 공급하는 태양의 에너지로 시작하여 다음과 같은 과정을 따른다.
식물성 플랑크톤 → 초식성 동물성 플랑크톤 → 육식 동물성 플랑크톤 → 필터피더 → 포식성 척추동물
식물성 플랑크톤은 다른 유기체들이 음식을 위해 필요하지 않다. 왜냐하면 그들은 햇빛을 에너지원으로 사용하여 무기 탄소로부터 직접 음식을 제조할 수 있기 때문이다. 이 과정을 광합성이라고 하며, 식물성 플랑크톤이 자연적으로 발생하는 탄소를 원형으로 변환시키는 결과를 낳는다. 이 때문에 식물성 플랑크톤은 해저의 1차 생산국이나 해양 먹이 사슬의 1차 생산국이라고 한다. 그들은 첫 번째 레벨이기 때문에 영양 수준이 1이라고 한다. 식물성 플랑크톤은 동물성 플랑크톤이라고 불리는 미세한 동물들에 의해 먹이 사슬의 다음 영양 수준에서 소비된다.
동물성 플랑크톤은 먹이 사슬에서 두 번째 영양 수준을 형성하며, 원생동물이라 불리는 미세한 단세포 생물뿐만 아니라 요각류, 크릴과 같은 작은 갑각류, 물고기, 오징어, 바닷가재, 게의 애벌레도 포함한다. 이 수준의 유기체는 1차 소비자로 생각할 수 있다.
차례로, 더 작은 초식성 동물성 플랑크톤은 더 큰 포식성 원생동물과 크릴과 같은 더 큰 육식성 동물성 플랑크톤과 작은 어류, 교육용, 여과용 물고기에 의해 소비된다. 이것은 먹이사슬에서 세 번째 영양분을 구성한다.


네 번째 영양 수준은 포획 어류를 소비하는 포식어, 해양 포유류, 바닷새 등으로 구성된다. 예로는 황새치, 물개, 간넷 등이 있다.
바다표범을 먹을 수 있는 오르카, 황어를 먹을 수 있는 숏핀 마코 상어 등 에이펙스 포식자가 5분의 1의 영양 수준을 이룬다. 고래고래는 동물성 플랑크톤과 크릴을 직접 섭취할 수 있어 영양가가 3, 4단계에 불과한 먹이 사슬로 이어진다.
실제로 영양 수준은 보통 단순한 정수가 아니다. 동일한 소비자 종이 종종 한 가지 이상의 영양 수준에 걸쳐 먹이를 주기 때문이다.[8][9] 예를 들어, 큰 해양 척추동물은 작은 포식성 어류를 먹을 수 있지만 필터 사료도 먹을 수 있다. 가오리는 갑각류를 먹지만 망치머리는 갑각류와 가오리를 모두 먹는다. 동물들도 서로를 먹을 수 있다; 대구는 가재뿐만 아니라 더 작은 대구를 먹고, 가재는 대구 유충을 먹는다. 어린 동물의 먹이 습관과 그 결과 영양 수준은 자라면서 변할 수 있다.
수산과학자 다니엘 폴리(Daniel Pauli)는 영양수준의 값을 1차 생산자와 디트리투스의 1개, 초식동물과 디트리보어의 2개(1차 소비자), 2차 소비자 3개 등으로 설정한다. 모든 소비자 종에 대한 영양 수준 TL의 정의는 다음과 같다.[10]

여기서 는 먹이 j의 부분 영양 수준이며, C 는 i의 식단에서 j의 부분을 나타낸다. 해양생태계의 경우 대부분의 어류를 비롯한 해양소비자의 영양수준이 2.0~5.0으로 평가된다. 비록 북극곰이나 범고래와 같은 해양 포유류의 꼭지점 포식자로부터 발생하지만,[11] 이 상한값인 5.0은 큰 물고기에게도 특이하다.[12] 대조적으로, 인간의 평균 영양 수준은 약 2.21로 돼지나 멸치와 거의 같다.[13][14]


택슨으로
1차 생산자

해양 먹이 그물의 밑부분에는 단세포 조류와 식물성 플랑크톤으로 알려진 식물성 유기체들이 있다. 식물성 플랑크톤은 형태학, 크기, 색소 유형에 근거한 분류학 그룹의 다양한 조합으로 나뉜 미세한 자생물의 그룹이다. 해양 식물성 플랑크톤은 주로 광자극으로 태양빛 지표수에 서식하며, 탄소를 고정하고 산소를 생산하기 위해서는 햇빛뿐만 아니라 질소, 인 등의 영양소가 필요하다. 그러나 일부 해양 식물성 플랑크톤은 황화수소, 철철, 암모니아와 같은 무기 전자원을 사용하는 화학 자생물로 심해 환기구 근처에 서식한다.[16]
해양 식물성 플랑크톤은 해양식품 거미줄의 기초를 이루고 있으며, 광합성에[17] 의한 전 세계 탄소 고정과 산소 생산의 약 절반을 차지하고 있으며, 전 세계 탄소 순환의 핵심 연결고리다.[18] 육지의 식물처럼 식물성 플랑크톤은 엽록소와 다른 경증하는 색소를 사용하여 광합성을 수행하며 대기 중의 이산화탄소를 흡수하여 연료를 위한 당분을 생산한다. 물 속의 엽록소는 물이 햇빛을 반사하고 흡수하는 방식을 변화시켜 과학자들이 식물성 플랑크톤의 양과 위치를 파악할 수 있게 한다. 이러한 측정은 과학자들에게 해양 환경의 건강에 대한 귀중한 통찰력을 주고, 과학자들이 해양 탄소 순환을 연구하는데 도움을 준다.[15]

식물성 플랑크톤 중에는 시아노박테리아라고 불리는 박테리아의 망울에 속하는 사람들이 있다. 해양 시아노박테리아는 알려진 가장 작은 광합성 유기체를 포함한다. 가장 작은 프로클로로코커스(Proclorococcus)는 가로 0.5~0.8마이크로미터에 불과하다.[19] 개별적인 숫자로 볼 때 프로클로로코쿠스는 아마도 지구상에서 가장 풍부한 종일 것이다: 1밀리리터의 표면 바닷물은 10만개 이상의 세포를 포함할 수 있다. 전세계적으로 수십억27 명의 사람들이 있을 것으로 추정된다.[20] 프로클로로코쿠스는 40°N에서 40°S 사이에 어디에나 있으며 해양의 과두영역(영양빈곤층) 지역에서 지배하고 있다.[21] 박테리아는 지구 대기 중 산소의 약 20%를 차지한다.[22]
- 식물성 플랑크톤은 해양 먹이 사슬의 밑부분을 형성한다.

식물성 플랑크톤

디노플라겔라테

디아톰스



바다에서 대부분의 1차 생산은 조류에 의해 이루어진다. 대부분의 1차 생산은 혈관 식물에 의해 이루어지는 육지와는 대조적이다. 조류는 단일 부유세포부터 부착된 해조류까지 다양하며, 혈관식물은 해초, 맹그로브 등의 집단에 의해 바다에서 대표된다. 해초나 해조류와 같은 더 큰 생산자는 대부분 연근대나 얕은 물에 국한되어 있는데, 그 곳에서 그들은 밑바닥 기질에 붙어 있고 여전히 광자대 안에 있다. 그러나 조류에 의한 1차 생산의 대부분은 식물성 플랑크톤에 의해 이루어진다.
따라서, 해양 환경에서, 첫 번째 하위 영양 수위는 주로 바다에 떠 있는 미세한 표류 생물인 식물성 플랑크톤에 의해 점유된다. 대부분의 식물성 플랑크톤은 너무 작아서 도움을 받지 않은 눈으로 개별적으로 볼 수 없다. 그것들은 충분히 많은 수의 물이 존재할 때 (흔히 초록색) 변색으로 나타날 수 있다. 그들은 광합성을 통해 생물량을 늘리기 때문에 바다의 태양빛 표면층(읍호테틱스 존)에서 산다.
식물성 플랑크톤의 가장 중요한 그룹은 디아톰과 디노플라겔라테스 이다. 일부 추정에 따르면, 디아톰은 전체 해양 생산의 45%까지 기여한다고 한다.[23] 일부 종은 길이가 2밀리미터에 이를 수 있지만, 규조류는 보통 미세하다.
1차 소비자

두 번째 영양 수준(주요 소비자)은 식물성 플랑크톤을 먹고 사는 동물성 플랑크톤이 차지하고 있다. 식물성 플랑크톤과 함께, 그들은 세계의 대부분의 거대한 어장을 지탱하는 먹이 피라미드의 기초를 형성한다. 많은 동물성 플랑크톤은 해양 지표수에서 식물성 플랑크톤과 함께 발견되는 작은 동물로, 작은 갑각류, 물고기 애벌레, 튀김(최근 부화한 물고기) 등이 있다. 대부분의 동물성 플랑크톤은 필터 공급자로, 그들은 물에 있는 식물성 플랑크톤에 무리를 주기 위해 첨가제를 사용한다. 몇몇 더 큰 동물성 플랑크톤은 또한 더 작은 동물성 플랑크톤을 먹고 산다. 어떤 동물성 플랑크톤은 포식자들을 피하기 위해 약간 뛰어다닐 수 있지만, 그들은 실제로 수영을 할 수 없다. 식물성 플랑크톤처럼 그들은 대신 조류, 조수, 바람과 함께 떠다닌다. 동물성 플랑크톤은 빠르게 번식할 수 있고, 그들의 개체수는 좋은 조건에서 하루에 30퍼센트까지 증가할 수 있다. 많은 사람들이 짧고 생산적인 삶을 살고 빨리 성숙에 도달한다.
과점 부자들은 목줄과 옷깃처럼 배열된 두드러진 구강 섬유를 가진 섬유의 집단이다. 그것들은 해양 플랑크톤 공동체에서 매우 흔하며, 보통 밀리리터 당 약 1개의 농도로 발견된다. 그들은 바다에서 가장 중요한 초식동물이며, 먹이 사슬의 첫 번째 연결고리다.[24]
동물성 플랑크톤의 다른 특히 중요한 그룹들은 요각류와 크릴이다. 요각류는 바다와 민물 서식지에서 발견되는 작은 갑각류 동물군이다. 그들은 바다에서 단백질의 가장 큰 공급원이며,[25] 물고기를 잡는데 중요한 먹잇감이 된다. 크릴은 다음으로 단백질의 가장 큰 원천이다. 크릴은 특히 더 작은 동물성 플랑크톤을 먹는 포식자 동물성 플랑크톤이다. 이것은 그들이 정말로 세 번째 영양수준인 2차 소비자들, 그리고 포획물고기와 함께 속한다는 것을 의미한다.
- 동물성 플랑크톤은 해양 먹이 사슬에서 2단계를 형성한다.

분할 웜

작은 새우처럼 생긴 갑각류

어린이 플랑크톤 오징어




식물성 플랑크톤과 동물성 플랑크톤이 함께 바다의 플랑크톤 대부분을 차지한다. 플랑크톤은 바다에 떠 있는 작은 표류 생물(그리스 플랑크토스 = 방랑자 또는 방랑자)에 적용되는 말이다. 정의에 따르면 플랑크톤으로 분류된 유기체는 해류를 거슬러 헤엄칠 수 없다; 그들은 주변 물살에 저항할 수 없고 그들의 위치를 통제할 수 없다. 해양 환경에서는 처음 두 영양 수준이 주로 플랑크톤에 의해 점유된다. 플랑크톤은 생산자와 소비자로 나뉜다. 생산자는 식물성 플랑크톤(그리스 피톤 = 식물)이고, 식물성 플랑크톤을 먹는 소비자는 동물성 플랑크톤(그리스 쿤 = 동물)이다.
해파리는 느리게 헤엄치는 동물이고, 대부분의 종은 플랑크톤의 일부를 형성한다. 전통적으로 해파리는 영양가 있는 막다른 골목으로 여겨져 왔고, 해양 먹이 거미줄의 작은 플레이어들, 해양 선피쉬와 가죽 등 바다 거북과 같은 몇몇 특별한 포식자 외에는 다른 유기체들에게 거의 영양적인 가치나 관심을 주지 않는 물을 주로 기반으로 하는 젤라틴성 유기체들이었다.[27][26] 그 견해는 최근에 도전받았다. 해파리, 그리고 보다 일반적으로 젤라틴이 많은 동물성 플랑크톤은 매우 다양하고, 단단한 부분 없이 연약하며, 보고 감시하기 어렵고, 급격한 인구 변동을 겪으며 종종 해안이나 심해에서 불편하게 산다. 해파리는 먹었을 때 진흙으로 변하고 빠르게 소화되기 때문에 과학자들이 포식자의 내장에서 해파리를 탐지하고 분석하는 것은 어렵다.[27] 그러나 해파리는 엄청난 숫자로 꽃을 피우며 참치, 창어, 황어뿐만 아니라 문어, 해삼, 게, 암페퍼드와 같은 다양한 조류와 무척추동물의 식단에서 주요 성분을 이루고 있는 것으로 나타났다.[28][26] "그들의 낮은 에너지 밀도에도 불구하고, 해파리의 포식자 에너지 예산에 대한 기여도는 빠른 소화, 낮은 포획 비용, 가용성, 그리고 더 많은 에너지가 풍부한 구성품에 대한 선택적 공급으로 인해 추정된 것보다 훨씬 더 클 수 있다. 해파리를 먹이면 해양 포식자가 플라스틱 섭취에 취약해질 수 있다."[26]
고품질의 소비자

- 해양 무척추동물
- 물고기
- 포리지 어류: 포리지 어류는 해양 먹이 그물에서 중심 위치를 차지한다. 그것이 먹는 유기체는 낮은 영양 수준이고, 그것을 먹는 유기체는 더 높은 영양 수준이다. 포획물고기는 먹이 그물에서 중간 수준을 차지하며, 고등어, 바닷새, 포유류의 지배적인 먹이 역할을 한다.[29]
- 프레데터 물고기
- 갈치
- 기타 해양 척추동물
2010년에 연구원들은 고래들이 고래 펌프라고 불리는 과정을 이용하여 해양의 깊은 곳에서 다시 표면으로 영양분을 운반하는 것을 발견했다.[30] 고래들은 크릴이 발견되는 바다에서 더 깊은 수위에서 먹이를 먹지만, 숨을 쉬기 위해 정기적으로 수면으로 돌아온다. 그곳에서 고래들은 질소와 철분이 풍부한 액체를 배설한다. 액체는 가라앉는 대신 식물성 플랑크톤이 소비하는 표면에 머문다. 메인만에서 고래 펌프는 강보다 더 많은 질소를 제공한다.[31]

혹등고래는 물고기를 잡아먹기 위해 아래에서 뛰어다닌다.

