플라워
Flower
꽃 또는 꽃이라고도 하는 꽃은 꽃이 피는 식물(Angiospermae과의 식물)에서 볼 수 있는 생식 구조입니다. 꽃은 발육하는 꽃을 감싸고 보호하는 분절, 꽃가루 매개자를 유인하는 꽃잎, 그리고 꽃이 피는 식물에서 배우자를 생산하는 배우자를 생산하는 생식 기관의 조합으로 이루어져 있습니다. 정자를 생산하는 수컷 배우자식물은 화분에서 생산되는 꽃가루 알갱이 안에 둘러싸여 있습니다. 암컷 배우자 식물은 카르펠에서 생성된 난자 내에 포함되어 있습니다.
대부분의 꽃이 피는 식물은 벌, 나방, 나비와 같은 동물에 의존하여 다른 꽃들 사이로 꽃가루를 옮기며, 밝은 색, 눈에 띄는 꽃잎, 매력적인 향기, 그리고 꽃가루 매개자들의 음식 공급원인 꿀의 생산을 포함한 다양한 전략에 의해 이러한 꽃가루 매개자들을 유인하도록 진화해 왔습니다.[1] 이러한 방식으로, 많은 꽃이 피는 식물들은 꽃가루 매개자들과 함께 진화하여 서로에게 제공하는 서비스(식물의 경우, 번식의 수단)에 상호 의존하고, 꽃가루 매개자의 경우, 식량의 공급원.[2]
꽃의 꽃받침에서 나온 꽃가루가 암술머리에 쌓이면 이것을 수분작용이라고 합니다. 어떤 꽃들은 같은 식물의 다른 꽃의 꽃가루를 사용하여 종자를 생산하는 자가 수분을 할 수 있지만, 다른 꽃들은 같은 종의 다른 꽃의 꽃에서 다른 꽃의 낙인으로 꽃가루가 옮겨질 때 자가 수분을 방지하고 교차 수분에 의존하는 메커니즘을 가지고 있습니다. 자가수분은 수술과 목관이 동시에 성숙하는 꽃에서 일어나 꽃가루가 꽃의 낙인에 착지할 수 있도록 배치됩니다. 이 수분은 수분 매개체에게 꿀과 꽃가루를 먹이로 제공하기 위해 식물의 투자를 필요로 하지 않습니다.[3] 어떤 꽃들은 수정 없이 디아스포라(parthenocarpy)를 생산합니다. 수정 후에 꽃의 난소는 씨앗을 함유한 과일로 발전합니다.
꽃은 예로부터 아름다움과 기분 좋은 향기로 사람들에게 높이 평가되어 왔으며, 또한 종교적, 의식적 또는 상징적인 대상, 또는 약과 음식의 공급원으로서 문화적 의미를 지니고 있습니다.
어원
꽃은 17세기에 갈라지기 전에 간 곡물과 식물의 생식 구조를 모두 지칭하는 중세 영국 밀가루에서 유래했습니다. 이것은 원래 이탈리아 꽃의 여신 플로라의 라틴어 이름에서 유래했습니다. 영어에서 꽃의 초기 단어는 blossom이었지만 지금은 과일 나무의 꽃만을 의미합니다.[4][5]
형태학

꽃의 형태,[6] 또는 그 형태와 구조는 꽃잎과 같은 비생식적인 구조로 구성된 식물 부분과 생식 또는 성적인 부분의 두 부분으로 나누어 생각할 수 있습니다. 전형적인 꽃은 짧은 줄기나 축의 끝에 붙어 있는 네 종류의 구조물로 이루어져 있는데, 이것을 리셉터클이라고 합니다. 이 각각의 부분이나 꽃 기관들은 소용돌이라고 불리는 나선형으로 배열되어 있습니다.[7] 꽃의 밑부분이나 가장 낮은 마디에서 시작하여 위쪽으로 작용하는 네 개의 주요 회오리는 꽃받침, 화관, 회향, 그리고 회향입니다. 꽃받침과 화관이 함께 꽃의 비생식 부분을 구성하는데, 경우에 따라서는 분화되지 않을 수도 있습니다. 이 경우, 그들은 테팔이라고 설명됩니다.[8]
페리안스
칼릭스
꽃받침이라고 통칭되는 꽃받침은 꽃의 가장 바깥쪽 소용돌이에 생기는 변형된 잎입니다. 그들은 넓은 기저부, 기공, 결절, 엽록소를 가지고 있다는 점에서 잎과 같습니다.[9] 꽃받침은 종종 왁스질이고 질기며, 꽃이 발달함에 따라 보호하기 위해 빨리 자랍니다.[9][10] 낙엽성일 수 있지만 과일 분산을 돕기 위해 더 일반적으로 자랍니다. 만약 꽃받침이 함께 융합된다면 그것은 가모세팔루스라고 불립니다.[9]
코롤라
꽃잎은 화관과 함께 거의 또는 완전히 섬유질이 없는 잎과 같은 구조로 근막의 가장 안쪽 소용돌이를 형성합니다. 그들은 종종 섬세하고 얇으며 수분을 촉진하기 위해 대개 색상, 모양 또는 향기가 납니다.[11] 잎과 모양은 비슷하지만 서로 거의 동시에 형성된다는 점에서 수술과 비교할 수 있지만 이후 성장이 지연됩니다. 화관이 함께 융합되면 심페틸러스라고 불립니다.[12]
생식

안드로에슘
안드로에시움, 즉 수술은 꽃가루를 생산하는 남성 부위의 전체입니다. 수술은 일반적으로 두 개의 맹장에 배열된 4개의 꽃가루 주머니로 구성된 또 다른 주머니로 구성되며 필라멘트 또는 줄기에 연결됩니다. 또 다른 것은 감수분열을 거친 후에 꽃가루가 되는 수생식물인 미세포자세포를 포함하고 있습니다. 꽃 기관 중에서 가장 넓은 차이를 보이지만, 안드로에시움은 보통 한 개의 회오리에만 국한되며, 드물게 두 개의 회오리에만 국한됩니다. 수술은 수, 크기, 모양, 방향, 꽃과의 연결점 등에서 다양합니다.[11][12]
일반적으로 수술은 한 가지 종류만 있지만, 꽃이 두 가지 종류를 가지고 있는 식물 종들이 있습니다: "정상적인" 것과 꽃가루 매개자를 유인하기 위한 멸균 꽃가루를 생산하는 꽃이 있는 것.[13]
자이에시움
지네시움 또는 카르펠은 가장 안쪽의 소용돌이에서 발견되는 꽃의 암컷 부분입니다. 각각의 카르펠은 꽃가루를 받는 낙인, 줄기 역할을 하는 스타일, 그리고 난자를 포함하는 난소로 구성되어 있습니다. 카르펠은 한 바퀴에서 여러 바퀴로 발생할 수 있으며, 함께 융합될 때 흔히 암술로 설명됩니다. 난소 안에서 난포는 후니쿨리(funiculi)라고 불리는 구조에 의해 태반에 부착됩니다.[14][15]
변화
이러한 배열은 "전형적인" 것으로 여겨지지만, 식물 종들은 꽃의 구조에 있어서 매우 다양한 변화를 보여줍니다.[16] 꽃의 네 가지 주요 부분은 일반적으로 기능이 아닌 용기의 위치에 의해 정의됩니다. 많은 꽃들은 일부 부분이 부족하거나 다른 기능으로 수정되거나 일반적으로 다른 부분처럼 보일 수 있습니다.[17] 풀과 같은 일부 과에서는 꽃잎이 크게 줄어듭니다. 많은 종에서 꽃받침은 알록달록하고 꽃잎처럼 생겼습니다. 다른 꽃들은 꽃잎 모양의 변형된 수술을 가지고 있습니다; 모란과 장미의 이중 꽃들은 대부분 꽃잎 모양의 수술입니다.[18]
많은 꽃들이 대칭을 가지고 있습니다. 어느 한 점에서 중심축을 통해 근일점을 이등분하여 대칭적인 반이 만들어지면 꽃은 동형 또는 규칙적이라고 합니다. 이것은 방사상 대칭의 한 예입니다. 꽃을 이등분하여 대칭적인 반을 만드는 한 줄만 만들어낼 때, 꽃은 불규칙하거나 접합형이라고 합니다. 드물게 대칭이 전혀 없는 경우를 비대칭이라고 합니다.[19][20]
꽃은 식물의 기저부에 직접 부착될 수 있습니다(가해, 지지 줄기 또는 줄기가 매우 감소하거나 부재합니다).[21] 꽃을 따르는 줄기나 줄기, 또는 꽃의 꽃이 피는 것을 꽃자루라고 부릅니다. 한 개의 꽃자루가 두 개 이상의 꽃을 지탱하는 경우, 각 꽃과 주축을 연결하는 줄기를 꽃자루라고 합니다.[22] 꽃줄기의 꼭짓점은 말단부종을 형성하는데, 이것을 토러스 또는 리셉터클이라고 합니다.[20]
대부분의 종에서 개별 꽃은 암술과 수술을 모두 가지고 있습니다. 식물학자들은 이 꽃들이 완벽하거나 양성 또는 자웅동체라고 설명합니다. 어떤 종류의 식물에서는 꽃이 불완전하거나 단성입니다: 수컷(주걱) 또는 암컷(주걱) 부분만 있습니다. 만약 같은 식물에 암수의 단성화가 나타난다면, 그 종은 단성화라고 불립니다.[23] 그러나 개별 식물이 암컷 또는 수컷이면 그 종을 다이오시어라고 부릅니다. 많은 꽃들은 수분 매개체를 유인하는 데 사용되는 설탕 유체를 생성하는 분비선인 꿀을 가지고 있습니다. 그들은 스스로 장기로 간주되지 않습니다.[24]
형광