,_Belmont_-_geograph.org.uk_-_529175.jpg)

미생물

해양 미생물이 해양 생태계에서 이전에 생각했던 것보다 훨씬 더 큰 역할을 한다는 인식이 최근 증가하고 있다. 메타게노믹스의 발전은 연구자들에게 미생물 세계를 볼 수 있는 강력한 렌즈와 살아있는 세계에 대한 이해를 혁신할 수 있는 잠재력을 제공하면서 이전에 숨겨져 있던 미세한 생물의 다양성을 밝혀낼 수 있는 능력을 준다.[33] 메타바코딩 식이 분석 기법은 식품 웹을 더 높은 수준의 분류학적 분해능으로 재구성하는 데 사용되고 있으며 상호작용의 거미줄에서 더 깊은 복잡성을 드러내고 있다.[34]
미생물은 해양 먹이 그물에서 중요한 역할을 한다. 바이러스 분진통로는 해양 미생물 미립자 유기물질(POM)을 미생물이 쉽게 섭취할 수 있는 용해된 유기물(DOM)로 재활용해 영양수준을 끌어올리는 것을 막는 메커니즘이다.[35] 바이러스 션팅은 해양 미생물 한 종이 미생물 환경을 지배하는 것을 방지함으로써 미생물 생태계 내의 다양성을 유지하는데 도움을 준다.[36] 바이러스성 션트 경로에 의해 재활용된 DOM은 해양 DOM의 다른 주요 공급원이 생성하는 양과 비교된다.[37]
일반적으로 용해된 유기탄소(DOC)는 박테리아 용해, 식물성 플랑크톤으로부터 고정 탄소의 누출 또는 배출, 갑작스러운 세포 노화, 동물성 플랑크톤에 의한 엉성한 먹이, 수생동물에 의한 폐기물 배출, 분해 또는 해체로 인해 해양 환경에 도입된다. 지구상의 식물과 토양에서 나오는 유기 입자의.[38] 미생물 순환에 있는 박테리아는 에너지를 많이 쓰는 물질을 성장시키기 위해 이 입자 디트리투스를 분해한다. 해양생태계 유기물질의 95% 이상이 고분자, 고분자량(HMW) 화합물(예: 단백질, 다당류, 지질)으로 구성되기 때문에 총 용존 유기물질(DOM)의 극히 일부만이 영양 수준이 높은 대부분의 해양생물에 쉽게 활용 가능하다. 이것은 용해된 유기 탄소가 대부분의 해양 유기체에게 직접 공급되지 않는다는 것을 의미한다; 해양 박테리아는 이 유기 탄소를 먹이 그물에 도입하여 영양 수준이 더 높은 추가 에너지를 공급하게 된다.

2011년 인공위성에 의해 이미징된 것처럼.



.jpg)


거대 해양 바이러스인 CroV는 해양 동물원 플레겔레이트 카페테리아 roenbergensis의 용해로 감염되어 사망을 초래한다.[42] 이것은 해안 생태계에 영향을 미친다. 왜냐하면 카페테리아 랑베르겐시스는 물에서 발견되는 박테리아를 먹기 때문이다. 광범위한 CroV 감염으로 인해 카페테리아 roenbergensis의 수가 적을 때, 박테리아의 수는 기하급수적으로 증가한다.[43] C. roenbergensis의 자연 개체군에 대한 CroV의 영향은 여전히 알려져 있지 않다. 그러나, 이 바이러스는 숙주에 매우 특이하며 밀접하게 관련된 다른 유기체들을 감염시키지 않는 것으로 밝혀졌다.[44] 구내식당 랑베르겐시스도 두 번째 바이러스인 '마바이러스 바이로피지'에 감염되는데, 이는 또 다른 특정 바이러스가 존재하는 경우에만 복제할 수 있다는 것을 의미하며, 이 경우 CroV가 존재하는 경우다.[45] 이 바이러스는 C. roenbergensis 세포의 생존으로 이어지는 CroV의 복제를 방해한다. 마바이러스는 C. roenbergensis의 세포 게놈에 통합될 수 있으며, 따라서 개체군에 면역성을 부여한다.[46]
곰팡이


서식지별
펠라직 거미줄

펠릭스 생태계를 위해 레전드르와 라솔자다간은 1995년 초식성 먹이 사슬과 미생물 고리를 식품 웹 최종 구성원으로 하는 영양학적 경로의 연속체를 제안했다.[51] 고전적인 선형 먹이 사슬 최종 구성원은 더 큰 식물성 플랑크톤에서 동물성 플랑크톤에 의해 방목되고 더 큰 동물성 플랑크톤이나 다른 포식자에 의해 동물성 플랑크톤에 의해 그 후에 포식하는 것을 포함한다. 이러한 선형 먹이 사슬에서 포식자는 높은 식물성 플랑크톤 바이오매스(식물성 플랑크톤, 초식동물 및 포식자가 있는 시스템에서) 또는 식물 플랑크톤 바이오매스를 감소시킬 수 있다. 포식자 풍요의 변화는 영양 폭포를 초래할 수 있다.[52] 미생물 루프 엔드 멤버는 기초 자원으로서 식물성 플랑크톤뿐만 아니라 유기 탄소 용해도 포함한다.[53] 용해된 유기탄소는 성장을 위해 이질성 박테리아에 의해 이용되며, 더 큰 동물성 플랑크톤에 의해 예측된다. 결과적으로, 용해된 유기 탄소는 박테리아-마이크로조플랑크톤 루프를 통해 동물성 플랑크톤으로 변환된다. 이 두 최종 멤버 탄소 처리 경로는 여러 수준에서 연결된다. 작은 식물성 플랑크톤은 마이크로조플랑크톤이 직접 섭취할 수 있다.[50]
오른쪽의 도표에서 알 수 있듯이 용해된 유기탄소는 유기탄소의 1차 생산자와 소비자에 의해 다방면으로 그리고 다양한 유기체에 의해 생산된다. 1차 생산자에 의한 DOC 방출은 누출에 의해 수동적으로 발생하며 영양소 제한 중에 불균형 성장 중에 활발히 발생한다.[54][55] 식물성 플랑크톤에서 용해된 유기농 수영장으로 가는 또 다른 직접적인 통로는 바이러스 용해와 관련이 있다.[56] 해양 바이러스는 특히 따뜻하고 낮은 위도의 바다에서 식물성 플랑크톤 사망의 주요 원인이다. 초식동물에 의한 엉성한 먹이 섭취와 소비자들의 불완전한 먹이 소화가 용해된 유기탄소의 또 다른 원천이다. 이질성 미생물들은 세포외 효소를 사용하여 미립자 유기 탄소를 용해하고 이것과 다른 용해된 유기 탄소 자원을 성장과 유지에 사용한다. 미생물 이형성생산의 일부는 마이크로조플랑크톤에 의해 사용된다; 이형성생성계의 또 다른 부분은 강력한 바이러스 투석을 받게 되고 이것은 다시 용해된 유기탄소의 방출을 야기한다. 미생물 루프의 효율성은 여러 요인에 따라 결정되지만 특히 이질성 미생물의 사망률에 대한 포식 및 바이러스 투석의 상대적 중요성에 따라 결정된다.[50]
- 펠라직 푸드 웨브

- 메소플라스틱 푸드 거미줄

중피세포종이 전 세계 탄소 예산에[58] 미치는 영향
DVM = diel 수직 마이그레이션NM = 비 마이그레이션




과학자들은 200미터에서 1,000미터 깊이의 이 중층 마법의 대부분 알려지지 않은 황혼 지대를 더 자세히 탐구하기 시작하고 있다. 이 층은 매년 대기에서 약 40억 톤의 이산화탄소를 제거하는 역할을 한다. 중간층에는 해양 어류 바이오매스의 대부분이 서식하고 있다.[59]
2017년 한 연구에 따르면, 나르코메두새는 중피성 먹이를 가장 많이 소비하고, 물리 사이포노포레스, 두족류, 두족류 등이 그 뒤를 이었다. 이른바 '젤리 웹'의 중요성은 이제 겨우 이해되기 시작했지만 맹수 물고기나 오징어와 유사한 생태학적 영향을 받는 깊은 펠리릭 푸드 웹에서 메두새, 카테노포어, 사이포노포어가 핵심 포식자가 될 수 있을 것으로 보인다. 전통적으로 젤라틴이 많은 포식자들은 해양 영양의 공급자가 아니라고 생각되었지만, 그들은 깊은 펠라믹 먹이 그물에서 실질적이고 필수적인 역할을 하는 것으로 보인다.[60] 중요한 활성 수송 메커니즘인 디엘 수직 이주는 메소즈 플랑크톤이 대기에서 이산화탄소를 분리시킬 수 있을 뿐만 아니라 다른 메소펠러성 유기체에 탄소 필요량을 공급할 수 있게 해준다.[61]
2020년까지 온실가스의 배출을 줄이더라도 2050년까지 지구온난화가 지금보다 7배 빠른 속도로 심해로 확산될 수 있다는 연구결과가 나왔다. 해양 생물 종들이 생존 온도에서 머물기 위해 움직여야 하기 때문에, 중극성과 더 깊은 층에서 온난화는 심해 먹이 그물에 큰 영향을 미칠 수 있다.[62][63]
- 황혼 속의 물고기는 2014년 2월 10일 해양 생태계에 새로운 빛을 던졌다.
- 2015년 6월 29일 뉴욕 타임즈의 해양 미스터리.
- 메소펠러릭 어류 -[64][65] 2010년 말라스피나 항법 탐험

바다 표면에서

바다 표면 서식지는 바다와 대기의 접점에 있다. 바다 표면의 생물필름과 같은 서식지는 흔히 네우스턴이라고 불리는 표면적 질식 미생물을 서식한다. 이 거대한 공기-물 인터페이스는 지구 표면적의 70% 이상을 차지하는 주요 공기-물 교환과정의 교차점에 위치한다. 박테리오네우스턴이라 불리는 해양 표면 미세층 박테리아는 온실가스의 공기-해상 가스 교환, 기후-활성 마린 생산과 같은 실용적인 응용으로 인해 관심을 끌고 있다.에어로졸과 바다의 원격감각.[67] 특정 관심사는 미생물 생화학 공정을 통한 계면활성제(표면 활성물질)의 생산과 분해다. 공해상에 존재하는 계면활성제의 주요 공급원으로는 식물성 플랑크톤,[68] 지상유출, 대기권 침적 등이 있다.[67]
색조류와는 달리 계면활성제 관련 박테리아는 해양 색채 이미지에서 보이지 않을 수 있다. 합성개구경 레이더를 사용하여 이러한 "보이지 않는" 계면활성제 관련 박테리아를 탐지할 수 있는 능력을 갖추는 것은 구름, 안개, 대낮에 관계없이 전천후 조건에서 엄청난 이점을 가진다.[67] 이것은 매우 강한 바람에서 특히 중요하다. 왜냐하면 이것들은 가장 격렬한 공해 가스 교환과 해양 에어로졸 생산이 일어나는 조건이기 때문이다. 따라서, 컬러 위성 사진 외에도, SAR 위성 사진은 해양과 대기의 경계에서 생물물리학적 과정, 공해 온실가스 교환 및 기후 활성 해양 에어로졸 생산에 대한 글로벌 그림에 대한 추가적인 통찰력을 제공할 수 있다.[67]
해저에서

해저(벤티크) 서식지는 바다와 지구 내부의 접점에 위치한다.
- 배액 및 환기구

해안망
연안에는 강과 대륙붕 위의 물이 포함된다. 그들은 전체 해양 면적의 약 8%를 차지하며 전체 해양 생산성의 약 절반을 차지한다. 영농화를 결정하는 주요 영양소는 해안가의 질소와 호수의 인이다. 둘 다 주변 바다나 인접 호수의 비료 역할을 하는 과노(seabird shat)에서 고농도로 발견된다. 요산은 지배적인 질소 화합물로, 광물화 과정에서 다른 질소 형태가 생성된다.[74]
생태계는, 심지어 겉으로 보기에 뚜렷한 경계를 가진 것들도, 다른 인접한 시스템들과 독립적으로 기능하는 경우는 거의 없다.[75] 생태학자들은 에너지와 영양소의 교차생태계 수송이 식물과 동물의 개체수와 공동체에 미치는 중요한 영향을 점점 더 인식하고 있다.[76][77] 잘 알려진 예로는 바닷새들이 어떻게 해양에서 유래한 영양소를 10% 인뿐만 아니라, 질소(N)가 최대 15~20% 함유된 배설물(과노) 형태로 번식 섬에 집중시키느냐 하는 것이다.[78][79][80] 이러한 영양소는 지상 생태계의 기능과 역동성을 극적으로 변화시키고 일차적, 이차적 생산성 증가를 지원할 수 있다.[81][82] 그러나 여러 분류학 그룹에 걸친 과노 퇴적으로 인해 많은 연구가 지상부품의 질소농축이 입증되었지만,[81][83][84][85] 해양생태계에 대한 역반응을 연구한 것은 극히 일부에 불과했고, 이러한 연구들의 대부분은 온대지방과 고 영양수에 국한되었다.[78][86][87][88] 열대지방에서는 산호초가 바다새 번식 인구가 많은 섬과 인접한 곳에서 발견될 수 있으며, 주변 해역에서 바닷새에서 유래한 영양소가 운반되어 국지적 영양농축에 영향을 받을 가능성이 있다. 과노가 열대 해양 생태계에 미치는 영향에 관한 연구는 과노에서 추출한 질소가 바닷물과 암초 1차 생산자를 풍부하게 만든다는 것을 시사한다.[86][89][90]
산호초 건설은 필수적인 질소 필요를 가지고 있으며, 질소가 일차 생산성의 주요 제한 영양소인 영양소가 부족한 열대 수역에서 번성하면서,[92] 그들은 이 원소를 보존하기 위한 특정한 적응을 개발했다. 그들의 설립과 유지관리는 부분적으로 주변 해역에서 용해된 무기질소(암모늄과 질산염)를 차지하고 보존할 수 있는 단세포 디노플라겔라테스, 심비오디닐륨 spp. (zooxanthellae)와 공생한 데 기인한다.[93][94][95] 이 동물원안첼래는 또한 동물 배설물을 재활용할 수 있고, 그 후에 그것들을 아미노산,[96] 암모늄 또는 요소로서 산호 숙주에게 다시 옮길 수 있다.[97] 산호는 또한 질소가 풍부한 침전물 입자와 플랑크톤을 섭취할 수 있다.[100][101] 해안 영양분 공급과 과잉 영양분 공급은 산호류에 강한 영향을 미치며 골격 성장 감소로 이어질 수 있다.[94][102][103][104][90]


오른쪽의 도표에서 (1) 암모니아는 NH3와 NH4+를 생성하며 (2) 질화물은 NH4+ 산화에 의해 NO3-를 생성한다. (3) 알칼리성 조건 하에서, 바닷새 배설물의 전형인 NH4+, (4) NH3는 급속하게 볼륨화하여 NH4+로 변형되며, 이는 군락 밖으로 운반되고, 먼 생태계로 수출되는 습윤을 통해 이루어진다.s, 부영양화된다. 인의 순환은 간단하고 기동성이 떨어졌다. 이 원소는 바닷새 배설물 물질의 많은 화학적 형태에서 발견되지만 가장 이동성이 높고 생물학적으로 이용 가능한 것은 직교인산염(5)으로 지하수나 표피수로 침출될 수 있다.[74]




대륙붕에.





DNA 바코드는 웹 노드에서 더 나은 분류학적 분해능을 가진 식품 웹 구조를 만드는 데 사용될 수 있다. 이것은 누가 누구를 먹는지에 대한 더 구체적인 종 식별과 더 큰 명확성을 제공한다. "DNA 바코드와 DNA 정보는 더 큰 상호작용 웹의 구축에 새로운 접근법을 허용하고 적절한 표본 크기를 달성하는 데 몇 가지 장애물을 극복할 수 있다."[34]
종 식별을 위해 새롭게 적용된 방법은 DNA 메타베이스코딩이다. 형태학을 통한 종 식별은 상대적으로 어렵고 많은 시간과 전문지식이 필요하다.[115][116] 높은 처리량 염기서열 DNA 메타코딩은 분류학적 할당을 가능하게 하고 따라서 이전의 DNA 증폭을 위해 선택된 그룹별 프라이머에 관한 완전한 샘플에 대한 식별을 가능하게 한다.
- 미생물 DNA 바코드
- 조류 DNA 바코드
- 물고기 DNA 바코드
- 식이요법 평가에서 DNA 바코드
- 다시마 숲
- Byrnes, J.E., Reynolds, P.L., Stachowicz, J.J. (2007) "Invasions and extractions to reform the canal food webs." PLOS ONE, 2(3): e295. doi:10.1371/journal.pone.0000295
폴라 웹


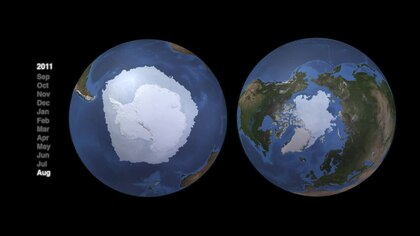
북극과 남극의 해양 시스템은 지형 구조가 매우 다르며 그 결과 음식 거미줄 구조가 매우 다르다.[117] 북극과 남극의 펠라매틱 먹이사슬은 주로 몇 가지 핵심 종에 의해 제어되는 특징적인 에너지 흐름을 가지고 있다. 그러나 어느 쪽에도 하나의 일반적인 웹은 없다. 대체 경로는 복원력과 에너지 흐름 유지를 위해 중요하다. 그러나, 이러한 더 복잡한 대안들은 상위 영양 수준 종에 대한 에너지 흐름을 적게 제공한다. "식품 웹 구조는 지역마다 비슷할 수 있지만, 중간크기의 수준을 지배하는 개별 종은 극지방마다 다르다."[118]



- 북극


북극의 먹이 그물은 복잡하다. 해빙의 손실은 궁극적으로 조류와 플랑크톤에서 물고기, 포유류에 이르기까지 먹이 그물 전체에 영향을 미칠 수 있다. 기후변화가 특정 종에 미치는 영향은 먹이사슬을 통해 파급될 수 있고, 다양한 다른 생물체에도 영향을 미칠 수 있다... 해빙의 감소는 북극곰의 주요 서식지 범위를 줄임으로써 개체수를 손상시킬 뿐만 아니라, 먹이 웹 효과를 통해 그들에게 부정적인 영향을 미치고 있다. 북극의 해빙 기간과 범위가 줄어들면 풍부한 해조류가 감소하게 되는데, 해조류는 얼음의 영양분이 풍부한 주머니에서 번성한다. 이 해조류는 바다표범을 포함한 많은 해양 포유류의 중요한 먹이인 북극 대구가 차례로 먹는 동물성 플랑크톤에 의해 먹혀진다. 바다표범은 북극곰에게 잡아먹힌다. 따라서, 해조류의 감소는 북극곰 개체수의 감소에 기여할 수 있다.[119]
2020년에 연구자들은 지난 20년간 북극해의 1차 생산량 측정에서 식물성 플랑크톤 농도가 높아 60% 가까이 증가했다고 보고했다. 그들은 새로운 영양소가 다른 바다에서 유입되고 있다는 가설을 세우고 이것은 북극해가 미래에 더 높은 영양 수준 생산과 추가적인 탄소 고정을 지원할 수 있다는 것을 의미한다.[120][121]





- 남극

남극해파리 디플룰마리스 얼음밑 남극해파리

해빙이 물러가고 상당한 탄소를 수출하는 로스해의 중요한 식물성 플랑크르인 알가 페이로시스티스 남극대륙의 군락지가 꽃을 피운다.[124]

남극 순환기류에서 발견된 페나테 디아톰 프래길리오피스 케겔렌시스(Fragilariopsis kerguelensis)는 전지구 규산염 펌프의 핵심 동인이다.[125]
.jpg)




다크 박스의 탄소 질량(Gt C)



기초종과 키스톤종
.jpg)

.jpg)
파운데이션 종의 개념은 1972년 폴 K에 의해 소개되었다. 해양 무척추동물과 조류 공동체의 특정 구성원들에게 그것을 적용한 데이톤.[132] 몇몇 지역의 연구를 통해, 활동량이 나머지 해양 공동체에 불균형적인 영향을 미치는 소수의 종들이 있다는 것이 분명했고, 따라서 그들은 공동체의 회복력을 위한 열쇠가 되었다. 데이튼의 견해는 기초 종에 초점을 맞추면 모든 공동체 구성원의 반응을 동시에 추적하는 극도로 어려운 작업을 시도하는 대신, 공동체 전체가 공해와 같은 장애에 어떻게 반응하는지를 보다 신속하게 이해할 수 있는 단순화된 접근법을 허용할 수 있을 것이라는 것이었다.
기초종은 생태계를 구성하고 환경을 형성하며 생태계를 정의하는 지배적인 역할을 하는 종이다. 그러한 생태계는 종종 해초원, 굴밭, 산호초, 다시마 숲, 맹그로브 숲과 같은 기초 종에서 이름을 따왔다.[133] 예를 들어, 붉은 맹그로브는 맹그로브 숲에서 흔한 기초 종이다. 맹그로브의 뿌리는 도미와 같은 어린 물고기들에게 양묘장을 제공한다.[134] 파운데이션 종은 먹이 그물에서 어떤 영양 수준도 차지할 수 있지만 생산자가 되는 경향이 있다.[135]


키스톤 종의 개념은 동물학자 로버트 T. 파인에 의해 1969년에 도입되었다.[136][137] 파인은 불가사리와 홍합을 포함한 (고조선과 저조선의 사이)의 해양 무척추동물들 사이의 관계에 대한 관찰과 실험을 설명하기 위해 이 개념을 개발했다. 어떤 바다별들은 다른 천적이 없는 성게, 홍합, 그리고 다른 조개류를 잡아먹는다. 해성이 생태계에서 제거되면 홍합 개체수가 걷잡을 수 없이 폭발해 다른 종도 대부분 쫓아낸다.[138]
키스톤 종은 생태계 먹이 그물 안에서 그 수에 불균형한 큰 효과를 가진 종이다.[139] 생물총량이나 생산성의 측정에 의해 키스톤 종(種)이 생태계의 작은 부분이었음에도 불구하고, 키스톤 종(種)이 제거되면 생태계는 극적인 변화를 경험할 수 있다.[140] 해달은 다시마 숲에 미치는 성게의 피해를 제한한다. 북미 서부 해안의 수달이 모피 때문에 상업적으로 사냥을 당했을 때, 그 수는 성게 개체수를 통제할 수 없을 정도로 낮은 수준으로 떨어졌다. 우르친들은 차례로 다시마 열매를 너무 무겁게 갈아엎어 다시마 숲이 크게 사라졌고, 거기에 의존하는 모든 종들도 함께 사라졌다. 해달의 재도입으로 다시마 생태계가 복원될 수 있게 됐다.[141][142]
위상학적 위치
영양 상호작용 네트워크는 해양 생태계의 기능에 대한 많은 정보를 제공할 수 있다. 먹이를 주는 습성을 넘어 다양한 유기체의 세 가지 추가 특성(이동성, 크기, 서식지)이 이 영양적인 관점을 보완할 수 있다.[143]

생태계의 적절한 기능을 지속하기 위해서는 1994년 로튼이 질문한 간단한 질문을 더 잘 이해할 필요가 있다. 생태계에서 종은 무엇을 하는가?[144] 생태학적 역할과 먹이 웹 포지션이 독립적이지 않기 때문에 어떤 종족이 다양한 네트워크 포지션을 차지하고 있는지에 대한 의문이 제기될 필요가 있다.[145][143] 키스톤 종을 식별하려는 최초의 시도 이후,[146][147] 먹이 그물에 대한 관심이 있었다.[148][149] 처음에 그들은 최상위 포식자였고, 그 다음에는 식물, 초식동물, 기생충도 있었다고 제안되었다.[150][151] 공동체 생태학과 보존 생물학 둘 다, 그들이 복잡한 영양망에 어디에 있는지 아는 것은 유용할 것이다.[143]
이러한 종류의 네트워크 분석의 예는 해양 먹이 거미줄의 데이터를 바탕으로 도표에 나타나 있다.[152] 그것은 웹 노드의 위상학적 위치와 유기체가 관여하는 이동성 가치 사이의 관계를 보여준다. 웹 노드는 이동성에 따라 형상을 구분하고, (A) 바텀업 그룹(sessile 및 드리프터)과 (B) 그룹을 강조하여 식품 웹 상단의 색상을 구분한다.[143]
유기체의 상대적 중요성은 시간과 공간에 따라 다르며, 큰 데이터베이스를 보는 것은 문제에 대한 일반적인 통찰력을 제공할 수 있다. 다른 종류의 유기체가 서로 다른 유형의 네트워크 위치를 점유하는 경우, 식품 웹 모델링에서 이를 조정하면 더 신뢰할 수 있는 예측이 나올 것이다. 구심성 지수의 각 다른(중간 영양 수준에서 학위 중심적 역할과 친밀감 centrality,[153]핵심과 keystoneness indexes,[154]과 구심성 지수의 대 영양 수준 유사성(대부분의high-centrality 종)[155]와 풍응답 계측을 통해 유기체의 먹이 그물에 비판적으로 중요한 자리를 이해하기 하고 있었다.s 영양 그룹에 특성 데이터를 추가함으로써 이러한 관심을 확장하는 것은 결과의 생물학적 해석에 도움이 된다. 중심성 지수 사이의 관계는 서식지 네트워크를 포함한 다른 네트워크 유형에 대해서도 연구되었다.[156] [157] 대규모 데이터베이스와 새로운 통계 분석을 통해 이와 같은 질문을 재조사하고 지식을 업데이트할 수 있다.[143]
암호 상호 작용

"일반적인 시야에 숨겨져 있는" 상호작용인 암호적 상호작용은 해양 플랑크톤 식품웹 전체에 걸쳐 발생하지만, 현재 확립된 방법에 의해 대부분 간과되고 있는데, 이는 이러한 상호작용에 대한 대규모 데이터 수집이 제한적이라는 것을 의미한다. 그럼에도 불구하고, 현재 증거는 이러한 상호작용 중 일부가 foodweb 역학 및 모델 결과에 지각할 수 있는 영향을 미칠 수 있음을 시사한다. 암호적 상호작용을 모델에 통합하는 것은 영양소나 에너지의 수송과 관련된 상호작용을 위해 특히 중요하다.[158]
이 도표는 혼합생리학, 존재유전학 및 종의 차이, 미생물 교차공급, 보조생리학 및 세포탄소 분할의 다섯 가지 암호적 상호작용에 의해 영향을 받는 물질 유동성, 모집단 및 분자 풀을 보여준다. 이러한 상호작용은 그들이 영향을 미치는 먹이 그물의 영역이 겹치면서 시너지 효과를 낼 수 있다. 예를 들어, 식물성 플랑크톤에 있는 세포 탄소 파티션은 생물 교차 공급에 이용되고 보조생리의 경우에 교환되는 유기 물질의 다운스트림 풀뿐만 아니라 존재생물과 종의 차이에 기초한 먹이 선택에도 영향을 미칠 수 있다.[158]
'수플랑크톤이 식물성 플랑크톤을 소비한다', '생플랑크톤이 무기질 영양소를 섭취한다', '총 1차 생산량이 먹이 그물에 이용 가능한 탄소의 양을 결정한다' 등의 단순화는 과학자들이 수생환경에서 일반적인 상호작용을 설명하고 모형화하는 데 도움을 주었다. 전통적인 방법은 이러한 일반화를 계량화하고 검증하는 데 초점을 맞췄지만, 최근 몇 년간 유전체학, 센서 감지 한계, 실험 방법 및 기타 기술의 급속한 발전은 플랑크톤 커뮤니티 내에서 상호작용의 일반화가 너무 단순할 수 있다는 것을 보여주었다. 이러한 기술 향상은 대량 샘플링 노력과 실험 방법이 그것들에 편중되어 있기 때문에 암호화된 것으로 보이는 많은 상호작용들을 노출시켰다.[158]
복잡성과 안정성