한 축에 두 개 이상의 꽃이 있는 종의 꽃들을 집합적으로 꽃송이라고 부릅니다. 어떤 꽃들은 한 송이의 꽃과 비슷한 형태로 배열된 많은 작은 꽃들로 이루어져 있습니다. 이것의 일반적인 예는 매우 큰 합성물(Asteraceae) 그룹의 대부분의 구성원들입니다. 예를 들어, 데이지나 해바라기 한 송이는 꽃이 아니라 꽃 머리입니다. 수많은 꽃(또는 꽃)으로 구성된 꽃입니다.[26] 꽃차례는 특수화된 줄기와 포엽으로 알려진 변형된 잎을 포함할 수 있습니다.[27]
플로럴 다이어그램 및 공식
플로럴 포뮬러는 특정 문자, 숫자 및 기호를 사용하여 꽃의 구조를 나타내는 방법으로, 꽃에 대한 실질적인 정보를 압축된 형태로 제공합니다. 그것은 일반적으로 다양한 장기 또는 특정 종의 수를 제공하는 분류군을 나타낼 수 있습니다. 꽃 공식은 19세기 초에 개발되었으며 그 이후로 사용이 감소했습니다. Prenner et al. (2010)은 공식의 기술적 능력을 넓히기 위해 기존 모델의 확장을 고안했습니다.[28] 꽃 공식의 형식은 세계의 각 지역마다 다르지만 동일한 정보를 전달합니다.[29][30][31][32]
꽃의 구조는 꽃 그림을 통해 표현할 수도 있습니다. 도식도를 사용하면 꽃의 구조와 진화를 모두 이해하기 위한 도구로 긴 설명이나 복잡한 그림을 대체할 수 있습니다. 이러한 도표는 융합과 대칭의 존재를 포함한 다양한 기관의 상대적인 위치뿐만 아니라 구조적인 세부 사항을 포함하여 꽃의 중요한 특징을 보여줄 수 있습니다.[33]
발전
꽃은 결정적인 정단형 운모(축이 설정된 크기로 자라는 것을 의미함)에서 수정된 싹 또는 축에 생깁니다. 압축된 중간 마디를 가지고 있으며, 고전적인 식물 형태학에서 고도로 변형된 잎으로 해석되는 베어링 구조를 가지고 있습니다.[34] 그러나 세부적인 발달 연구에 따르면 수술은 종종 변형된 줄기(caulomes)처럼 시작되며 경우에 따라 가지와 유사할 수도 있습니다.[35][16] 꽃이 피는 식물의 안드로에시움 발달의 전체 다양성을 고려하여 변형된 잎(필로솜), 변형된 줄기(칼로솜), 변형된 가지(슈트) 사이의 연속체를 찾습니다.[36][37]
전이
개화로의 전환은 식물이 수명 주기 동안 만드는 주요 단계 변화 중 하나입니다. 전환은 수정과 종자 형성에 유리한 시기에 이루어져야 하며, 따라서 최대의 번식 성공을 보장합니다. 이러한 요구를 충족시키기 위해 식물은 식물 호르몬 수준의 변화와 계절에 따른 온도 및 광주기 변화와 같은 중요한 내인성 및 환경 신호를 해석할 수 있습니다.[38] 많은 다년생 식물과 대부분의 2년생 식물은 꽃을 피우기 위해 번데기화가 필요합니다. 이러한 신호의 분자적 해석은 Constans, Flowing Locus C 및 Flowing Locus T를 포함한 다양한 유전자를 포함하는 Florigen이라고 알려진 복잡한 신호의 전달을 통해 이루어집니다. 플로리겐은 번식하기 좋은 조건에서 잎에서 생산되며 싹과 성장 팁에서 작용하여 여러 가지 다양한 생리학적 및 형태학적 변화를 유도합니다.[39]

전환의 첫 번째 단계는 식물성 줄기 원시의 꽃 원시로의 전환입니다. 이것은 잎, 새싹, 줄기 조직의 세포 분화를 생식 기관으로 성장할 조직으로 바꾸기 위해 생화학적 변화가 일어나면서 발생합니다. 줄기 끝의 중앙 부분의 성장은 멈추거나 평평해지고 측면은 줄기 끝의 바깥쪽을 중심으로 둥글거나 나선형으로 돌출부를 발달시킵니다. 이 돌출부는 암술, 꽃잎, 수술 및 카르펠로 발전합니다. 일단 이 과정이 시작되면, 비록 꽃 형성 사건의 초기 시작이 어떤 환경 신호에 의존적이었다고 할지라도, 대부분의 식물에서 그것은 되돌릴 수 없고 줄기가 꽃을 피웁니다.[40]
장기발육
ABC 모델은 꽃의 발달을 담당하는 유전자를 설명하는 간단한 모델입니다. 세 가지 유전자 활동이 조합적인 방식으로 상호 작용하여 꽃 정점 융모막 내에서 원시 기관의 발달 정체를 결정합니다. 이런 유전자 기능을 A, B, C라고 합니다. A 유전자는 정점 유륜의 바깥쪽과 아래쪽 대부분의 부분에서만 발현되고, 이것은 하나의 작은 조각이 됩니다. 두 번째 소용돌이에서는 A와 B 유전자가 모두 발현되어 꽃잎이 형성됩니다. 세 번째 회오리에서는 B와 C 유전자가 상호작용하여 수술을 하고 꽃의 중심에 있는 C 유전자만으로도 카르펠이 생깁니다. 이 모델은 Arabidopsis thaliana와 스냅드래곤 Antirrhinum majus의 이상한 꽃과 돌연변이에 대한 연구를 기반으로 합니다. 예를 들어, B 유전자 기능이 상실되었을 때, 돌연변이 꽃은 정상적인 꽃잎 형성 대신 첫 번째 회오리에서는 평소와 같이, 두 번째 회오리에서도 꽃받침과 함께 생산됩니다. 세 번째 소용돌이에서는 B 기능이 부족하지만 C 기능이 존재하는 것이 네 번째 소용돌이를 모방하여 세 번째 소용돌이에서도 카르펠이 형성됩니다.[41]
기능.
꽃의 주요 목적은 개체와 종의 번식입니다[42]. 모든 꽃이 피는 식물은 이종포자성입니다. 즉, 모든 개별 식물은 두 가지 종류의 포자를 생산합니다. 미세포자는 꽃받침 안에서 감수분열에 의해 생성되고, 메가포자는 난소 안에 있는 난자 안에서 생성됩니다. 다른 곤충은 일반적으로 4개의 미세포자낭으로 구성되며 난포는 내장형 거대포자낭입니다. 두 종류의 포자는 모두 포자낭 안에서 배우자식물로 발달합니다. 모든 이종포자성 식물과 마찬가지로 배우자식물도 포자 내부에서 발달합니다. 즉, 내포자성 식물입니다.
수분

꽃은 식물의 생식 기관이기 때문에 꽃가루 내에 포함된 정자와 난소에 포함된 난자의 결합을 매개합니다.[10] 수분작용은 꽃가루가 암술머리에서 암술머리로 이동하는 것입니다.[43] 보통 꽃가루는 한 식물에서 다른 식물로 옮겨지는데, 교차 수분이라고 알려져 있지만, 많은 식물들이 자가 수분을 할 수 있습니다. 교차 수분은 종의 생존에 기여하는 유전적 변이를 허용하기 때문에 선호됩니다.[44] 많은 꽃들은 수분을 위해 바람, 물, 동물, 특히 곤충과 같은 외부 요인에 의존합니다. 그러나 [45]새, 박쥐, 심지어 일부 피그미 주머니쥐와 같은 더 큰 동물도 사용할 수 있습니다.[46][47] 이를 달성하기 위해 꽃은 한 식물에서 같은 종의 다른 식물로 꽃가루를 옮기는 것을 권장하는 특정 디자인을 가지고 있습니다. 이 과정이 일어날 수 있는 기간(꽃이 완전히 확장되어 기능하는 시기)을 개화라고 하며,[48] 따라서 수분 생물학의 연구를 신학이라고 합니다.[49]
꽃이 피는 식물은 보통 꽃가루의 이동을 최적화해야 한다는 진화적 압력에 직면하며, 이는 일반적으로 꽃의 형태와 식물의 행동에 반영됩니다.[50] 꽃가루는 여러 '벡터' 또는 방법을 통해 식물 사이에 전달될 수 있습니다. 꽃이 피는 식물의 약 80%는 생물학적 또는 살아있는 매개체를 사용합니다. 다른 것들은 비생물적인 또는 무생물적인 벡터를 사용하고 일부 식물은 여러 벡터를 사용하지만 대부분은 고도로 전문화되어 있습니다.[51]
일부는 이 그룹들 사이에 또는 바깥에 있지만,[52] 대부분의 꽃들은 다음과 같은 두 개의 넓은 그룹의 수분 방법으로 나눌 수 있습니다.
생물수분
생물학적 매개체를 사용하는 꽃은 곤충, 박쥐, 새 또는 다른 동물을 유인하고 사용하여 한 꽃에서 다음 꽃으로 꽃가루를 옮깁니다. 종종 그들은 모양에 특화되어 있으며 꽃가루 알갱이가 꽃가루의 유인물(예를 들어, 꿀, 꽃가루 또는 짝)을 찾아 착륙할 때 꽃가루 알갱이가 꽃가루 매개자의 몸으로 전달되도록 보장하는 수술 배열을 가지고 있습니다.[53] 같은 종의 많은 꽃에서 이 유인 물질을 추구하기 위해 꽃가루 매개자는 꽃가루를 방문하는 모든 꽃의 동일한 뾰족한 정밀도로 배열된 스티그마로 옮깁니다.[54] 많은 꽃들이 수분을 보장하기 위해 꽃 부분들 사이의 단순한 근접성에 의존하는 반면, 다른 꽃들은 수분을 보장하고 자가 수분을 방지하기 위해 정교한 디자인을 가지고 있습니다.[44] 꽃은 곤충, 새, 박쥐, 도마뱀,[47] 심지어 달팽이와 민달팽이를 포함한 동물을 사용합니다.[55]
유치방법