먹이사슬은 포식자-사전 상호작용의 복잡한 네트워크를 조직할 수 있는 프레임워크를 제공한다. 먹이사슬 모델은 먹이사슬의 네트워크다. 각각의 먹이사슬은 알가나 식물과 같은 유기체인 1차 생산자나 자가영양으로부터 시작되는데, 이것은 그들 자신의 음식을 제조할 수 있다. 다음으로 사슬에는 1차 생산자를 먹이는 유기체가 있으며, 사슬은 이런 식으로 연속적인 포식자의 줄로서 계속된다. 각 사슬의 유기체는 1차 생산자로부터 얼마나 많은 연결고리를 제거하느냐에 따라 영양학적 수준으로 분류된다. 사슬의 길이, 즉 영양 수준은 에너지나 영양소가 식물에서 최상위 포식자로 이동하면서 마주치는 종의 수를 측정한 것이다.[161] 식품 에너지는 한 유기체에서 다음 유기체, 그리고 그 다음 유기체로 흘러가고, 각 수준에서 어느 정도의 에너지가 손실된다. 주어진 영양 수준에는 같은 포식자와 먹이를 가진 한 종이나 한 무리의 종이 있을 수 있다.[162]
1927년, 찰스 엘튼은 먹이사슬의 사용에 관한 영향력 있는 종합을 발표했는데, 그 결과 먹이 그물들이 생태학의 중심 개념이 되었다.[163] 1966년 로버트 페인의 인터티드 해안에 대한 실험적이고 서술적인 연구 이후 먹이 그물에 대한 관심이 높아져 먹이 그물의 복잡성이 종의 다양성과 생태학적 안정성을 유지하는 열쇠가 되었음을 시사했다.[164] 로버트 메이와 스튜어트 핌을 포함한 많은 이론 생태학자들은 이 발견과 다른 발견으로 인해 먹이 그물의 수학적 특성을 조사하게 되었다. 이들의 분석에 따르면 복잡한 먹이사슬은 단순한 먹이사슬보다 안정성이 떨어져야 한다.[165]: 75–77 [166]: 64 자연에서 관찰된 식품 웹의 복잡성과 식품 웹 모델의 수학적 취약성 사이의 명백한 역설은 현재 집중적인 연구와 논쟁의 영역이다. 역설은 부분적으로 푸드 웹의 지속성과 푸드 웹의 평형 안정성 사이의 개념적 차이 때문일 수 있다.[165][166]
영양성 폭포는 웹에서 영양성분이 억제되면 먹이 그물에서 발생할 수 있다.
예를 들어, 포식자가 포식자의 풍부함을 줄이거나 행동을 변화시켜 포식자로부터 다음 영양 수치를 방출할 수 있을 정도로 포식자가 포식하는 데 효과적일 경우 상층 계단식 포식 현상이 발생할 수 있다. 하향식 계단식 계단식 계단식 계단식 계단식 계단식 계단식 계단식 계단식 계단식 계단식으로, 최상위 소비자/준비자가 1차 소비자 집단을 제어한다. 결국 1차 생산자 인구는 번성한다. 최상위 포식자를 제거하면 먹이 거미줄의 역학을 변화시킬 수 있다. 이 경우 1차 소비자는 1차 생산자를 과대포화하여 착취하게 된다. 결국 소비자 인구를 지탱할 1차 생산자가 충분하지 않을 것이다. Top-down food web 안정성은 높은 영양수준의 경쟁과 포부에 달려있다. 침습적인 종들은 또한 최상위 포식자가 되거나 제거함으로써 이 폭포를 바꿀 수 있다. 이러한 상호작용이 항상 부정적인 것은 아닐 수 있다. 연구에 따르면, 특정 침입 종이 폭포를 옮기기 시작했으며, 그 결과, 생태계 파괴가 복구되었다.[167][168] 복잡하고 개방적인 대서양 생태계의 폭포의 예는 1980년대와 1990년대에 북서 대서양에서 발생했다. 지속적인 남획에 의해 대서양 대구(Gadus morhua)와 다른 갈린 어류를 제거함으로써, 특히 북대게(Chionoecetes opilio)와 북새우(Pandalus borealis)와 같은 더 작은 어류 및 무척추동물(Pandalus borealis)을 위한 먹이 종의 풍부함을 증가시켰다. 이러한 먹이 종의 풍부함은 간접적인 효과로서 작은 물고기와 무척추동물의 먹이 역할을 하는 동물성 플랑크톤 집단을 변화시켰다.[169] 인간이 사냥과 낚시를 통해 여러 곳에서 해왔듯이, 하향식 폭포는 먹이 그물에서 최상위 포식자를 제거하는 연쇄효과를 이해하는 데 중요할 수 있다.
하위 계단식 계단식에서는 1차 생산자의 인구가 항상 더 높은 영양 수준에서 에너지의 증가/감소를 통제할 것이다. 주요 생산자는 광합성이 필요한 식물, 식물성 플랑크톤, 동물성 플랑크톤이다. 빛도 중요하지만, 1차 생산자 집단은 시스템의 영양소의 양에 의해 변화된다. 이 먹이사슬은 자원의 가용성과 한계에 의존한다. 처음에 많은 양의 영양소가 있다면 모든 인구는 성장을 경험할 것이다.[170][171]
지상 비교

지상 바이오매스 피라미드와 비교했을 때, 수생 피라미드는 일반적으로 기저에 뒤집혀 있다.
.jpg)
.jpg)
해양 환경은 바이오매스 피라미드에 변혁을 일으킬 수 있다. 특히 소비자의 바이오매스(복식기, 크릴, 새우, 포리지 어류)는 일반적으로 1차 생산자의 바이오매스보다 크다. 이는 해양의 1차 생산자들이 대부분 초소형 식물성 플랑크톤으로 빠르게 성장하고 번식하는 r-전략적 특성을 가지고 있기 때문에 작은 질량이 1차 생산의 빠른 속도를 가질 수 있기 때문이다. 이와는 대조적으로, 성숙한 숲과 같은 많은 지상 1차 생산자들은 천천히 성장하고 재생산하는 K-전략적 특성을 가지고 있기 때문에 동일한 1차 생산 비율을 달성하려면 훨씬 더 큰 질량이 필요하다. 생산률을 이를 달성하는 바이오매스의 평균 양으로 나눈 것을 유기체의 생산/바이오매스(P/B)비라고 한다.[173] 생산량은 시간의 단위당 면적당 질량 또는 에너지의 이동량으로 측정된다. 대조적으로, 바이오매스 측정은 단위 면적 또는 부피 당 질량 단위로 이루어진다. P/B 비율은 역 시간 단위(예: 1/개월)를 이용한다. 이 비율은 주어진 영양 수준에서 바이오매스의 양과 비교한 에너지 흐름의 양을 추정할 수 있도록 하며, 영양 수준 사이의 경계를 정할 수 있다. 가장 일반적으로 P/B 비율은 영양 수준과 유기체의 크기가 증가함에 따라 감소하며, 작고 덧없는 유기체는 크고 오래 지속되는 유기체보다 높은 P/B 비율을 가지고 있다.
예: 이 털북숭이 소나무는 수천 년 동안 살 수 있고, 생산량/바이오매스 비율이 매우 낮다. 시아노박테리움 프로클로로코쿠스는 약 24시간 동안 서식하며, 생산/바이오매스 비율이 매우 높다.
바다에서 대부분의 1차 생산은 조류에 의해 이루어진다. 대부분의 1차 생산은 혈관 식물에 의해 이루어지는 육지와는 대조적이다.
해양생태계와[174] 육상생태계의 생산성 비교 | |||
|---|---|---|---|
| 생태계 | 순 1차 생산성 연간 10억 톤 | 총 식물 바이오매스 억 톤 | 회전시간 몇 해 |
마린 | 45–55 | 1–2 | 0.02–0.06 |
지상파 | 55–70 | 600–1000 | 9–20 |

플랑크토닉 해조류나 수생식물 같은 수생 생산자는 지상 생태계의 목질 나무에 존재하는 2차 성장의 큰 축적이 부족하다. 그러나, 그들은 더 큰 생물총량을 지원할 수 있을 만큼 충분히 빠르게 번식할 수 있다. 이것은 피라미드를 뒤집는다. 1차 소비자들은 그들이 소비하는 생산자들보다 더 많은 바이오매스를 축적하는 긴 수명과 느린 성장률을 가지고 있다. 식물성 플랑크톤을 먹는 동물성 플랑크톤은 단지 며칠만 사는 반면, 식물성 플랑크톤을 먹는 동물성 플랑크톤은 몇 주 동안 살고, 동물성 플랑크톤을 먹는 물고기는 몇 년 연속으로 산다.[176] 또한 수생 포식자는 작은 소비자보다 낮은 사망률을 보이는 경향이 있어 피라미드 반전 패턴의 원인이 된다. 개체 구조, 이동률, 먹이 환경 보호 등은 바이오매스가 역류된 피라미드의 또 다른 가능한 원인이다. 그러나 에너지 피라미드는 열역학 제2법칙에 의해 규정되어 있기 때문에 모든 식품 에너지의 원천이 포함된다면 언제나 직립 피라미드 모양을 갖게 될 것이다."[177][178]
생산되는 대부분의 유기 물질은 결국 소비되고 무기질 탄소로 재생된다. 퇴적물을 축적해 매장함으로써 유기물이 보존되는 비율은 연간 0.2억~04억톤에 불과해 전체 생산량의 극히 일부에 해당한다.[50] 전 세계 식물성 플랑크톤 생산량은 연간 약 500억톤, 식물성 플랑크톤 바이오매스는 약 10억톤으로, 1주일의 회전시간을 의미한다. 해양 마크로피티는 전 세계 바이오매스량은 비슷하지만 연간 생산량은 10억톤에 불과해 이직 시간이 1년임을 시사한다.[179] 이러한 높은 이직률(지상 식물의 전지구적 이직률 1~20년)[174]은 꾸준한 생산뿐만 아니라 유기물질의 효율적인 소비도 의미한다. 여러 가지 유기 물질 손실 경로(자체세포와 이단세포에 의한 생산, 방목, 바이러스 투석, 폐경로)가 있지만 결국 모두 호흡과 무기탄소의 방출로 귀결된다.[50]
.jpeg)
인공적 효과

- 남획
- 산성화
프테로포드와 깨지기 쉬운 별들이 함께 북극 먹이 그물의 밑부분을 형성하고 둘 다 산성화에 의해 심각하게 손상된다. pteropods 껍질은 산성화가 증가하면서 용해되고, 부서지기 쉬운 별들은 맹장을 재배할 때 근육량을 잃는다.[181] 게다가 이 부서지기 쉬운 별의 알은 북극 산성화에 따른 예상 조건에 노출되면 며칠 안에 죽는다.[182] 산성화는 북극의 먹이사슬을 파괴할 위험이 있다. 북극해는 빠르게 변화하고 있으며 아라곤산염으로 과포화되는 과정에서 발전하고 있다.[183] 북극의 먹이사슬은 단순한 것으로 여겨지는데, 이는 먹이 사슬에 작은 유기체에서 더 큰 포식자에 이르는 몇 단계가 있다는 것을 의미한다. 예를 들어, pteropods는 "더 큰 플랑크톤, 물고기, 바닷새, 고래 등 많은 상위 포식자들의 주요 먹이 품목"이다.[184]
.jpg)

- 기후변화
"우리의 결과는 미래의 기후 변화가 어떻게 잠재적으로 해양 먹이사슬을 약화시킬 수 있는지 보여주는데, 이는 식품 웹의 단순화와 생산자-소비자간 역학관계의 변화로 이어지며, 이는 둘 다 벤트릭 커뮤니티의 구조에 중요한 영향을 미친다."[185][186]
"...온도는 해저의 1차 식량 생산자(예: 조류)에서 중간 소비자(식충 동물)로, 해양 식량 거미줄의 최상단에 있는 포식자로의 에너지의 중요한 흐름을 감소시킨다. 이러한 에너지 전달의 장애는 잠재적으로 최상위 포식자들의 식량 공급의 감소로 이어질 수 있으며, 이는 다시 이들 먹이 그물 안에 있는 많은 해양 생물들에게 부정적인 영향을 줄 수 있다. 울라씨는 "기후변화로 식물의 생산성이 높아진 상황에서 이는 주로 시아노박테리아(작은 청록조류)가 늘어난 데 따른 것"이라고 설명했다. "그러나 이러한 시아노박테리아들은 대부분 먹을 수 없고 초식동물에 의해 소비되지 않기 때문에 이러한 1차 생산성이 증가했다고 해서 먹이 그물망을 지원하지 않는다. 지구온난화의 영향하에서 생태계가 어떻게 기능하는지를 이해하는 것은 생태연구의 과제다. 해양 온난화에 대한 대부분의 연구는 오직 한 종 또는 몇 종만을 기반으로 한 단순화된 단기적인 실험을 포함한다."[186]


참고 항목
참조
- ^ 미국 에너지부(2008) 탄소 순환 및 생물 정량화 페이지 81, 워크샵 보고서 DOE/SC-108, 미국 에너지 과학부
- ^ Campbell, Mike (22 June 2011). "The role of marine plankton in sequestration of carbon". EarthTimes. Retrieved 22 August 2014.
- ^ 우리는 왜 바다를 신경 써야 하는가? NOAA: National Ocean Service. 업데이트: 2020년 1월 7일. 2020년 3월 1일 검색됨
- ^ 하인리히스, M.E., 모리, C., L.(2020) "수생생물들과 그들의 화학적 환경 사이의 복합적 상호작용" "다양한 관점에서 설명" In: Jungblut S, Liebich V, Bode-Dalby M. (Eds) YOUMASTRA 9-The Oceans: 우리의 연구, 우리의 미래 279-297페이지. 스프링거. 도이:10.1007/978-3-030-20389-4_15.
- ^ Dunne, J.A., Williams, R.J., 그리고 N.D. Martinez(2002) "식품-웹 구조와 네트워크 이론: 연결성과 크기의 역할" 국립과학원 절차, 99(20): 12917–12922. 도이:10.1073/pnas.192407699.
- ^ J.A. Dunne(2006) "음식 웹의 네트워크 구조" In: M Pascual and J. A. Dunne (Eds.) 생태적 네트워크: 구조와 먹이사슬의 역학 관계를 연결, 27-86페이지. ISBN 9780199775057.
- ^ Karlson, A.M., Gorokhova, E, Gårdmark, A, Pekan-Hekim, Z, Casini, M, Albertson, J, Sundelin, B, Karlsson, O., Bergström, L.(2020). "소비자의 생리학적 상태를 발트해에서 먹이 웹 구조와 먹이 가치에 연결" Ambio, 49(2): 391–406. doi:10.1007/s13280-019-01201-1
- ^ 오덤, W. E. Hald, E. J. (1975) "에스타린 맹그로브 공동체의 디트리투스에 기반한 먹이 그물". L. E. 크로닌의 265–286쪽, 에드. 에스투아린 연구. 제1권. 뉴욕 학술신문.
- ^ Pimm, S. L.; Lawton, J. H. (1978). "On feeding on more than one trophic level". Nature. 275 (5680): 542–544. Bibcode:1978Natur.275..542P. doi:10.1038/275542a0. S2CID 4161183.
- ^ Pauly, D.; Palomares, M. L. (2005). "Fishing down marine food webs: it is far more pervasive than we thought" (PDF). Bulletin of Marine Science. 76 (2): 197–211. Archived from the original (PDF) on 14 May 2013.
- ^ Cortés, E. (1999). "Standardized diet compositions and trophic levels of sharks". ICES J. Mar. Sci. 56 (5): 707–717. doi:10.1006/jmsc.1999.0489.
- ^ Pauly, D.; Trites, A.; Capuli, E.; Christensen, V. (1998). "Diet composition and trophic levels of marine mammals". ICES J. Mar. Sci. 55 (3): 467–481. doi:10.1006/jmsc.1997.0280.
- ^ 연구원들은 인간의 영양 수준을 처음으로 계산한다. Phys.org. . 2013년 12월 3일.
- ^ 본혼모, S, 더블로카, L, 르파페, O, 바드, J, 카플란, D.M, 샤소트, E., 니에블라스(A.E.E.E.2013) "세계의 먹이사슬과 인간 영양수준을 먹어 치운다" 국립과학원 절차, 110(51): 20617–20620. doi:10.1073/pnas.1305827110.
- ^ a b 엽록소 NASA 지구 관측소. 2019년 11월 30일에 접속.
- ^ Guo, Ruoyu; Liang, Yantao; Xin, Yu; Wang, Long; Mou, Shanli; Cao, Chunjie; Xie, Ruize; Zhang, Chuanlun; Tian, Jiwei; Zhang, Yongyu (26 September 2018). "Insight Into the Pico- and Nano-Phytoplankton Communities in the Deepest Biosphere, the Mariana Trench". Frontiers in Microbiology. Frontiers Media SA. 9: 2289. doi:10.3389/fmicb.2018.02289. ISSN 1664-302X. PMC 6168665. PMID 30319587.
- ^ Field, C. B. (10 July 1998). "Primary Production of the Biosphere: Integrating Terrestrial and Oceanic Components". Science. 281 (5374): 237–240. doi:10.1126/science.281.5374.237. PMID 9657713.
- ^ Mouw, Colleen B.; Barnett, Audrey; McKinley, Galen A.; Gloege, Lucas; Pilcher, Darren (20 October 2016). "Global ocean particulate organic carbon flux merged with satellite parameters". Earth System Science Data. 8 (2): 531–541. doi:10.5194/essd-8-531-2016. ISSN 1866-3508.
- ^ Kettler GC, Martiny AC, Huang K, Zucker J, Coleman ML, Rodrigue S, Chen F, Lapidus A, Ferriera S, Johnson J, Steglich C, Church GM, Richardson P, Chisholm SW (December 2007). "Patterns and implications of gene gain and loss in the evolution of Prochlorococcus". PLOS Genetics. 3 (12): e231. doi:10.1371/journal.pgen.0030231. PMC 2151091. PMID 18159947.
- ^ Nemiroff, R.; Bonnell, J., eds. (27 September 2006). "Earth from Saturn". Astronomy Picture of the Day. NASA.
- ^ Partensky F, Hess WR, Vaulot D (March 1999). "Prochlorococcus, a marine photosynthetic prokaryote of global significance". Microbiology and Molecular Biology Reviews. 63 (1): 106–27. doi:10.1128/MMBR.63.1.106-127.1999. PMC 98958. PMID 10066832.
- ^ "The Most Important Microbe You've Never Heard Of". npr.org.
- ^ Mann, D. G. (1999). "The species concept in diatoms". Phycologia. 38 (6): 437–495. doi:10.2216/i0031-8884-38-6-437.1.
- ^ Lynn DH (24 June 2008). The Ciliated Protozoa: Characterization, Classification, and Guide to the Literature (3 ed.). Springer. p. 122. ISBN 978-1-4020-8239-9.
- ^ 올덴부르크 칼 폰 오시에츠키 대학 웨이백머신 2009-01-01
- ^ a b c d 헤이스·G.C.·도일·T.K.·J.D. Houghton(2018) "해파리의 영양학적 중요성에 대한 패러다임의 변화?" 생태·진화의 동향, 33(11): 874-884. doi:10.1016/j.tree.2018.09.001
- ^ a b G. 해밀턴, G. (2016) "해파리의 비밀스러운 삶: 오랫동안 해양 생태계에서 마이너 플레이어로 여겨졌던 해파리는 사실 해양 먹이 그물의 중요한 부분이다." 자연, 531(7595): 432-435. 도이:10.1038/531432a
- ^ 카르도나, L, 드 케베도, I.A., 보렐, A., A. (2012) "지중해의 정점 포식자들에 의한 젤라틴성 플랑크톤의 대량 소비" PLOS ONE, 7(3): e31329. doi:10.1371/journal.pone.0031329
- ^ 2016년 4월 7일, 해양식품 웹 밑바닥의 작은 포리지 물고기 보호 전국 공영 라디오 방송
- ^ Roman, J. & McCarthy, J.J. (2010). "The Whale Pump: Marine Mammals Enhance Primary Productivity in a Coastal Basin". PLOS ONE. 5 (10): e13255. Bibcode:2010PLoSO...513255R. doi:10.1371/journal.pone.0013255. PMC 2952594. PMID 20949007. e13255.CS1 maint: 작성자 매개변수 사용(링크)
- ^ Brown, Joshua E. (12 October 2010). "Whale poop pumps up ocean health". Science Daily. Retrieved 18 August 2014.
- ^ Raina, J.B. (2018) "소규모에서의 생명 수생". mSystems, 3(2): e00150-17. doi:10.1128/mSystems.00150-17.
 자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다.
자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다. - ^ Marco, D, ed. (2011). Metagenomics: Current Innovations and Future Trends. Caister Academic Press. ISBN 978-1-904455-87-5.
- ^ a b c 로슬린, T.와 마자네바, S. (2016) "식품 웹 건설에서 DNA 바코드의 사용 - 지구와 수생 생태학자들이 단결한다!" 게놈, 59(9): 603–628. doi:10.1139/gen-2015-0229.
- ^ Wilhelm, Steven W.; Suttle, Curtis A. (1999). "Viruses and Nutrient Cycles in the Sea". BioScience. 49 (10): 781–788. doi:10.2307/1313569. ISSN 1525-3244. JSTOR 1313569.
- ^ 와인바우어, 마르쿠스 G 등 "바이러스 용해와 양성자 방목의 시너지 및 적대적 효과가 박테리아 바이오매스, 생산 및 다양성에 미치는 영향" 환경미생물학 9.3(2007) : 777-788.
- ^ 로빈슨, 캐롤, 그리고 나가파 라마야. "미생물 이질적 대사율은 미생물 탄소 펌프에 제약을 가한다." 미국과학진흥협회, 2011.
- ^ Van den Meersche, K, Middelburg, J.J., Soetaert, K, Van Rizswijk, P, Boschker, H.T. 및 Heip, C.H. (2004) "실험 꽃피는 동안 탄소 질소 결합과 녹갈균 상호작용: 13C 추적기 실험 모델링" Limnology and Oceanography, 49(3): 862–878. doi:10.4319/lo. 2004.49.3.0862.
- ^ 케스 L, 지우어 JK(2018) "해양 기후 변화에 대한 피토플랑크톤 반응 – 소개" Jungblut S, Liebich V, Bode M. (Eds) YOUMESTAR 8–Oceans Through Borders: 서로에게서 배우는 55-72페이지, 스프링어. 도이:10.1007/978-3-19-319-93284-2_5. 자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다.
- ^ 하인리히스, M.E., 모리, C., L.(2020) "수생생물들과 그들의 화학적 환경 사이의 복합적 상호작용" "다양한 관점에서 설명" 인: 유마레스 9-대양: 우리의 연구, 우리의 미래 279-297페이지. 스프링거. 도이:10.1007/978-3-030-20389-4_15.
- ^ M.G. (2019) "비바 라비다비루스! 거대한 DNA 바이러스를 기생시키는 바이로피지의 다섯 가지 특징"이라고 말했다. PLoS 병원균, 15(3) doi:10.1371/1998.ppat.1007592. 자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다.
- ^ Fischer, M. G.; Allen, M. J.; Wilson, W. H.; Suttle, C. A. (2010). "Giant virus with a remarkable complement of genes infects marine zooplankton" (PDF). Proceedings of the National Academy of Sciences. 107 (45): 19508–19513. Bibcode:2010PNAS..10719508F. doi:10.1073/pnas.1007615107. PMC 2984142. PMID 20974979.
- ^ Matthias G. Fischer; Michael J. Allen; William H. Wilson; Curtis A. Suttle (2010). "Giant virus with a remarkable complement of genes infects marine zooplankton" (PDF). Proceedings of the National Academy of Sciences. 107 (45): 19508–19513. Bibcode:2010PNAS..10719508F. doi:10.1073/pnas.1007615107. PMC 2984142. PMID 20974979.
- ^ Massana, Ramon; Javier Del Campo; Christian Dinter; Ruben Sommaruga (2007). "Crash of a population of the marine heterotrophic flagellate Cafeteria roenbergensis by viral infection". Environmental Microbiology. 9 (11): 2660–2669. doi:10.1111/j.1462-2920.2007.01378.x. PMID 17922751. S2CID 30191542.
- ^ Fischer MG, Suttle CA (April 2011). "A virophage at the origin of large DNA transposons". Science. 332 (6026): 231–4. Bibcode:2011Sci...332..231F. doi:10.1126/science.1199412. PMID 21385722. S2CID 206530677.
- ^ Fischer MG, Hackl (December 2016). "Host genome integration and giant virus-induced reactivation of the virophage mavirus". Nature. 540 (7632): 288–91. Bibcode:2016Natur.540..288F. doi:10.1038/nature20593. PMID 27929021. S2CID 4458402.
- ^ 가가미, M, 미키, T, 타키모토, G. (2014) "마이컬롭: 수생식용 거미줄 속의 치트리스드" 미생물학 분야 프론티어, 5: 166. doi:10.3389/fmicb.2014.00166. 자료는 이 출처에서 복사되었으며, Creative Commons Attribution 3.0 International License에 따라 이용할 수 있다.
- ^ Amend, A., Burgaud, G., Cunliffe, M., Edgcomb, V.P., Ettinger, C.L., Gutiérrez, M.H., Heitman, J., Hom, E.F., Ianiri, G., Jones, A.C. and Kagami, M. (2019) "Fungi in the marine environment: Open questions and unsolved problems". MBio, 10(2): e01189-18. doi:10.1128/mBio.01189-18. 자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다.
- ^ 구티에레스 MH, 자라 AM, 판토자 S(2016) "풍구 기생충은 칠레 중부 험볼트 현 시스템의 상류 생태계에 있는 해양 규조류를 감염시킨다." Environment Microbiol, 18(5): 1646–1653. doi:10.111/1462-2920.13257.
- ^ a b c d e 미델 뷔르흐, 잭은 J.(2019년)."그 반환 유기에서 무기 탄소에".해양 탄소 Biogeochemistry.지구 시스템 과학에 SpringerBriefs.를 대신하여 서명함. 37–56. doi:10.1007/978-3-030-10822-9_3.아이 에스비엔 978-3-030-10821-2.S2CID 104330175.재료는 창조적 공용 귀인 4.0국제 라이센스 하에 가능하다 이 원본에서 복사되었다.
- ^ Legendre L, Rassoulzadegan F(1995) "해양에서의 플랑크톤과 영양소 역학". 오필리아, 41:153–172.
- ^ 페이스 ML, 콜 JJ, 카펜터 SR, 키첼 JF(1999) "다양한 생태계에서 드러난 트로피컬 폭포" 트렌드 에콜 에볼루션, 14: 483–488.
- ^ 아잠 F, 펜첼 T, 필드 JG, 그레이 JS, 마이어 레이일 LA, 팅스타드 F(1983) "바다에서 물기둥 미생물의 생태학적 역할" 마에콜-프로그 세르, 10: 257–263.
- ^ Anderson TR과 LeB Williams PJ(1998) "영어 채널의 스테이션 E1에서 용해된 유기탄소의 계절적 사이클을 모델링. 에스타 코스트 쉘프 Sci, 46: 93–109.