식물들은 한 장소에서 다른 장소로 이동할 수 없기 때문에, 많은 꽃들은 분산된 개체군에 있는 개체들 사이에 꽃가루를 옮길 수 있도록 동물들을 유인하기 위해 진화해 왔습니다. 가장 일반적으로 꽃은 곤충에 의해 수분되는 것으로, 곤충을 사랑하는 그리스어로 문자 그대로 "곤충을 사랑하는" 것으로 알려져 있습니다.[57] 이 곤충들을 유인하기 위해 꽃들은 영양이 풍부한 꿀을 찾는 동물들을 유인하는 다양한 부분에 보통 꿀이라는 분비선을 가지고 있습니다.[58] 어떤 꽃들은 꿀보다는 기름을 생산하는 elaiophore라고 불리는 분비선을 가지고 있습니다.[59] 새들과 벌들은 색시를 가지고 있어서 그들이 다채로운 꽃을 찾을 수 있게 해줍니다.[60] 어떤 꽃들은 수분 매개자가 어디서 꿀을 찾을지를 보여주는 과즙 안내자라고 불리는 패턴을 가지고 있습니다; 그것들은 벌과 다른 곤충들에게 보이는 자외선 아래에서만 볼 수 있을지도 모릅니다.[61]
꽃은 또한 향기로 꽃가루 매개자를 유인하기도 하지만, 모든 꽃 향기가 인간에게 매력적인 것은 아닙니다. 많은 꽃들은 썩은 살에 이끌려 죽은 동물의 냄새가 나는 곤충에 의해 수분됩니다. 이 꽃들은 종종 카리온 꽃이라고 불리는데, 여기에는 라플레시아속 식물과 타이탄 아룸이 포함됩니다.[60] 박쥐와 나방을 포함한 야간 방문객에 의해 수분되는 꽃들은 꽃가루 매개자를 유인하기 위해 향기에 집중할 가능성이 높기 때문에 그러한 꽃들은 대부분 흰색입니다.[62] 박쥐에 의해 수분되는 일부 식물은 꽃 위에 음파 반사 꽃잎을 가지고 있는데, 이것은 박쥐가 그것들을 찾는데 도움이 되고,[63] 한 종의 선인장인 Espostoa frutescens는 대신 박쥐의 초음파를 흡수하는 두발이라고 불리는 소리를 흡수하는 털과 털로 둘러싸인 꽃을 가지고 있습니다.[64]
꽃은 또한 모양에 특화되어 있으며 꽃가루 알갱이가 유인 물질을 찾아 착륙할 때 꽃가루 알갱이가 꽃가루 매개자의 몸으로 전달되도록 보장하는 수술 배열을 가지고 있습니다. 다른 꽃들은 수분 매개체를 유인하기 위해 모방이나 의사 교배를 사용합니다. 예를 들어, 많은 난초들은 색깔, 모양, 그리고 향기가 암컷 벌이나 말벌을 닮은 꽃을 만듭니다. 수컷들은 짝을 찾아 한 꽃에서 다음 꽃으로 옮겨 다니며 꽃을 수분시킵니다.[65][66]
꽃가루 매개자 관계
많은 꽃들이 하나 또는 몇 개의 특정 수분 생물들과 밀접한 관계를 가지고 있습니다. 예를 들어, 많은 꽃들은 한 종의 특정 곤충만을 유인하기 때문에 성공적인 번식을 위해 그 곤충에 의존합니다. 이 밀접한 관계는 꽃과 꽃가루 매개자가 서로의 필요에 맞추기 위해 오랜 시간에 걸쳐 함께 발전했기 때문에 공진화의 한 예입니다.[67] 그러나 이러한 밀접한 관계는 소멸의 부정적인 영향을 복합적으로 미치지만, 그러한 관계에 있는 구성원 중 하나의 소멸은 거의 확실하게 다른 구성원의 소멸을 의미하기 때문입니다.[68]
비생물적 수분작용


비생물적이거나 무생물적인 매개체를 사용하는 꽃들은 꽃가루를 한 꽃에서 다음 꽃으로 옮기기 위해 바람이나 훨씬 덜 일반적으로 물을 사용합니다.[51] 바람에 흩날리는 (유성적인) 종에서는 작은 꽃가루 알갱이들이 바람에 [69]의해, 때로는 수천 킬로미터씩 다른 꽃으로 옮겨집니다. 일반적인 예로는 풀, 자작나무, 그리고 파갈레스목,[70] 끈끈이, 그리고 많은 사초목에 속하는 많은 다른 종들이 있습니다. 그들은 꽃가루 매개자를 끌어들일 필요가 없기 때문에 크고, 화려하거나, 화려한 꽃이 자라지 않는 경향이 있고, 꿀이나 눈에 띄는 향기가 없습니다. 이 때문에 식물들은 보통 수 천 개의 작은 꽃들을 가지고 있는데, 이 꽃들은 꽃가루를 받을 확률을 높이기 위해 비교적 크고 깃털 같은 스티그마를 가지고 있습니다.[65] 곤충성 꽃의 꽃가루는 보통 크고 끈적이며 단백질이 풍부한 반면에, 곤충의 경우 일반적으로 입자가 작고, 매우 가볍고, 매끄럽고, 영양가가 거의 없습니다.[71][72] 바람이 꽃가루를 효과적으로 집어 들고 운반하기 위해, 꽃들은 일반적으로 길고 얇은 필라멘트의 끝에 느슨하게 붙어 있거나 바람에 움직이는 캣킨 주위에 꽃가루가 형성됩니다. 이것의 더 드문 형태는 개별 꽃들이 바람에 의해 움직일 수 있는 것을 포함합니다(Pendulous). 심지어 덜 일반적으로; 꽃가루를 바람에 방출하기 위해 폭발하는 꽃들.[71]
물을 통한 수분(hydrophily)은 훨씬 더 드문 방법으로, 생물학적으로 수분된 꽃의 약 2%에서만 발생합니다.[51] 이것의 일반적인 예로는 Calitriche autumnalis, Valisneria spiralis 및 일부 해초가 있습니다. 이 그룹의 대부분의 종들이 공유하는 한 가지 특징은 꽃가루 알갱이 주변에 엑신, 즉 보호층이 없다는 것입니다.[73] Paul Knuth는 1906년에 두 가지 유형의 친수성 수분을 확인했고 Ernst Schwarzenbach는 1944년에 세 번째 유형을 추가했습니다. 크누트는 자신의 두 그룹을 'Hyphydrogamy'와 더 흔한 'Ephydrogamy'[74]라고 이름 지었습니다. 수생식물에서 수분작용은 수면 아래에서 일어나므로 꽃가루 알갱이는 일반적으로 음으로 부력이 있습니다. 이 방법을 보여주는 해양 식물의 경우 대개 스티그마는 뻣뻣한 반면 담수종은 작고 깃털 같은 스티그마를 가지고 있습니다.[75] 수생대내 수분작용은 물 표면에서 일어나기 때문에 꽃가루는 부유할 수 있는 밀도가 낮지만 많은 사람들이 뗏목을 사용하고 소수성을 띠고 있습니다. 해양 꽃은 떠다니는 실 같은 스티그마를 가지고 있고 조수에 적응할 수 있는 반면 담수종은 물에 움푹 들어간 곳을 만듭니다.[75] Schwarzenbach가 제시한 세 번째 범주는 꽃가루를 운반을 통해 물 위로 운반하는 꽃들입니다. 이것은 떠다니는 식물(Lemnoideae)부터 수술용 꽃(Valisneria)까지 다양합니다. 이 그룹에 속하는 대부분의 종들은 건조하고 구형의 꽃가루를 가지고 있으며, 때로는 더 큰 덩어리로 형성되기도 하며, 물 속에 움푹 패인 곳을 형성하는 암컷 꽃을 가지고 있습니다. 운반 방법은 다양합니다.[75]
매커니즘
꽃은 교차 수분과 자가 수분의 두 가지 메커니즘에 의해 수분될 수 있습니다. 각각의 장점과 단점이 있기 때문에 어떤 메커니즘도 다른 메커니즘보다 명백하게 더 나은 것은 아닙니다. 식물은 서식지와 생태학적 틈새에 따라 이러한 메커니즘 중 하나 또는 둘 모두를 사용합니다.[76]
교차수분
교차 수분은 같은 종의 다른 식물의 꽃가루에 의한 목탄의 수분입니다. 다른 식물의 꽃가루 안에 들어 있는 정자의 유전적 구성이 다르기 때문에, 이들의 조합은 성생식 과정을 거쳐 유전적으로 구별되는 새로운 식물이 될 것입니다. 각각의 새로운 식물은 유전적으로 구별되기 때문에, 다른 식물들은 생리적이고 구조적인 적응에 있어서 다양성을 보이기 때문에 전체적으로 개체군은 환경에서 불리한 발생에 더 잘 대비됩니다. 따라서 교차 수분은 종의 생존을 증가시키고 보통 이러한 이유로 꽃들이 선호합니다.[44][77]
자가수분
_flowers.jpg)
자가수분은 같은 꽃이나 같은 식물에 있는 다른 꽃의 꽃가루에 의해 꽃의 목관이 수분되어 무성생식을 통해 유전자 클론이 생성되는 [44]것입니다. 이는 종자 생산의 신뢰성, 생산 가능 속도를 높이고 필요한 에너지 양을 낮춥니다.[78] 그러나 가장 중요한 것은 유전적 변이를 제한한다는 것입니다. 또한 자가 수분은 주로 열성 유해 돌연변이의 발현으로 인해 근친 우울증을 유발합니다.[79][80]
같은 꽃이나 식물의 꽃가루에 의해 배란이 수정되는 극단적인 자가 수정의 경우는 민들레가 많은 것처럼 항상 자가 수정을 하는 꽃에서 발생합니다.[81] 어떤 꽃들은 자가수분을 해서 꽃이 열리기 전에 절대로 열리지 않거나 자가수분을 하지 않는 꽃들이 있습니다; 이 꽃들은 cleistogamy라고 불립니다; 예를 들어, Viola속의 많은 종들이 이것을 보여줍니다.[82]
반대로, 많은 종의 식물은 자가 수분을 방지하여 자가 수정을 방지하는 방법을 가지고 있습니다. 같은 식물의 암수꽃과 암수꽃이 동시에 출현하거나 성숙하지 않거나 같은 식물의 꽃가루가 배란을 수정하지 못할 수 있습니다. 자기 꽃가루에 대한 화학적 장벽을 가지고 있는 후자의 꽃 유형을 자가불적합이라고 합니다.[23][83] Clianthus puniceus(사진)에서는 자가수분을 '보험정책'으로 전략적으로 활용하고 있습니다. 꽃가루 매개자, 이 경우 새가 C. puniceus를 방문하면 새의 꽃가루가 낙인 덮개를 문지르고 낙인 속으로 들어갈 수 있습니다. 그러나 수분 매개자가 방문하지 않으면 꽃 자체의 꽃차례가 자가 수분을 통해 꽃을 수분시킬 수 있도록 스티그마틱 덮개가 자연적으로 떨어집니다.[78]
알러지
꽃가루는 천식 및 기타 호흡기 알레르기의 큰 원인이며, 이는 전 세계 사람의 10~50%에게 영향을 미칩니다. 이 숫자는 기후 변화로 인해 기온이 상승함에 따라 식물들이[citation needed] 꽃가루를 더 많이 생산하고 있다는 것을 의미하기 때문에 증가하는 것으로 보이며, 이는 알레르기 유발성도 더 높습니다. 그러나 꽃가루는 크기가 작고 자연 환경에 널리 분포하기 때문에 피하기가 어렵습니다. 알레르기를 일으키는 꽃가루의 대부분은 풀, 자작나무, 참나무, 그리고 끈끈이와 같은 바람이 분산된 꽃가루에 의해 생성되는 것입니다. 꽃가루에 있는 알레르기 물질은 수분 과정에서 필요할 것으로 생각되는 단백질입니다.[84][85]
수정
_(1914)_(14590686978).jpg)
유대교 회당이라고도 불리는 수정은 수분작용 후에 일어나는데, 이것은 꽃가루가 수술에서 카르펠로 이동하는 것입니다. 원형질체의 융합인 원형질체와 핵의 융합인 핵분열체를 모두 포함합니다. 꽃가루가 꽃의 오명에 떨어지면 꽃가루 관이 만들어지기 시작합니다. 꽃가루 관은 화풍을 타고 난소로 흘러갑니다. 난소의 가장 중앙 부분을 관통한 후 난자 장치로 들어가 하나의 시너지 효과를 냅니다. 이 시점에서 꽃가루 관의 끝은 터져서 두 개의 정자 세포를 방출하는데, 그 중 하나는 난자로 가는 동시에 세포막과 원형질의 대부분을 잃게 됩니다. 그러면 정자의 핵이 난자의 핵과 융합하여 이배체(각 염색체의 두 개의 사본) 세포인 접합체(zygote)가 형성됩니다.[86]
수정에서는 형질전환(플라즈마가미), 즉 성세포 전체가 융합되는 결과만 나타나는 반면, 속씨식물(꽃피는 식물)에서는 핵분열과 형질전환을 모두 포함하는 이중 수정(double militation)이라고 알려진 과정이 발생합니다. 이중 수정에서 두 번째 정자 세포도 이어서 상승기로 들어가 중앙 세포의 두 극성 핵과 융합합니다. 세 개의 핵은 모두 반수체이기 때문에 3배체인 배유핵이 크게 됩니다.[86]
종자발육