- ^ Van den Meersche K, Middelburg JJ, Soetaert K, Van Rizsik P, Boschker HTS, Heip CHR(2004) "실험 꽃 피울 때의 탄소-질소 결합과 녹조-박테리아 상호작용: 13C 추적자 실험을 모델링" 림놀 오셔노그르, 49: 862–878.
- ^ Suttle CA (2005) "바다 속의 바이러스" 자연, 437: 356–361.
- ^ 시겔이 만들어 낸 데이비드 A.;Buesseler, 켄 O;Behrenfeld, 마이클 J.;Benitez-Nelson, 클라우디아 R., 간수장님, 엠마뉴엘, 브레진스키,인 MarkA.;Burd, 아드리안, 칼슨, 크레이그 a.;d'Asaro, 에릭 A.,.Doney, 스콧 C;페리, 메리 j가, 스탠리, Rachel은 H.R.;스타인 버그, 데보라 K.(2016년)."예측의 수출과 운명은 글로벌 해양 네트워크 총생산:.그 EXPORTS 과학 Plan".해양 과학에 국경. 3.doi:10.3389/fmars.2016.00022.재료는 창조적 공용 귀인 4.0국제 라이센스 하에 가능하다 이 원본에서 복사되었다.
- ^ 왕, F, 우, Y, 첸, Z, 장, G, 장, J, 정, S, 카트너(Kattner, G.) "안정적인 동위원소와 지방산으로 삽화된 남중국해 중피성 어류의 친밀한 상호작용" 해양 과학 분야의 프론티어, 5: 522. doi:10.3389/fmars.2018.00522.
- ^ a b 톨레프슨, 제프 (2020년 2월 27일) 황혼지대로 진입: 과학자들은 바다의 신비로운 중간 네이처 뉴스에 뛰어들어. 도이:10.1038/d41586-020-00520-8.
- ^ a b C.A.의 조이, Haddock, S.H.와 B.H.의 로비슨(2017)은 "현장 먹이 관찰로 밝혀진 심층 펠러지 식품 거미줄 구조"라고 말했다. 왕립학회 B: 생물과학의 진행, 284년(1868년): 20172116. doi:10.1098/rspb.2017.216. 소재는 이 출처에서 복사되었으며, Creative Commons Atttribution 4.0 International License에 따라 사용할 수 있다.
- ^ 켈리, T.B., 데이비슨, P.C., 괴릭케, R., 랜드리, M.R., 오만, M.R. 스투켈, M.R. (2019) "중간성 식품 웹을 유지하기 위한 메소즈오플랑크톤 디엘 수직이동의 중요성" 해양 과학의 프런티어, 6: 508. doi:10.3389/fmars.2019.00508.
- ^ 더 가디언지는 2020년 5월 25일 심해에서의 기후 변화가 세기 중반까지 7배 더 빨라질 수 있다고 보도했다.
- ^ Brito-Morales, I., Schoeman, D.S., Molinos, J.G., Burrows, M.T., Klein, C.J., Arafeh-Dalmau, N., Kaschner, K., Garilao, C., Kesner-Reyes, K. and Richardson, A.J. (2020) "Climate velocity reveals increasing exposure of deep-ocean biodiversity to future warming". 자연 기후 변화, 페이지 1-6. doi:10.5281/zenodo.3596584.
- ^ Irigoien, X., Klevjer, T.A., Røstad, A., Martinez, U., Boyra, G., Acuña, J.L., Bode, A., Echevarria, F., Gonzalez-Gordillo, J.I., Hernandez-Leon, S. and Agusti, S. (2014) "Large mesopelagic fishes biomass and trophic efficiency in the open ocean". 자연 통신, 5: 3271. doi:10.1038/ncomms4271
- ^ 해양의 어류 바이오매스는 2014년 2월 7일 추정된 유레크알레트보다 10배 높다.
- ^ Choy, C.A., Wabnitz, C.C., Weijerman, M., Woodworth-Jefcoats, P.A. and Polovina, J.J. (2016) "Finding the way to the top: how the composition of oceanic mid-trophic micronekton groups determines apex predator biomass in the central North Pacific". 해양 생태 진행 시리즈, 549: 9–25. doi:10.3354/mps11680.
- ^ a b c d e 쿠라타, N, 벨라, K, 해밀턴, B, 시브지, M, 솔로비예프, A, 맷, S, 타르타르, A., 그리고 W. (2016) "대양의 거의 표면층에 있는 항산화 박테리아" 자연: 과학 보고서, 6(1): 1–8. doi:10.1038/srep19123. 자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다.
- ^ 슈티치, 브이, 조소비치, B, 마르첸코, E, 비하리, N., 크르시니치, F.(1981) "해양 식물성 플랑크톤에 의한 계면활성제 생산" 해양화학, 10(6): 505–520. doi:10.1016/0304-4203(81)90004-9.
- ^ Levin LA, Baco AR, Bowden DA, Colaco A, Cordes EE, Cunha MR, Demopoulos AWJ, Gobin J, Grupe BM, Le J, Metaxas A, Netburn AN, Rouse GW, Thurber AR, Tunnicliffe V, Van Dover CL, Vanreusel A and Watling L (2016). "열수 분출과 메탄 침출: 영향권에 대해 다시 생각해 보십시오." 앞. 3월 과학 3:72. 도이:10.3389/fmars.2016.00072
- ^ 포르테일, M, 올루, K, 두부아, SF, 에스코바르-브리오네스, E, 겔리나스, Y, 메노트, L., 사라진, J. (2016) "과이마스 분지 열수 분출구의 식품-웹 복잡성과 냉수 침투" PLOS ONE, 11(9): 페이지0162263. doi:10.1371/journal.pone.0162263.
- ^ 베르나르디노 AF, 레빈 LA, 서버 AR, 스미스 CR(2012년). "환기구, 침출물, 유기폭포의 침전물 매크로푸나의 비교구성, 다양성, 영양생태학" PLOS ONE, 7(4): e33515. pmid:22496753. doi:10.1371/journal.pone.0033515.
- ^ 포르테일 M, 올루 K, 에스코바르-브리오네스 E, 카프라이스 JC, 메노 L, 와엘레스 M, 외 (2015) "과이마스 분지 내 환기구와 침투 매크로파우날 공동체에 대한 비교 연구" 생물지질학. 12(18): 5455–79. doi:10.5194/bg-12-5455-2015.
- ^ Harris, P.T.; Macmillan-Lawler, M.; Rupp, J.; Baker, E.K. (2014). "Geomorphology of the oceans". Marine Geology. 352: 4–24. Bibcode:2014MGeol.352....4H. doi:10.1016/j.margeo.2014.01.011.
- ^ a b c X.L. Otero, De La Peena-Lastra, S, Pérez-Alberti, A, Ferreira, T.O., M.A. Huerta-Diaz(2018) "질소와 인의 순환에서 중요한 글로벌 동인으로서 새로운 식민지" 자연 통신, 9(1): 1–8. doi:10.1038/s41467-017-02446-8. 자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다.
- ^ Barrett, Kyle; Anderson, Wendy B.; Wait, D. Alexander; Grismer, L. Lee; Polis†, Gary A.; Rose†, Michael D. (2005). "Marine subsidies alter the diet and abundance of insular and coastal lizard populations". Oikos. 109: 145–153. doi:10.1111/j.0030-1299.2005.13728.x.
- ^ Polis, Gary A.; Hurd, Stephen D. (1996). "Linking Marine and Terrestrial Food Webs: Allochthonous Input from the Ocean Supports High Secondary Productivity on Small Islands and Coastal Land Communities". The American Naturalist. 147 (3): 396–423. doi:10.1086/285858. S2CID 84701185.
- ^ Gende, Scott M.; Edwards, Richard T.; Willson, Mary F.; Wipfli, Mark S. (2002). "Pacific Salmon in Aquatic and Terrestrial Ecosystems". BioScience. 52 (10): 917. doi:10.1641/0006-3568(2002)052[0917:PSIAAT]2.0.CO;2. ISSN 0006-3568.
- ^ a b Gagnon, Karine; Rothäusler, Eva; Syrjänen, Anneli; Yli-Renko, Maria; Jormalainen, Veijo (2013). "Seabird Guano Fertilizes Baltic Sea Littoral Food Webs". PLOS ONE. 8 (4): e61284. Bibcode:2013PLoSO...861284G. doi:10.1371/journal.pone.0061284. PMC 3623859. PMID 23593452.
- ^ Mizota, Chitoshi; Noborio, Kosuke; Mori, Yoshiaki (2012). "The Great Cormorant (Phalacrocorax carbo) colony as a "hot spot" of nitrous oxide (N2O) emission in central Japan". Atmospheric Environment. 57: 29–34. Bibcode:2012AtmEn..57...29M. doi:10.1016/j.atmosenv.2012.02.007.
- ^ Bird, Michael I.; Tait, Elaine; Wurster, Christopher M.; Furness, Robert W. (2008). "Stable carbon and nitrogen isotope analysis of avian uric acid". Rapid Communications in Mass Spectrometry. 22 (21): 3393–3400. Bibcode:2008RCMS...22.3393B. doi:10.1002/rcm.3739. PMID 18837063.
- ^ a b Caut, Stéphane; Angulo, Elena; Pisanu, Benoit; Ruffino, Lise; Faulquier, Lucie; Lorvelec, Olivier; Chapuis, Jean-Louis; Pascal, Michel; Vidal, Eric; Courchamp, Franck (2012). "Seabird Modulations of Isotopic Nitrogen on Islands". PLOS ONE. 7 (6): e39125. Bibcode:2012PLoSO...739125C. doi:10.1371/journal.pone.0039125. PMC 3377609. PMID 22723945.
- ^ Mulder, Christa P. H.; Anderson, Wendy B.; Towns, David R.; Bellingham, Peter J. (8 September 2011). Seabird Islands: Ecology, Invasion, and Restoration. ISBN 9780199735693.
- ^ McFadden, Tyler Neal; Kauffman, J. Boone; Bhomia, Rupesh K. (2016). "Effects of nesting waterbirds on nutrient levels in mangroves, Gulf of Fonseca, Honduras". Wetlands Ecology and Management. 24 (2): 217–229. doi:10.1007/s11273-016-9480-4. S2CID 6021420.
- ^ Zwolicki, Adrian; Zmudczyńska-Skarbek, Katarzyna Małgorzata; Iliszko, Lech; Stempniewicz, Lech (2013). "Guano deposition and nutrient enrichment in the vicinity of planktivorous and piscivorous seabird colonies in Spitsbergen". Polar Biology. 36 (3): 363–372. doi:10.1007/s00300-012-1265-5. S2CID 12110520.
- ^ Doughty, Christopher E.; Roman, Joe; Faurby, Søren; Wolf, Adam; Haque, Alifa; Bakker, Elisabeth S.; Malhi, Yadvinder; Dunning, John B.; Svenning, Jens-Christian (2016). "Global nutrient transport in a world of giants". Proceedings of the National Academy of Sciences. 113 (4): 868–873. Bibcode:2016PNAS..113..868D. doi:10.1073/pnas.1502549112. PMC 4743783. PMID 26504209.
- ^ a b Honig, Susanna E.; Mahoney, Brenna (2016). "Evidence of seabird guano enrichment on a coral reef in Oahu, Hawaii". Marine Biology. 163 (2). doi:10.1007/s00227-015-2808-4. S2CID 87850538.
- ^ Kolb, GS; Ekholm, J.; Hambäck, PA (2010). "Effects of seabird nesting colonies on algae and aquatic invertebrates in coastal waters". Marine Ecology Progress Series. 417: 287–300. Bibcode:2010MEPS..417..287K. doi:10.3354/meps08791.
- ^ Wainright, S. C.; Haney, J. C.; Kerr, C.; Golovkin, A. N.; Flint, M. V. (1998). "Utilization of nitrogen derived from seabird guano by terrestrial and marine plants at St. Paul, Pribilof Islands, Bering Sea, Alaska". Marine Biology. 131: 63–71. doi:10.1007/s002270050297. S2CID 83734364.
- ^ Staunton Smith, J.; Johnson, CR (1995). "Nutrient inputs from seabirds and humans on a populated coral cay". Marine Ecology Progress Series. 124: 189–200. Bibcode:1995MEPS..124..189S. doi:10.3354/meps124189.
- ^ a b c 로랭, 앤, Houlbrèque, 패니, Benzoni, 프란체스카. Barjon, 루시,.Tremblay-Boyer, 로라, Menkes, 크리스토프;Gillikin, 데이비드 P.;Payri, 클로드, 주르당, 모랭, Boussarie, 제르맹, Verheyden, Anouk, 비달, 에릭(2017년)."reef-building 산호에 대한 원격 태평양 섬들에서 바다새는 공급 질소".과학 보고서. 7(1):3721.Bibcode:2017NatSR...7.3721L. doi:10.1038/s41598-017-03781-y. 1.5473863.PMID 28623288.S2CID 6539261.재료는 창조적 공용 귀인 4.0국제 라이센스 하에 가능하다 이 원본에서 복사되었다.
- ^ Connell, Des W. (4 May 2018). Pollution in Tropical Aquatic Systems. ISBN 9781351092777.
- ^ Hatcher, Bruce Gordon (1990). "Coral reef primary productivity. A hierarchy of pattern and process". Trends in Ecology & Evolution. 5 (5): 149–155. doi:10.1016/0169-5347(90)90221-X. PMID 21232343.
- ^ Falkowski, Paul G.; Dubinsky, Zvy; Muscatine, Leonard; McCloskey, Lawrence (1993). "Population Control in Symbiotic Corals". BioScience. 43 (9): 606–611. doi:10.2307/1312147. JSTOR 1312147.
- ^ a b Marubini, F.; Davies, P. S. (1996). "Nitrate increases zooxanthellae population density and reduces skeletogenesis in corals". Marine Biology. 127 (2): 319–328. doi:10.1007/BF00942117. S2CID 85085823.
- ^ 무스카틴, L. (1990) "탄소와 산호초에서 에너지 유동에서 공생 해조류의 역할," 에코시스템 월드, 25: 75–87.
- ^ Ferrier, M. Drew (1991). "Net uptake of dissolved free amino acids by four scleractinian corals". Coral Reefs. 10 (4): 183–187. Bibcode:1991CorRe..10..183F. doi:10.1007/BF00336772. S2CID 25973061.