접합체가 형성된 후에 그것은 유사분열이라고 불리는 핵과 세포 분열을 통해 성장하기 시작하고 결국 작은 세포 그룹이 됩니다. 그것의 한 부분은 배아가 되고, 다른 부분은 현탁액이 됩니다; 배아를 배유 안으로 강제로 집어넣고 나중에 감지할 수 없는 구조입니다. 이때 두 개의 작은 원시도 형성되는데, 이것이 나중에 떡잎이 되어 에너지 저장소로 사용됩니다. 이러한 원시 식물 중 하나에서 자라는 식물을 단엽 식물이라고 하고, 두 개에서 자라는 식물을 쌍엽 식물이라고 합니다. 다음 단계는 어뢰 단계라고 하며, 다음과 같은 몇 가지 주요 구조의 성장을 포함합니다: 래디클(배아근), 에피코틸(배아줄기), 하이포코틸(근/슛접합). 마지막 단계에서는 종자 주위에 혈관 조직이 발달합니다.[87]
과실발육
씨가 배란에서 형성되고 있는 난소는 열매로 자랍니다. 다른 모든 주요 꽃 부분들은 이 발달 중에 죽습니다: 스타일, 낙인, 꽃받침, 수술 그리고 꽃잎을 포함합니다. 열매는 외피층, 중피층, 살이 많은 부분, 내피층, 내피층의 세 가지 구조를 가지고 있는 반면, 열매 벽은 과피층이라고 불립니다. 과일마다 크기, 모양, 질긴 정도, 두께가 다릅니다. 종자의 분산을 촉진하거나 가능하게 하고, 그렇게 하면서 종자를 보호하는 것이 바로 과실의 목적인 종자 분산의 방법과 직결되기 때문입니다.[87]
종자분산
.jpg)


일반적으로 꽃의 수분, 수정, 그리고 마지막으로 씨앗과 열매가 발달한 후에 식물에서 과일을 멀리 분산시키는 메커니즘을 사용합니다.[91] 속씨식물(꽃피는 식물)에서 씨앗은 모체와 딸 식물 사이의 경쟁을 [92]강요하지 않고 새로운 영역의 집락화를 가능하게 하기 위해 식물에서 멀리 분산됩니다. 많은 식물들이 이들 중 하나 또는 그 이상에 속하지만, 그것들은 종종 두 가지 범주로 나뉩니다.[93]
알로코리
알로코리에서 식물은 외부 벡터 또는 매개체를 사용하여 씨앗을 식물로부터 멀리 운반합니다. 이것들은 새나 개미와 같은 생물(살아있는)일 수도 있고, 바람이나 물과 같은 생물(살아있는)일 수도 있습니다.[93][94][95]
생체벡터
많은 식물들이 생물학적 벡터를 사용하여 씨앗을 멀리 분산시킵니다. 이 방법은 동물원(Zoochory)이라는 포괄적인 용어에 해당하는 반면, 엔도주코리(Endozoochory)는 과일을 먹을 수 있도록 동물을 유인하기 위해 과일을 재배하도록 적응된 식물을 말합니다. 일단 먹으면 그들은 일반적으로 동물의 소화기관을 거쳐 식물로부터 멀리 떨어져 나갑니다.[95] 일부 씨앗은 동물의 전어에서 지속되거나 심지어 동물을 통과한 후 더 잘 발아하도록 특별히 조정되었습니다.[96][97] 새(오르니토코리), 박쥐(키롭토코리), 설치류, 영장류, 개미(미르메코리),[98] 비조류 사우롭시드(사우로코리), 일반 포유류(맘마리오코리),[96] 심지어 물고기도 먹을 수 있습니다.[99] 일반적으로 과일은 육질이고 영양가가 높으며 분산제에 대한 추가 "보상"으로 화학적 유인 물질을 가질 수 있습니다. 이것은 형태학적으로 더 많은 펄프, 아릴, 그리고 때때로 다른 살이 많은 구조인 엘라이오솜(주로 개미를 위한)이 존재할 때 반영됩니다.[100]
에피주코리(Epizoochory)는 아카에나(Acaena) 속의 많은 종들처럼 씨앗이 동물에 달라붙도록 적응되어 그런 방식으로 분산된 식물에서 발생합니다.[101] 일반적으로 이 식물 종자에는 동물이 쉽게 잡기 위해 갈고리나 점성이 있는 표면이 있는데, 여기에는 털이 있는 새와 동물이 포함됩니다. 어떤 식물들은 동물들이 씨앗을 퍼뜨리도록 속이기 위해 미메시스, 즉 모방을 사용하는데 이것들은 종종 특별하게 적응된 색깔을 가지고 있습니다.[100][102]
동물원의 마지막 유형은 Synzoochory라고 불리는데, 이것은 씨앗의 소화나 의도치 않게 몸에 씨앗을 운반하는 것이 아니라 동물들이 의도적으로 씨앗을 운반하는 것을 포함합니다. 이것은 보통 많은 새들과 모든 개미들에게 사용되는 동물(스토마토코리라고 불리는)의 입이나 부리에 있습니다.[103]

.jpg)
비생물적 매개체
비생물적 분산 식물은 바람, 물 또는 자체 메커니즘의 매개체를 사용하여 씨앗을 그들로부터 멀리 운반합니다.[95][94] 기억은 바람을 매개체로 사용하여 식물의 씨앗을 분산시키는 것을 포함합니다. 이 씨앗들은 바람을 타고 이동해야 하기 때문에 거의 항상 작고 때로는 먼지처럼 생기고, 표면적 대 부피 비율이 높으며, 때로는 백만 개까지 대량으로 생산됩니다. 텀블 잡초와 같은 식물은 씨앗이 바람과 함께 굴러 떨어지게 하기 위해 전체 촬영을 분리합니다. 또 다른 일반적인 적응은 씨앗이 공기 중에 더 오래 머물게 하고 따라서 더 멀리 이동하게 하는 날개, 깃털 또는 풍선과 같은 구조물입니다.
Hydrochory에서 식물은 물을 통해 씨앗을 분산시키도록 적응되어 있기 때문에 일반적으로 부력이 있고 물에 대한 상대 밀도가 낮습니다. 일반적으로 종자는 소수성 표면, 작은 크기, 털, 점액, 기름 및 때로는 종자 내의 공기 공간과 함께 형태학적으로 적응됩니다.[100] 이 식물들은 조류의 표면, 조류의 표면 아래, 그리고 식물에 비가 내려 식물에 씨앗이 흩어지는 세 가지 범주로 나뉩니다.[104]
오토코리
오토코리에서, 식물은 씨앗을 그들로부터 멀리 운반하기 위해 그들 자신의 벡터를 만듭니다. 이를 위한 적응은 일반적으로 과일이 폭발하여 Hura crepitans와 같이 탄도적으로 씨앗을 강제로 제거하는 것을 포함하거나 [105]때로는 기어가는 디아스포라를 생성하는 것을 포함합니다.[100] 이러한 방법은 씨앗을 분산시킬 수 있는 거리가 상대적으로 작기 때문에 외부 벡터와 쌍을 이루는 경우가 많습니다.[102]
진화
육상 식물은 약 4억 2천 5백만 년 동안 존재해 왔지만, 최초의 육상 식물은 수생 식물인 포자를 단순하게 적응시켜 번식시켰습니다. 바다에서 식물과 일부 동물들은 단순히 자신의 유전자 복제물을 흩뿌려 다른 곳에서 떠다니며 자랄 수 있습니다. 이것이 초기 식물이 번식하는 방법입니다. 그러나 식물들은 바다보다 육지에서 훨씬 더 가능성이 높은 건조 및 기타 피해를 처리하기 위해 이러한 사본을 보호하는 방법을 곧 진화시켰습니다. 보호자는 꽃을 진화시키지는 않았지만 씨앗이 되었습니다. 초기 종자를 가진 식물에는 은행과 침엽수가 있습니다.

꽃이 어떻게 진화했는지 정확하게 보여주는 화석 증거는 아직 없습니다. 특히 씨앗 양치식물과 같은 멸종된 여러 종류의 짐노스프들이 꽃의 조상으로 제안되어 왔습니다. 화석 기록에 비교적 현대적인 꽃들이 갑자기 등장한 것은 진화론에 너무나 큰 문제를 던져 찰스 다윈에 의해 "끔찍한 미스터리"라고 불렸습니다.
최근에 발견된 고세균(Archiefructus)과 같은 배유 화석은 배유 화석의 추가적인 발견과 함께 배유의 특성이 어떻게 일련의 단계에서 획득되었는지를 시사합니다. 중국에서 온 꽃이 피는 식물의 초기 화석인 Archefractus yannes는 약 1억 2천 5백만 년 전의 것입니다.[106][107] 훨씬 이전부터 중국에서는 1억 2,500만년에서 1억 3,000만년 된 Archiefructus sinensis가 있습니다. 2015년에는 1억 3천만 년 된 식물(1억 3천만 년 된 Montsechia vidalii, 스페인에서 발견됨)이 1억 3천만 년 된 것으로 주장되었습니다.[108] 2018년, 과학자들은 가장 초기의 꽃들이 약 1억 8천만년 전에 시작되었다고 보고했습니다.[109]
_fragment.jpg)
최근 DNA 분석(분자 계통학)[110]에 따르면 태평양 뉴칼레도니아 섬에서 발견된 암보렐라 트리코포다는 자매군의 나머지 개화 식물의 유일한 종이며 형태학적 연구에 따르면 가장 초기 개화 식물의 특징일 수 있는 특징을 가지고 있습니다.[111]
백악기나 그 직전에 꽃이 피었다는 확실한 증거 [112][113]외에도 2억 5천만 년 전에 꽃이 피었다는 정황적 증거가 있습니다. 식물이 꽃을 방어하기 위해 사용하는 화학물질인 올레인은 그 당시 진화한 [114]거간토프테리드를 포함하여 오래된 화석 식물에서 발견되었으며, 비록 그들이 꽃이 피는 식물 그 자체는 아니지만, 왜냐하면 그들의 줄기와 가시만 자세히 보존되어 있는 것이 발견되었기 때문입니다. 그것은 석화의 가장 초기 예 중 하나입니다.
꽃은 유전적으로 정상적인 잎과 줄기 성분을 식물에 적응시킨 것일 뿐이기 때문에 잎과 줄기 구조의 유사성은 매우 중요할 수 있습니다. 왜냐하면 꽃은 보통 새로운 싹을 형성하는 역할을 하는 유전자의 조합입니다.[115] 가장 원시적인 꽃들은 다양한 수의 꽃 부분을 가지고 있었던 것으로 생각되며, 종종 서로 떨어져 있습니다. 꽃들은 나선형으로 자라고, 양성화(식물에서 이것은 같은 꽃에 있는 암수 부분을 의미함)되고, 난소(암수 부분)에 의해 지배되는 경향이 있었을 것입니다. 꽃이 더 발달함에 따라 일부 변형은 서로 융합된 부분을 발전시켰고, 훨씬 더 구체적인 숫자와 디자인을 가지고 꽃이나 식물당 특정한 성별을 갖거나 최소한 "난소 열위"를 했습니다.
일반적인 가정은 꽃의 기능은 처음부터 동물을 번식 과정에 참여시키는 것이었습니다. 꽃가루는 밝은 색과 명백한 모양 없이 흩어질 수 있으며, 따라서 다른 이점을 제공하지 않는 한 식물의 자원을 사용하는 것이 부담이 될 것입니다. 꽃이 갑자기 완전히 발달한 이유 중 하나는 꽃이 섬처럼 고립된 환경에서 진화했기 때문인데, 꽃을 가진 식물이 오늘날 많은 섬 종들이 발달하는 방식과 같은 특정 동물(예를 들어 말벌)과 매우 전문화된 관계를 발전시킬 수 있었기 때문입니다. 오늘날 무화과 말벌이 하는 것처럼 한 식물에서 다른 식물로 꽃가루를 옮기는 가상의 말벌과의 이러한 공생 관계는 결국 식물과 그들의 파트너 모두가 고도의 전문화를 개발하는 결과를 가져올 수 있었습니다. 섬 유전학은 특히 열등한 과도기적 형태를 필요로 하는 것으로 보이는 급진적인 적응에 관한 한 종분화의 일반적인 원천으로 여겨집니다. 말벌의 예는 우발적인 것이 아닙니다. 공생 식물 관계를 위해 특별히 진화한 것으로 보이는 벌들은 말벌의 후손입니다.
마찬가지로, 식물 번식에 사용되는 대부분의 과일은 꽃의 일부가 확대되는 것에서 나옵니다. 이 과일은 종종 그것을 먹으려는 동물들에게 의존하는 도구이며, 따라서 그 안에 들어 있는 씨앗을 흩뿌립니다.
그러한 많은 공생 관계가 본토 유기체와의 경쟁에서 살아남기에는 너무 취약한 상태로 남아 있지만, 꽃은 육지 식물 생명체의 지배적인 형태가 되기 위해 (그들의 실제 기원이 무엇이든) 퍼져나가는 비정상적으로 효과적인 생산 수단임이 증명되었습니다.
꽃의 진화는 오늘날까지 계속되고 있습니다; 현대의 꽃들은 자연에서 수분될 수 없을 정도로 인간에 의해 아주 깊은 영향을 받아왔습니다. 많은 현대의 길들여진 꽃들은 땅이 흐트러져야 싹이 트는 단순한 잡초였습니다. 일부는 인간의 농작물과 함께 자라는 경향이 있었고, 가장 예쁜 것은 아름다움 때문에 뽑히지 않아 인간의 애정에 대한 의존과 특별한 적응을 하게 되었습니다.[116]
색
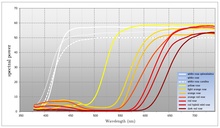
많은 꽃이 피는 식물은 식물이 끌어들이려는 수분 매개체의 가시 파장 범위 내에서 가능한 한 많은 빛을 반사합니다. 가시광선의 전 범위를 반사하는 꽃은 일반적으로 인간 관찰자에 의해 흰색으로 인식됩니다. 흰 꽃의 중요한 특징은 눈에 보이는 스펙트럼에 걸쳐 똑같이 반사된다는 것입니다. 많은 꽃이 피는 식물들이 꽃가루 매개자를 유인하기 위해 흰색을 사용하는 반면, 색상의 사용 또한 널리 퍼져 있습니다 (심지어 같은 종 내에서도). 색상을 사용하면 꽃이 피는 식물이 유인하려는 수분 매개체에 대해 더 구체적으로 설명할 수 있습니다.[citation needed] 인간의 색 재현 기술(CMYK)이 사용하는 컬러 모델은 스펙트럼을 광범위한 흡수 영역으로 나누는 색소의 변조에 의존합니다. 대조적으로 꽃이 피는 식물은 흡수와 반사 사이의 전환점 파장을 이동시킬 수 있습니다. 대부분의 꽃가루 매개자의 시각계가 가시 스펙트럼을 원형으로 본다고 가정하면 꽃이 피는 식물은 스펙트럼의 한 영역의 빛을 흡수하고 다른 영역의 빛을 반사하여 색을 생성한다고 말할 수 있습니다. CMYK를 사용하면 광범위한 흡수 영역의 진폭의 함수로 색상이 생성됩니다. 대조적으로 꽃이 피는 식물은 반사되는 빛의 주파수(또는 오히려 파장)를 수정하여 색을 만듭니다. 대부분의 꽃은 스펙트럼의 파란색에서 노란색 영역의 빛을 흡수하고 스펙트럼의 녹색에서 빨간색 영역의 빛을 반사합니다. 많은 종의 꽃이 피는 식물의 경우, 그것들이 생산하는 색을 특징짓는 전환점입니다. 색상은 흡수와 반사 사이의 전환점을 이동함으로써 조절될 수 있으며 이러한 방식으로 개화 식물은 유인하려는 수분 매개체를 지정할 수 있습니다.[citation needed] 일부 개화 식물은 흡수 영역을 조절하는 능력도 제한적입니다. 이것은 일반적으로 파장에 대한 제어만큼 정확하지 않습니다. 인간 관찰자들은 이것을 포화도(색의 흰색의 양)로 인식할 것입니다.
고전분류학
.png)
식물의 분류와 식별을 연구하는 식물 분류학에서 식물의 꽃의 형태는 광범위하게 사용되며 수천 년 동안 사용되어 왔습니다. 비록 식물 분류학의 역사가 적어도 기원전 300년 정도까지 거슬러 올라가지만,[118] 테오프라스토스의 저술로 현대 과학의 기초는 18세기와 19세기의 작품들에 기초를 두고 있습니다.[119]
칼 린네 (1707–1778)는 대부분의 직장 생활을 자연사 교수로 보낸 스웨덴의 식물학자입니다. 그의 대표적인 저서인 1757년의 종 플랜타룸은 그의 분류 체계와 이항 명명법의 개념을 제시하고 있으며, 그 중 후자는 오늘날에도 여전히 전 세계에서 사용되고 있습니다.[119][note 1] 그는 주로 수술의 수, 길이 및 결합을 기반으로 24개의 클래스를 식별했습니다. 처음 10개의 클래스는 직접적으로 수술의 수를 따르지만([117]옥탄드리아는 8개의 수술 등이 있습니다), 11개 클래스는 11-20개의 수술, 12개와 13개 클래스는 20개의 수술이 있으며 부착 지점만 다릅니다. 다음 5개의 분류는 수술의 길이와 마지막 5개는 식물의 번식 능력의 특성, 수술이 자라는 곳, 꽃이 숨겨져 있거나 존재하는 경우(예: 양치식물)를 다룹니다. 이 분류법은 인공적이기는 하지만 이후 70년 동안 광범위하게 사용되다가 다른 식물학자의 시스템으로 대체되었습니다.[117][120]
Antoine Laurent de Jussieu (1748–1836)는 1787년 그의 연구로 식물 속 (Generas plantarum: secundum or dines naturales disposita)을 대신하여 식물을 분류하는 새로운 방법을 제시한 프랑스의 식물학자입니다. 식물은 자엽이 있는 경우의 수와 수술 위치로 나누었습니다.[120] 그 다음으로 중요한 분류 체계는 19세기 후반 식물학자 조셉 돌턴 후커 (1817–1911)와 조지 벤담 (1800–1884)으로부터 왔습니다. 그들은 드 쥐슈와 오귀스트의 초기 작품인 피라무스 드 칸돌을 바탕으로 아직도 세계의 많은 허브리아에서 사용되는 시스템을 고안했습니다. 식물은 자엽의 수와 꽃의 성질에 의해 가장 높은 수준으로 나뉘었다가, 목(과), 속(과), 종(종)으로 떨어졌습니다. 이 분류 체계는 1862년에서 1883년 사이에 3권으로 그들의 플랜타룸(Genus plantarum)에 출판되었습니다.[121] 일부 환경에서는 가장 높이 평가되고 "최고의 분류 시스템"으로 간주됩니다.[122]
다윈의 '종의 기원에 대하여' 이후 과학적 사고의 발달로, 많은 식물학자들이 더 많은 계통발생학적 방법을 사용하게 되었고, 유전자 염기서열 분석, 세포학 및 고생물학의 사용이 점점 더 보편화되었습니다. 그럼에도 불구하고, 꽃의 성질이나 꽃의 개화와 같은 형태학적 특성은 여전히 식물 분류학의 기반을 이루고 있습니다.[122][123]
상징성