- ^ Furla, P.; Allemand, D.; Shick, J. M.; Ferrier-Pagès, C.; Richier, S.; Plantivaux, A.; Merle, P. L.; Tambutté, S. (2005). "The Symbiotic Anthozoan: A Physiological Chimera between Alga and Animal". Integrative and Comparative Biology. 45 (4): 595–604. doi:10.1093/icb/45.4.595. PMID 21676806.
- ^ Mills, Matthew M.; Lipschultz, Fredric; Sebens, Kenneth P. (2004). "Particulate matter ingestion and associated nitrogen uptake by four species of scleractinian corals". Coral Reefs. 23 (3): 311–323. doi:10.1007/s00338-004-0380-3. S2CID 13212636.
- ^ Mills, M. M.; Sebens, K. P. (2004). "Ingestion and assimilation of nitrogen from benthic sediments by three species of coral". Marine Biology. 145 (6): 1097–1106. doi:10.1007/s00227-004-1398-3. S2CID 84698653.
- ^ Houlbrèque, F.; Tambutté, E.; Richard, C.; Ferrier-Pagès, C. (2004). "Importance of a micro-diet for scleractinian corals". Marine Ecology Progress Series. 282: 151–160. Bibcode:2004MEPS..282..151H. doi:10.3354/meps282151.
- ^ Ferrier-Pagès, C.; Witting, J.; Tambutté, E.; Sebens, K. P. (2003). "Effect of natural zooplankton feeding on the tissue and skeletal growth of the scleractinian coral Stylophora pistillata". Coral Reefs. 22 (3): 229–240. doi:10.1007/s00338-003-0312-7. S2CID 44869188.
- ^ Marubini, Francesca; Thake, Brenda (1999). "Bicarbonate addition promotes coral growth". Limnology and Oceanography. 44 (3): 716–720. Bibcode:1999LimOc..44..716M. doi:10.4319/lo.1999.44.3.0716.
- ^ Ferrier-Pagès, C.; Leclercq, N.; Jaubert, J.; Pelegrí, SP (2000). "Enhancement of pico- and nanoplankton growth by coral exudates". Aquatic Microbial Ecology. 21: 203–209. doi:10.3354/ame021203.
- ^ Renegar, DA; Riegl, BM (2005). "Effect of nutrient enrichment and elevated CO2 partial pressure on growth rate of Atlantic scleractinian coral Acropora cervicornis". Marine Ecology Progress Series. 293: 69–76. Bibcode:2005MEPS..293...69R. doi:10.3354/meps293069.
- ^ 피터슨, J.K, 홀머, M, 테르만센, M., 하슬러, B.(2019) "이발관을 통한 영양 추출" In: Smaal A, Fererira J, Grant J, Petersen J, Strand ø. (eds) Marine Bivalves의 상품 및 서비스, 179–208페이지. 스프링거. 도이:10.1007/978-3-19-96776-9_10. ISBN 9783319967769
- ^ 헤이먼스, JJ, 콜, M, 리브루카토, S, 모리셋, L., 크리스텐슨, V. (2014) "해양식품 거미줄의 생태학적 지표에 나타난 세계적인 패턴: 모델링 접근법" PLOS ONE, 9(4). doi:10.1371/journal.pone.0095845.
- ^ 프라노비, F, 리브루카토, S, 라이체비치, S, 그란조토, A, 파스트레스, R., 지오바나르디, O. (2003) "베니스 석호의 기계조개 준설: 영양 질량 균형 모델로 평가된 생태계 효과" 해양생물학, 143(2): 393–403. doi:10.1007/s00227-003-1072-1.
- ^ 미국 지질조사국(USGS) "14장: 먹이 변화와 물새 서식지 변화" 그림 14.1.체서피크 만 생태계를 위한 미국 지질조사 과학의 합성 및 환경관리에 대한 시사점 USGS Circular 1316.
 이 글은 공개 도메인에 있는 이 출처의 텍스트를 통합한다..
이 글은 공개 도메인에 있는 이 출처의 텍스트를 통합한다.. - ^ Perry, M.C., Osenton, P.C., Wells-Berlin, A.M., and Kidwell, D.M., 2005, Food selection among Atlantic Coast sea ducks in relation to historic food habits, [abs.] in Perry, M.C., Second North American Sea Duck Conference, November 7–11, 2005, Annapolis, Maryland, Program and Abstracts, USGS Patuxent Wildlife Research Center, Maryland, 123 p. (p. 105).
- ^ 보우서(Bowser) A.K.Diamond.A.W., J.A.A.A.Addison(2013) "퍼핀에서 플랑크톤까지: 메인만 북부의 바닷새 먹이 사슬에 대한 DNA 기반 분석" PLOS ONE, 8(12): e83152. doi:10.1371/journal.pone.0083152
- ^ 레레이 M, 마이어 CP, 밀스 SC(2015) "산호 서식지 포식 물고기의 메타바코딩 식이 분석은 산호 상호주의자들이 고도로 분할된 일반주의 식단에 대한 작은 기여를 보여준다." PeerJ, 3: e1047. doi:10.7717/peerj.1047.
- ^ 콜, M, 슈미트, A, 로마누크, T, 로제, H.K. (2011년) "다른 공간적 규모와 인간의 영향에 걸친 해초 공동체의 음식 웹 구조" PLOS ONE, 6(7): e22591. doi:10.1371/journal.pone.0022591. 자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다.
- ^ Stat, M., Huggett, M.J., Bernasconi, R., DiBattista, J.D., Berry, T.E., Newman, S.J., Harvey, E.S. and Bunce, M. (2017) "Ecosystem biomonitoring with eDNA: metabarcoding across the tree of life in a tropical marine environment". 과학 보고서, 7(1): 1–11. doi:10.1038/s41598-017-12501-5.
- ^ 아처, 스테파니. K.;칸, 아만다 S.,.Thiess, 메리, 법, 로렌, Leys, 샐리는 P.;Johannessen, 소피아 C;Layman, 크레이그 a.;.버크, 릴리, 던햄, Anya(9월 24일 2020년)."재단의 종 Abundance 영향에 식품 웹 위상 유리 스펀지초에".해양 과학에 국경.둔 미디어 SA.7.doi:10.3389/fmars.2020.549478.ISSN 2296-7745.재료는 창조적 공용 귀인 4.0국제 라이센스 하에 가능하다 이 원본에서 복사되었다.
- ^ Lobo, Eduardo A.; Heinrich, Carla Giselda; Schuch, Marilia; Wetzel, Carlos Eduardo; Ector, Luc (2016), Necchi JR, Orlando (ed.), "Diatoms as Bioindicators in Rivers", River Algae, Springer International Publishing, pp. 245–271, doi:10.1007/978-3-319-31984-1_11, ISBN 9783319319834
- ^ Stevenson, R. Jan; Pan, Yangdong; van Dam, Herman (2010), Smol, John P.; Stoermer, Eugene F. (eds.), "Assessing environmental conditions in rivers and streams with diatoms", The Diatoms (2 ed.), Cambridge University Press, pp. 57–85, doi:10.1017/cbo9780511763175.005, ISBN 9780511763175
- ^ 매카시, J.J., 칸자니, O.F., 리어리, N.A., 도켄, D.J.와 화이트, K.S. (Eds.)(2001) 기후 변화 2001: 영향, 적응 및 취약성: 캠브리지 대학 출판부의 정부간 기후변화위원회 제3차 평가 보고서에 대한 작업 그룹 II의 기여. ISBN 9780521015004
- ^ Murphy, E. J.; Cavanagh, R. D.; Drinkwater, K. F.; Grant, S. M.; Heymans, J. J.; Hofmann, E. E.; Hunt, G. L.; Johnston, N. M. (2016). "Understanding the structure and functioning of polar pelagic ecosystems to predict the impacts of change". Proceedings of the Royal Society B: Biological Sciences. 283 (1844). doi:10.1098/rspb.2016.1646. PMC 5204148. PMID 27928038.
- ^ 기후가 생태계에 미치는 영향: 식품 웹 중단EPA. 2020년 2월 11일에 접속. 이 글은 공개 도메인에 있는 이 출처의 텍스트를 통합한다..
- ^ "A 'regime shift' is happening in the Arctic Ocean, scientists say". phys.org. Retrieved 16 August 2020.
- ^ Lewis, K. M.; Dijken, G. L. van; Arrigo, K. R. (10 July 2020). "Changes in phytoplankton concentration now drive increased Arctic Ocean primary production". Science. 369 (6500): 198–202. doi:10.1126/science.aay8380. ISSN 0036-8075. PMID 32647002. S2CID 220433818. Retrieved 16 August 2020.
- ^ 스토커 D.K.와 라브렌티예프 P.J. (2018) "극해의 미생물학적 플랑크톤: 범북극적 검토" 해양과학 분야의 프론티어, 5:292. doi:10.3389/fmars.2018.00292
- ^ Kilias, 에스텔 S.;Junges, 레안 드루;Šupraha, 루카, 레오나드, 가이, Metfies, 카트자, 리처즈, 토마스 A. 토마(2020년)."규조류와Chytrid 진균류 분배와 동시 발생. 바다 얼음을 녹과 북극해에 관련이 있".통신 생물학. 3(1):183.doi:10.1038/s42003-020-0891-7. 1.7174370.PMID 32317738.S2CID 216033140.재료는 창조적 공용 귀인 4.0국제 라이센스 하에 가능하다 이 원본에서 복사되었다.
- ^ Bender, Sara J.; Moran, Dawn M.; McIlvin, Matthew R.; Zheng, Hong; McCrow, John P.; Badger, Jonathan; Ditullio, Giacomo R.; Allen, Andrew E.; Saito, Mak A. (2018). "Colony formation in Phaeocystis antarctica: Connecting molecular mechanisms with iron biogeochemistry". Biogeosciences. 15 (16): 4923–4942. doi:10.5194/bg-15-4923-2018.
- ^ Pinkernell, Stefan; Beszteri, Bánk (2014). "Potential effects of climate change on the distribution range of the main silicate sinker of the Southern Ocean". Ecology and Evolution. 4 (16): 3147–3161. doi:10.1002/ece3.1138. PMC 4222203. PMID 25473469.
- ^ 캐번, E.L.;Belcher A., Atkinson은 A.;힐, S.L.;가와구치, S.;매코맥 S., 마이어, B, 니콜, S.;Ratnarajah, L.;슈미트, K, 스타인 버그, D.K.;Tarling, G.A.;보이드, P.W.(2019년)."남극 크릴 새우의 생물 지구 화학의 싸이클에서 찾하".네이쳐 커뮤니케이션. 10(1):4742.doi:10.1038/s41467-019-12668-7. 1.6800442.PMID 31628346..재료는 창조적 공용 귀인 4.0국제 라이센스 하에 가능하다 이 원본에서 복사되었다.
- ^ a b Cordone, Georgina; Marina, Tomás I.; Salinas, Vanesa; Doyle, Santiago R.; Saravia, Leonardo A.; Momo, Fernando R. (2018). "Effects of macroalgae loss in an Antarctic marine food web: Applying extinction thresholds to food web studies". PeerJ. 6: e5531. doi:10.7717/peerj.5531. PMC 6139014. PMID 30225167.
- ^ Marina, Tomás I.; Salinas, Vanesa; Cordone, Georgina; Campana, Gabriela; Moreira, Eugenia; Deregibus, Dolores; Torre, Luciana; Sahade, Ricardo; Tatián, Marcos; Barrera Oro, Esteban; De Troch, Marleen; Doyle, Santiago; Quartino, María Liliana; Saravia, Leonardo A.; Momo, Fernando R. (2018). "The Food Web of Potter Cove (Antarctica): Complexity, structure and function". Estuarine, Coastal and Shelf Science. 200: 141–151. doi:10.1016/j.ecss.2017.10.015.
- ^ Koh, Eileen Y.; Martin, Andrew R.; McMinn, Andrew; Ryan, Ken G. (2012). "Recent Advances and Future Perspectives in Microbial Phototrophy in Antarctic Sea Ice". Biology. 1 (3): 542–556. doi:10.3390/biology1030542. PMC 4009807. PMID 24832507.
- ^
- Azam, F.; Fenchel, T.; Field, J. G.; Gray, J. S.; Meyer-Reil, L. A.; Thingstad, F. (1983). "The Ecological Role of Water-Column Microbes in the Sea". Marine Ecology Progress Series. 10 (3): 257–263. doi:10.3354/meps010257. JSTOR 24814647.
- Fenchel, Tom (2008). "The microbial loop – 25 years later". Journal of Experimental Marine Biology and Ecology. 366 (1–2): 99–103. doi:10.1016/j.jembe.2008.07.013.
- ^ 라미, 티, 코에닉스, C, 홀브룩, S.J., 밀러, R.J., 스타이어, 에이씨, 리드, D.C.(2020) "창작종들은 거대한 다시마 숲에서 다양성을 증가시켜 지역사회 안정을 도모한다." 생태학, e02987. doi:10.1002/ecy.2987.
- ^ 1972년 P. K. 데이튼. 지역사회의 회복력과 남극 맥머도 사운드의 벤토에 대한 잠재적 영향에 대한 이해를 돕기 위해. <보존 문제에 관한 콜로키움 절차>, <캔자스 로렌스>에서 81-96쪽.
- ^ 거대한 다시마는 2020년 2월 4일 캘리포니아 남부의 해양 생태계에 강한 토대인 National Science Foundation을 제공한다.
- ^ Angelini, Christine; Altieri, Andrew H.; et al. (October 2011). "Interactions among Foundation Species and Their Consequences for Community Organization, Biodiversity, and Conservation". BioScience. 61 (10): 782–789. doi:10.1525/bio.2011.61.10.8.
- ^ Ellison, Aaron M.; Bank, Michael S.; et al. (November 2005). "Loss of foundation species: consequences for the structure and dynamics of forested ecosystems". Frontiers in Ecology and the Environment. 3 (9): 479–486. doi:10.1890/1540-9295(2005)003[0479:LOFSCF]2.0.CO;2.
- ^ Paine, R. T. (1969). "A Note on Trophic Complexity and Community Stability". The American Naturalist. 103 (929): 91–93. doi:10.1086/282586. JSTOR 2459472. S2CID 83780992.
- ^ "Keystone Species Hypothesis". University of Washington. Archived from the original on 10 January 2011. Retrieved 3 February 2011.