많은 꽃들은 서양 문화에서 중요한 상징적 의미를 가지고 있습니다.[124] 꽃에 의미를 부여하는 관행은 플로리그래피로 알려져 있습니다. 일반적인 예로는 다음과 같은 것들이 있습니다.
- 붉은 장미는 사랑, 아름다움, 열정의 상징으로 주어집니다.[125]
- 양귀비는 죽음의 시기에 위로의 상징입니다. 영국, 뉴질랜드, 호주, 캐나다에서는 전쟁 중에 전사한 군인들을 기념하기 위해 빨간 양귀비를 입습니다.
- 이리즈/릴리는 매장에서 "부활/생명"을 나타내는 상징으로 사용됩니다. 그것은 또한 별(태양)과 꽃잎이 피거나 빛나는 것과 관련이 있습니다.
- 데이지는 순수의 상징입니다.

다양하고 다채로운 모습 때문에 꽃은 오랫동안 시각 예술가들이 가장 좋아하는 주제이기도 했습니다. 반 고흐의 해바라기 시리즈나 모네의 수련과 같은 유명한 화가들의 가장 유명한 그림들 중 일부는 꽃입니다. 또한 꽃은 영구적인 3차원 꽃 예술 작품을 만들기 위해 건조, 동결 건조 및 압착됩니다.
조지아 오키프, 이모겐 커닝햄, 베로니카 루이스 드 벨라스코, 주디 시카고와 같은 예술가들의 작품과 사실 아시아와 서양 고전 예술에서 볼 수 있듯이 예술 [126]내의 꽃들은 여성의 생식기를 대표하기도 합니다. 세계의 많은 문화들은 꽃을 여성성과 연관시키는 경향이 두드러집니다.
섬세하고 아름다운 꽃의 다양성은 특히 18세기에서 19세기 낭만주의 시대의 수많은 시인들의 작품에 영감을 주었습니다. 유명한 예로는 윌리엄 워즈워스의 '구름처럼 외롭게 떠돌아다녔다'와 윌리엄 블레이크의 '아! 해바라기'가 있습니다.
꿈에서의 그들의 상징성 또한 논의되어 왔는데, "꽃을 피우는 잠재력"을 포함한 가능한 해석들이 있습니다.[127]
꽃, 정원, 봄의 계절의 로마 여신은 플로라입니다. 봄, 꽃, 자연의 그리스 여신은 클로리스입니다.
힌두교 신화에서 꽃은 중요한 위치를 차지합니다. 힌두교의 3대 신 중 하나인 비슈누는 연꽃 위에 똑바로 서 있는 모습이 자주 묘사됩니다.[128] 비슈누와의 연관성과는 별개로, 힌두교의 전통은 연꽃이 영적인 의미가 있다고 생각합니다.[129] 예를 들어, 그것은 힌두교의 창조 이야기에 등장합니다.[130]
인간의 이용
이 섹션은 검증을 위해 추가 인용이 필요합니다. (2018년 11월)(본의 및 알아보기 |

역사는 꽃이 수천 년 동안 다양한 목적을 위해 인간에 의해 사용되어 왔다는 것을 보여줍니다. 이것의 초기 예는 약 4,500년 전 고대 이집트에서 꽃이 여성의 머리를 장식하는 데 사용되었던 것입니다. 모네의 수련이나 윌리엄 워즈워스의 수선화에 대한 시 "나는 구름처럼 외롭게 방황했다"와 같은 꽃들은 예술에 몇 번이고 영감을 주었습니다.[131]

현대에 사람들은 꽃과 꽃이 피는 식물을 재배하고, 구입하고, 입거나, 그 밖의 방법들을 모색해 왔습니다. 이는 부분적으로 그들의 기분 좋은 외모와 냄새 때문입니다. 전세계적으로, 사람들은 그들의 삶에서 중요한 사건들을 기념하기 위해 꽃을 사용합니다.
- 신생아 또는 그리스도 성탄화의 경우
- 사교 행사나 휴일에 착용하는 코사지나 부토니어로
- 사랑이나 존경의 증표로서
- 신부 파티를 위한 웨딩 플라워 및 결혼식장 장식용
- 집 안의 장식을 밝게 하는 것처럼.
- 뱃놀이 파티, 웰컴홈 파티, '당신 생각' 선물에 대한 추억의 선물로
- 장례식 꽃과 슬픔에 잠긴 사람들에 대한 위로의 표현을 위해
- 예배용. 기독교에서는 종종 찬송가 꽃들이 교회를 장식합니다.[132] 힌두교 문화에서 신도들은 보통[133] 사원에 선물로 꽃을 가지고 옵니다.


재스민과 같은 꽃은 수세기 동안 중국에서 전통적인 차를 대체하는 것으로 사용되어 왔습니다. 가장 최근에 전세계에서 전통적으로 사용되는 많은 다른 허브와 꽃들은 다양한 꽃차를 준비하는 것에 중요성을 얻고 있습니다.[citation needed]
그래서 사람들은 집 주변에서 꽃을 기르거나, 거주 공간의 일부를 화원에 바치거나, 야생화를 따거나, 꽃집에서 상업적으로 재배된 꽃을 구입합니다. 꽃 생산 및 무역은 공정 무역 제품으로서의 가용성을 통해 개발도상국을 지원합니다.[134]

꽃은 다른 주요 식물 부분(종자, 열매, 뿌리, 줄기, 잎)에 비해 적은 식량을 제공하지만, 여전히 몇 가지 중요한 채소와 향신료를 제공합니다. 꽃 채소로는 브로콜리, 콜리플라워, 아티초크가 있습니다. 가장 비싼 향신료인 샤프란은 크로커스의 말린 스티그마로 이루어져 있습니다. 다른 꽃 향신료는 정향과 캐퍼입니다. 홉 꽃은 맥주 맛을 내기 위해 사용됩니다. 마리골드 꽃은 닭에게 먹여서 달걀 노른자에 소비자들이 더 바람직하다고 생각하는 황금색을 줍니다. 말린 마리골드 꽃은 조지아 요리에서 향신료와 착색제로 사용되기도 합니다. 민들레와 장로의 꽃은 종종 와인으로 만들어집니다. 벌에서 채취한 꽃가루인 벌 꽃가루는 어떤 사람들에게는 건강식품으로 여겨집니다. 꿀은 벌을 가공한 꽃꿀로 이루어져 있으며, 종종 꽃의 종류, 예를 들어 오렌지 꽃꿀, 클로버 꿀, 투펠로 꿀의 이름을 따서 붙여집니다.
수백 개의 신선한 꽃들이 먹을 수 있지만, 식품으로 널리 판매되는 것은 극소수에 불과합니다. 샐러드에 고명으로 첨가되는 경우가 많습니다. 호박꽃은 빵가루에 찍어 튀겨집니다. 일부 식용 꽃에는 나스투르티움, 국화, 카네이션, 캐테일, 일본의 꿀떡, 치커리, 옥수수꽃, 카나, 해바라기 등이 있습니다.[135] 데이지, 장미, 바이올렛과 같은 먹을 수 있는 꽃은 가끔 사탕에 담근다.[136]
향기와 약효로 선택된 국화, 장미, 재스민, 재스민, 캐모마일 등의 꽃은 차와 혼합하거나 자체적으로 티산으로 사용됩니다.[137]
꽃은 선사시대부터 장례 의식에 사용되어 왔습니다: 스페인 엘 미론 동굴에 있는 한 여성의 무덤에서 꽃가루의 흔적이 발견되었습니다.[138] 많은 문화들이 꽃과 삶과 죽음의 연관성을 그리고, 그들의 계절적인 귀환 꽃들 때문에 또한 부활을 암시하는데, 이것은 많은 사람들이 무덤에 꽃을 놓는 이유를 설명할지도 모릅니다. 에우리피데스의 희곡 페니키아 여인들에 기록된 바와 같이 고대 그리스인들은 고인의 머리 위에 꽃으로 된 왕관을 놓았고,[139] 그들은 또한 화환과 꽃잎으로 무덤을 덮었습니다. 꽃은 고대 이집트의 매장지에서 널리 사용되었고,[140] 오늘날까지 멕시코 사람들은 아즈텍 조상들이 그랬던 것과 같은 방식으로 죽은 자들의 날 기념식에서[141] 꽃을 두드러지게 사용합니다.