- ^ Paine, R. T. (1966). "Food web complexity and species diversity". American Naturalist. 100 (910): 65–75. doi:10.1086/282400. JSTOR 2459379. S2CID 85265656.
- ^ Paine, R. T. (1995). "A Conversation on Refining the Concept of Keystone Species". Conservation Biology. 9 (4): 962–964. doi:10.1046/j.1523-1739.1995.09040962.x.
- ^ Davic, Robert D. (2003). "Linking Keystone Species and Functional Groups: A New Operational Definition of the Keystone Species Concept". Conservation Ecology. Retrieved 3 February 2011.
- ^ Szpak, Paul; Orchard, Trevor J.; Salomon, Anne K.; Gröcke, Darren R. (2013). "Regional ecological variability and impact of the maritime fur trade on nearshore ecosystems in southern Haida Gwaii (British Columbia, Canada): evidence from stable isotope analysis of rockfish (Sebastes spp.) bone collagen". Archaeological and Anthropological Sciences. 5 (2): 159–182. doi:10.1007/s12520-013-0122-y. S2CID 84866250.
- ^ Cohn, J. P. (1998). "Understanding Sea Otters". BioScience. 48 (3): 151–155. doi:10.2307/1313259. JSTOR 1313259.
- ^ a b c d e f Endrédi, Anett, Patonai, Katalin, Podani, 헝가리, Libralato, 시몬. Jordán, 페렌츠(2021년)."누구는 어디에 있해양 식품 웨브에?네트워크 Positions"의Trait-Based 분석.해양 과학에 국경. 8.doi:10.3389/fmars.2021.636042.재료는 창조적 공용 귀인 4.0국제 라이센스 하에 가능하다 이 원본에서 복사되었다.
- ^ Lawton, John H. (1994). "What do Species do in Ecosystems?". Oikos. 71 (3): 367–374. doi:10.2307/3545824. JSTOR 3545824.
- ^ Luczkovich, Joseph J.; Borgatti, Stephen P.; Johnson, Jeffrey C.; Everett, Martin G. (2003). "Defining and Measuring Trophic Role Similarity in Food Webs Using Regular Equivalence". Journal of Theoretical Biology. 220 (3): 303–321. Bibcode:2003JThBi.220..303L. doi:10.1006/jtbi.2003.3147. PMID 12468282.
- ^ Paine, Robert T. (1966). "Food Web Complexity and Species Diversity". The American Naturalist. 100 (910): 65–75. doi:10.1086/282400. S2CID 38675566.
- ^ Paine, Robert T. (1969). "The Pisaster-Tegula Interaction: Prey Patches, Predator Food Preference, and Intertidal Community Structure". Ecology. 50 (6): 950–961. doi:10.2307/1936888. JSTOR 1936888.
- ^ Mills, L. Scott; Soulé, Michael E.; Doak, Daniel F. (1993). "The Keystone-Species Concept in Ecology and Conservation". BioScience. 43 (4): 219–224. doi:10.2307/1312122. JSTOR 1312122.
- ^ Power, Mary E.; Tilman, David; Estes, James A.; Menge, Bruce A.; Bond, William J.; Mills, L. Scott; Daily, Gretchen; Castilla, Juan Carlos; Lubchenco, Jane; Paine, Robert T. (1996). "Challenges in the Quest for Keystones". BioScience. 46 (8): 609–620. doi:10.2307/1312990. JSTOR 1312990.
- ^ Bond, W. J. (1994). "Keystone Species". Biodiversity and Ecosystem Function. pp. 237–253. doi:10.1007/978-3-642-58001-7_11. ISBN 978-3-540-58103-1.
- ^ Marcogliese, David J.; Cone, David K. (1997). "Food webs: A plea for parasites". Trends in Ecology & Evolution. 12 (8): 320–325. doi:10.1016/s0169-5347(97)01080-x. PMID 21238094.
- ^ Pinnegar, John.K; Polunin, Nicholas V.C (2004). "Predicting indirect effects of fishing in Mediterranean rocky littoral communities using a dynamic simulation model". Ecological Modelling. 172 (2–4): 249–267. doi:10.1016/J.ECOLMODEL.2003.09.010.
- ^ Jordán, Ferenc; Benedek, Zsófia; Podani, János (2007). "Quantifying positional importance in food webs: A comparison of centrality indices". Ecological Modelling. 205 (1–2): 270–275. doi:10.1016/j.ecolmodel.2007.02.032.
- ^ Endrédi, Anett; Senánszky, Vera; Libralato, Simone; Jordán, Ferenc (2018). "Food web dynamics in trophic hierarchies". Ecological Modelling. 368: 94–103. doi:10.1016/j.ecolmodel.2017.11.015.
- ^ Scotti, M.; Jordán, F. (2010). "Relationships between centrality indices and trophic levels in food webs". Community Ecology. 11: 59–67. doi:10.1556/comec.11.2010.1.9.
- ^ Baranyi, Gabriella; Saura, Santiago; Podani, János; Jordán, Ferenc (2011). "Contribution of habitat patches to network connectivity: Redundancy and uniqueness of topological indices". Ecological Indicators. 11 (5): 1301–1310. doi:10.1016/j.ecolind.2011.02.003.
- ^ Pereira, Juliana; Saura, Santiago; Jordán, Ferenc (2017). "Single‐node vs. Multi‐node centrality in landscape graph analysis: Key habitat patches and their protection for 20 bird species in NE Spain". Methods in Ecology and Evolution. 8 (11): 1458–1467. doi:10.1111/2041-210X.12783.
- ^ a b c d 밀레트, N.C., 그로스, J., 존슨, W.M., 융블루스, M.J., 수터, E.A. (2018) "평범한 시야에 숨겨져 있다. 해양 플랑크톤에서 암호화된 상호작용의 중요성" Limnology and Oceanography Letters, 3(4): 341–356. doi:10.1002/lol2.10084. 자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다.
- ^ a b c Luypaert, T, Hagan, J.G., McCarthy, M.L., Poti, "2020" "인류세 내 해양 생물다양성 현황" 인: 유마레스 9-대양: 우리의 연구, 우리의 미래, 57-82, 스프링거. 도이:10.1007/978-3-030-20389-4_4.
- ^ 에스테스 JA, 팅커 MT, 윌리엄스 TM 외 연구진(1998) "해양과 근해 생태계를 잇는 해달에 대한 킬러 고래 포식" 과학, 282: 473–476. doi:10.1126/과학.282.5388.473.
- ^ Post, D. M. (1993). "The long and short of food-chain length". Trends in Ecology and Evolution. 17 (6): 269–277. doi:10.1016/S0169-5347(02)02455-2.
- ^ Jerry Bobrow, Ph.D.; Stephen Fisher (2009). CliffsNotes CSET: Multiple Subjects (2nd ed.). John Wiley and Sons. p. 283. ISBN 978-0-470-45546-3.
- ^ 엘튼 CS (1927) 동물 생태학. 2001년 재발행. 시카고 대학교 출판부
- ^ Paine RT (1966). "Food web complexity and species diversity". The American Naturalist. 100 (910): 65–75. doi:10.1086/282400. S2CID 85265656.
- ^ a b 5월 RM(2001) 모델 생태계의 안정성과 복잡성 프린스턴 대학교 출판부에서 1973년판을 새로운 서문으로 재인쇄했다. ISBN 978-0-691-08861-7
- ^ a b 1982년 판을 새로운 서문으로 재인쇄한 Pimm SL (2002) Chicago Press의 Food Webs University of Chicago Press. ISBN 978-0-226-66832-1
- ^ Kotta, J.; Wernberg, T.; Jänes, H.; Kotta, I.; Nurkse, K.; Pärnoja, M.; Orav-Kotta, H. (2018). "Novel crab predator causes marine ecosystem regime shift". Scientific Reports. 8 (1): 4956. Bibcode:2018NatSR...8.4956K. doi:10.1038/s41598-018-23282-w. PMC 5897427. PMID 29651152.
- ^ Megrey, Bernard and Werner, Francisco. "Evaluating the Role of Topdown vs. Bottom-up Ecosystem Regulation from a Modeling Perspective" (PDF).CS1 maint: 여러 이름: 작성자 목록(링크)
- ^ Frank, K. T.; Petrie, B.; Choi, J. S.; Leggett, W. C. (2005). "Trophic Cascades in a Formerly Cod-Dominated Ecosystem". Science. 308 (5728): 1621–1623. Bibcode:2005Sci...308.1621F. doi:10.1126/science.1113075. ISSN 0036-8075. PMID 15947186. S2CID 45088691.
- ^ Matsuzaki, Shin-Ichiro S.; Suzuki, Kenta; Kadoya, Taku; Nakagawa, Megumi; Takamura, Noriko (2018). "Bottom-up linkages between primary production, zooplankton, and fish in a shallow, hypereutrophic lake". Ecology. 99 (9): 2025–2036. doi:10.1002/ecy.2414. PMID 29884987.
- ^ Lynam, Christopher Philip; Llope, Marcos; Möllmann, Christian; Helaouët, Pierre; Bayliss-Brown, Georgia Anne; Stenseth, Nils C. (February 2017). "Trophic and environmental control in the North Sea". Proceedings of the National Academy of Sciences. 114 (8): 1952–1957. doi:10.1073/pnas.1621037114. PMC 5338359. PMID 28167770.
- ^ "Oldlist". Rocky Mountain Tree Ring Research. Retrieved 8 January 2013.
- ^ Wetzel, Robert (2001). Limnology: Lake and river ecosystems. San Diego: Academic Press. ISBN 9780127447605.
- ^ a b Field, C. B.; Behrenfeld, M. J.; Randerson, J. T.; Falkowski, P. (1998). "Primary Production of the Biosphere: Integrating Terrestrial and Oceanic Components". Science. 281 (5374): 237–240. Bibcode:1998Sci...281..237F. doi:10.1126/science.281.5374.237. PMID 9657713.
- ^ 바온, Y.M., 필립스, R., 마일로, R. (2018) "지구상의 바이오매스 분포" 국립과학원 절차, 115(25): 6506–6511. doi:10.1073/pnas.171184211.
- ^ Spellman, Frank R. (2008). The Science of Water: Concepts and Applications. CRC Press. p. 167. ISBN 978-1-4200-5544-3.
- ^ Odum, E. P.; Barrett, G. W. (2005). Fundamentals of Ecology (5th ed.). Brooks/Cole, a part of Cengage Learning. ISBN 978-0-534-42066-6. Archived from the original on 20 August 2011.
- ^ Wang, H.; Morrison, W.; Singh, A.; Weiss, H. (2009). "Modeling inverted biomass pyramids and refuges in ecosystems" (PDF). Ecological Modelling. 220 (11): 1376–1382. doi:10.1016/j.ecolmodel.2009.03.005. Archived from the original (PDF) on 7 October 2011.
- ^ Smith, S. V. (1981). "Marine Macrophytes as a Global Carbon Sink". Science. 211 (4484): 838–840. Bibcode:1981Sci...211..838S. doi:10.1126/science.211.4484.838. PMID 17740399.
- ^ Maureaud, A, Gascuel, D, Colléter, M, Palomares, M.L., Du Pontavice, H, Pauly, D., Cheung, W.W. "2017년" "해양식량 웹의 영양 기능의 세계적 변화" PLOS ONE, 12(8): e0182826. doi:10.1371/journal.pone.0182826
- ^ "Effects of Ocean Acidification on Marine Species & Ecosystems". Report. OCEANA. Retrieved 13 October 2013.
- ^ "Comprehensive study of Arctic Ocean acidification". Study. CICERO. Archived from the original on 10 December 2013. Retrieved 14 November 2013.
- ^ Lischka, S.; Büdenbender J.; Boxhammer T.; Riebesell U. (15 April 2011). "Impact of ocean acidification and elevated temperatures on early juveniles of the polar shelled pteropod Limacina helicina : mortality, shell degradation, and shell growth" (PDF). Report. Biogeosciences. pp. 919–932. Retrieved 14 November 2013.
- ^ "Antarctic marine wildlife is under threat, study finds". BBC Nature. Retrieved 13 October 2013.
- ^ 울라, H, 나겔커켄, I, 골든버그, S.U.와 포덤, 디에이 (2018) "기후 변화는 변화하는 영양 흐름과 시아노박테리아 증식을 통해 해양 식품 웹 붕괴를 몰고 올 수 있다." PLoS 생물학, 16(1): e200346. doi:10.1371/journal.pbio.2346
- ^ a b 기후변화로 인해 해양식품 웹 사이트 사이언스데일리 붕괴 사고 발생 2018년 1월 9일.
- ^ IUCN (2018) IUCN 멸종 위기 종 목록: 버전 2018-1