부여

꽃을 주는 전통은 꽃이 종종 약과 약초의 특성을 가지고 있었던 선사시대로 거슬러 올라갑니다. 고고학자들은 꽃잎의 잔재를 여러 무덤에서 발견했습니다. 꽃은 처음에 제물과 매장물로 사용되었습니다. 고대 이집트인들과 후에 그리스와 로마인들은 꽃을 사용했습니다. 이집트에서는 기원전[citation needed] 1540년경 붉은 양귀비, 노란 아라운, 콘플라워, 백합 등이 그려진 매장물이 발견됐습니다. 꽃을 준 기록은 중국의 글과 이집트의 상형문자는 물론 그리스 로마 신화에도 등장합니다. 꽃을 주는 행위는 중세에 부부가 꽃을 통해 애정을 드러내면서 번성했습니다.
꽃을 주는 전통은 다양한 형태로 존재합니다. 그것은 러시아 문화와 민속의 중요한 부분입니다. 학생들이 선생님께 꽃을 드리는 것은 흔한 일입니다. 열애 중 노란 꽃을 준다는 것은 러시아에서는 헤어짐을 의미합니다. 요즘은 꽃다발 형태로 꽃을 선물하는 경우가 많습니다.[142][143][144]
참고 항목
메모들
- ^ 그의 초기 작품들: Systema Naturae (1735)와 Genus plantarum (1737)도 이 분야에서 영향을 미쳤습니다.[117]
참고문헌
인용
- ^ "Plant Pollination Strategies". fs.usda.gov. 12 December 2022. Retrieved 16 February 2023.
- ^ "Pollinators in the Landscape II: Plants and Pollinators". ag.umass.edu. University of Massachusetts Amherst. 9 September 2015.
- ^ "Self-Pollination and Cross-Pollination Biology for Majors II". courses.lumenlearning.com. Archived from the original on 2020-07-21. Retrieved 2020-07-21.
- ^ Cresswell, Julia (2010). Oxford dictionary of word origins. Internet Archive. Oxford University Press. pp. 165 & 172. ISBN 978-0-19-954793-7.
- ^ "Definition of BLOSSOM". www.merriam-webster.com. Archived from the original on 2021-06-24. Retrieved 2021-06-16.
- ^ "Morphology Definition of Morphology by Oxford Dictionary on Lexico.com also meaning of Morphology". Lexico Dictionaries English. Archived from the original on March 5, 2020. Retrieved 2021-07-08.
- ^ De Craene & P. 2010, 4페이지
- ^ De Craene & P. 2010, 5-6쪽.
- ^ a b c De Craene & P. 2010, 7페이지
- ^ a b D. Mause 2016, 페이지 225.
- ^ a b D. Mause 2016, 페이지 226.
- ^ a b De Craene & P. 2010, 8페이지
- ^ 곤충-식물 생물학
- ^ D. Mause 2016, 페이지 229.
- ^ De Craene & P. 2010, 14페이지
- ^ a b Sattler, R. (1973). Organogenesis of Flowers. A Photographic Text-Atlas. University of Toronto Press. ISBN 978-0-8020-1864-9.
- ^ Endress, Peter K. (1996-07-25). Diversity and Evolutionary Biology of Tropical Flowers. Cambridge University Press. p. 11. ISBN 978-0-521-56510-3.
- ^ Reynolds, Joan; Tampion, John (1983). Double flowers: a scientific study. London: [Published for the] Polytechnic of Central London Press [by] Pembridge Press. p. 41. ISBN 978-0-86206-004-6.
- ^ De Craene & P. 2010, 25페이지
- ^ a b Weberling, Focko (1992). Morphology of Flowers and Inflorescences. Cambridge University Press. pp. 17–19. ISBN 0-521-25134-6.
- ^ D. Mause 2016, 페이지 243.
- ^ De Craene & P. 2010, 410쪽.
- ^ a b D. Mause 2016, 페이지 239.
- ^ De Craene & P. 2010, 21페이지
- ^ "Zantedeschia aethiopica". New Zealand Plant Conservation Network. Retrieved 2022-05-08.
- ^ D. Mause 2016, p. 228
- ^ De Craene & P. 2010, 22-24쪽.
- ^ Prenner, Gernard (February 2010). "Floral formulae updated for routine inclusion in formal taxonomic descriptions". Taxon. 59 (1): 241–250. doi:10.1002/tax.591022. Archived from the original on 2018-03-29.
- ^ De Craene & P. 2010, 38페이지
- ^ Stephen Downie; Ken Robertson. "Digital Flowers: Floral Formulas". University of Illinois. Archived from the original on 4 March 2016. Retrieved 28 January 2014.
- ^ "Plant Taxonomy - Biology 308". employees.csbsju.edu. Archived from the original on Jun 24, 2014.
- ^ Sharma 2009, pp. 165–166.
- ^ De Craene & P. 2010, 36쪽.
- ^ Eames, A.J. (1961). Morphology of the Angiosperms. New York: McGraw-Hill Book Co.
- ^ Leins, Peter (2010). Flower and fruit : morphology, ontogeny, phylogeny, function and ecology. Claudia Erbar. Stuttgart: Schweizerbart. ISBN 978-3-510-65261-7. OCLC 678542669.
- ^ Sattler, R. (1988). "A dynamic multidimensional approach to floral development". In Leins, P.; Tucker, S.C. & Endress, P.K. (eds.). Aspects of Floral Development. Berlin: J. Cramer/Borntraeger. pp. 1–6.
- ^ Sattler, R. & Jeune, B. (1992). "Multivariate analysis confirms the continuum view of plant form". Annals of Botany. 69 (3): 249–262. doi:10.1093/oxfordjournals.aob.a088338.
- ^ Ausín, I.; et al. (2005). "Environmental regulation of flowering". Int J Dev Biol. 49 (5–6): 689–705. doi:10.1387/ijdb.052022ia. PMID 16096975.
- ^ Turck, F.; Fornara, F.; Coupland, G. (2008). "Regulation and Identity of Florigen: Flowering Locus T Moves Centre Stage". Annual Review of Plant Biology. 59: 573–594. doi:10.1146/annurev.arplant.59.032607.092755. hdl:11858/00-001M-0000-0012-374F-8. PMID 18444908. S2CID 39798675. Archived from the original on 2019-12-15. Retrieved 2019-07-23.
- ^ Searle, I.; et al. (2006). "The transcription factor FLC confers a flowering response to vernalization by repressing meristem competence and systemic signaling in Arabidopsis". Genes Dev. 20 (7): 898–912. doi:10.1101/gad.373506. PMC 1472290. PMID 16600915.
- ^ D. Mause 2016, pp. 392–395.
- ^ Beekman, Madeleine; Nieuwenhuis, Bart; Ortiz-Barrientos, Daniel; Evans, Jonathan P. (2016). "Sexual selection in hermaphrodites, sperm and broadcast spawners, plants and fungi". Philosophical Transactions: Biological Sciences. 371 (1706): 5. doi:10.1098/rstb.2015.0541. ISSN 0962-8436. JSTOR 26143395. PMC 5031625. PMID 27619704.
The sole purpose of flowers is sex.
- ^ 워커 2020, 9페이지
- ^ a b c d D. Mause 2016, 페이지 238.
- ^ Turner, Vivienne (1984). "Banksia Pollen as a Source of Protein in the Diet of Two Australian Marsupials Cercartetus nanus and Tarsipes rostratus". Oikos. 43 (1): 53–61. doi:10.2307/3544245. ISSN 0030-1299. JSTOR 3544245. Archived from the original on 2021-06-24. Retrieved 2021-06-20 – via JSTOR.
[T]he honey possum, Tarsipes rostratus (Turner 1983). This marsupial is highly specialized for feeding at flowers and known to visit several species of Banksia
- ^ 워커 2020, 65쪽.
- ^ a b 워커 2020, 69~83쪽.
- ^ 워커 2020, 페이지 120.
- ^ Baker, Herbert G. (1983-01-01). "CHAPTER 2 - an Outline of the History of Anthecology, or Pollination Biology**Based on, but expanded and updated from, a paper published in the N.Z. J. Bot. (Baker, 1979), by permission of the editor". An Outline of the History of Anthecology, or Pollination Biology. p. 8. doi:10.1016/B978-0-12-583980-8.50009-0. ISBN 9780125839808. Archived from the original on 2021-06-24. Retrieved 2021-06-20 – via Elsevier Science Direct.
{{cite book}}:journal=무시됨(도움말) - ^ Friedman, Jannice (2011). "Gone with the wind: understanding evolutionary transitions between wind and animal pollination in the angiosperms". The New Phytologist. 191 (4): 911–913. doi:10.1111/j.1469-8137.2011.03845.x. ISSN 0028-646X. JSTOR 20869225. PMID 21834912.
- ^ a b c Ackerman, J. D. (2000-03-01). "Abiotic pollen and pollination: Ecological, functional, and evolutionary perspectives". Plant Systematics and Evolution. 222 (1): 167–185. doi:10.1007/BF00984101. ISSN 1615-6110. S2CID 36015720. Archived from the original on 2022-02-21. Retrieved 2021-06-20.
- ^ 워커 2020, 46쪽
- ^ D. Mause 2016, pp. 239–240.
- ^ 워커 2020, 68쪽.
- ^ 크누스, 뮐러 & 아인스워스 데이비스 1906, 페이지 72–80.
- ^ Vereecken, Nicolas J.; Wilson, Carol A.; Hötling, Susann; Schulz, Stefan; Banketov, Sergey A.; Mardulyn, Patrick (2012-12-07). "Pre-adaptations and the evolution of pollination by sexual deception: Cope's rule of specialization revisited". Proceedings of the Royal Society B: Biological Sciences. 279 (1748): 4786–4794. doi:10.1098/rspb.2012.1804. PMC 3497092. PMID 23055065.
- ^ 워커 2020, 81쪽.
- ^ 워커 2020, 112-113쪽.
- ^ Schäffler, Irmgard; Dötterl, Stefan (2011). "A day in the life of an oil bee: phenology, nesting, and foraging behavior" (PDF). Apidologie. 42 (3): 409–424. doi:10.1007/s13592-011-0010-3. S2CID 29392750.
- ^ a b 워커 2020, pp. 107–108.
- ^ 워커 2020, 페이지 121.
- ^ 워커 2020, 페이지 173
- ^ 거룩한 말하는 식물! 박쥐와 소통하는 꽃
- ^ 초음파 흡수, 박쥐를 꽃으로 유혹하는 에콰도르 선인장
- ^ a b D. Mause 2016, 페이지 241.
- ^ Peakall, Rod; Ebert, Daniel; Poldy, Jacqueline; Barrow, Russell A.; Francke, Wittko; Bower, Colin C.; Schiestl, Florian P. (2010). "Pollinator specificity, floral odour chemistry and the phylogeny of Australian sexually deceptive Chiloglottis orchids: implications for pollinator-driven speciation". New Phytologist. 188 (2): 437–450. doi:10.1111/j.1469-8137.2010.03308.x. ISSN 1469-8137. PMID 20561345.
- ^ D. Mause 2016, 페이지 240.
- ^ Bawa, K. S. (1990). "Plant-Pollinator Interactions in Tropical Rain Forests". Annual Review of Ecology and Systematics. 21: 415. doi:10.1146/annurev.es.21.110190.002151. ISSN 0066-4162. JSTOR 2097031. Archived from the original on 2021-06-25. Retrieved 2021-06-20 – via JSTOR.
First, the effect may be direct, as, for example, the loss of one of the interacting partners in species-specific interactions may lead to the extinction of the other.
- ^ Robledo‐Arnuncio, Juan José (April 2011). "Wind pollination over mesoscale distances: an investigation with Scots pine". New Phytologist. 190 (1): 222–233. doi:10.1111/j.1469-8137.2010.03588.x. ISSN 0028-646X. PMID 21175640.
- ^ Sofiev, M.; Siljamo, P.; Ranta, H.; Linkosalo, T.; Jaeger, S.; Rasmussen, A.; Rantio-Lehtimaki, A.; Severova, E.; Kukkonen, J. (2013-01-01). "A numerical model of birch pollen emission and dispersion in the atmosphere. Description of the emission module". International Journal of Biometeorology. 57 (1): 45–58. Bibcode:2013IJBm...57...45S. doi:10.1007/s00484-012-0532-z. ISSN 1432-1254. PMC 3527742. PMID 22410824. Archived from the original on 2022-02-21. Retrieved 2021-06-15.
- ^ a b 크누스, 뮐러 & 아인스워스 데이비스 1906, 68-72쪽.
- ^ Höcherl, Nicole; Siede, Reinhold; Illies, Ingrid; Gätschenberger, Heike; Tautz, Jürgen (2012-02-01). "Evaluation of the nutritive value of maize for honey bees". Journal of Insect Physiology. 58 (2): 278–285. doi:10.1016/j.jinsphys.2011.12.001. ISSN 0022-1910. PMID 22172382. Archived from the original on 2021-06-24. Retrieved 2021-06-20.
- ^ 워커 2020, 페이지 51.
- ^ 크누스, 뮐러 & 아인스워스 데이비스 1906, 68-69쪽.
- ^ a b c Cox, Paul Alan (1988-11-01). "Hydrophilous pollination". Annual Review of Ecology and Systematics. 19 (1): 261–279. doi:10.1146/annurev.es.19.110188.001401. ISSN 0066-4162. Archived from the original on 2021-06-24. Retrieved 2021-06-20.
- ^ 워커 2020, 36-37쪽.
- ^ D. Mause 2016, 페이지 222.
- ^ a b 워커 2020, 42쪽.
- ^ 찰스워스 D, 윌리스 JH. 근친상간 우울증의 유전자. Nat Rev Genet. 2009 Nov; 10(11):783-96. doi: 10.1038/nrg2664. PMID 19834483
- ^ 번스타인 H, 호프 FA, 미쇼드 RE. 성 진화의 분자적 기초. Adv Genet. 1987;24:323-70. doi: 10.1016/s0065-2660(08)60012-7. PMID 3324702
- ^ McPeek, Tamara M.; Wang, Xianzhong (2007). "Reproduction of Dandelion (Taraxacum officinale) in a Higher CO2 Environment". Weed Science. 55 (4): 334. doi:10.1614/WS-07-021. ISSN 0043-1745. JSTOR 4539580. S2CID 86250272. Archived from the original on 2021-06-25. Retrieved 2021-06-20 – via JSTOR.
- ^ Gilmartin, A. J.; Brown, Gregory K. (1985). "Cleistogamy in Tillandsia capillaris (Bromeliaceae)". Biotropica. 17 (3): 256. doi:10.2307/2388227. ISSN 0006-3606. JSTOR 2388227. Archived from the original on 2021-06-25. Retrieved 2021-06-20.
- ^ 워커 2020, 페이지 34.
- ^ Chen, Miaolin; Xu, Jie; Devis, Deborah; Shi, Jianxin; Ren, Kang; Searle, Iain; Zhang, Dabing (2016). "Origin and Functional Prediction of Pollen Allergens in Plants". Plant Physiology. 172 (1): 341–357. doi:10.1104/pp.16.00625. ISSN 0032-0889. PMC 5074609. PMID 27436829.
- ^ Constible, Juanita; Sandro, Luke; Lee, Richard E. (2008). "Ah-choo! Increased Risk of Pollen Allergies in the Northern Hemisphere". Journal of College Science Teaching. 37 (4): 82–89. ISSN 0047-231X. JSTOR 42992782. Archived from the original on 2021-06-25. Retrieved 2021-06-20.
- ^ a b D. Mause 2016, 페이지 234.
- ^ a b D. Mause 2016, pp. 235-237.
- ^ Wyman, Tarryn; Kelly, Dave (2017). "Quantifying seed dispersal by birds and possums in a lowland New Zealand forest". New Zealand Journal of Ecology. 41 (1). doi:10.20417/nzjecol.41.4. Archived from the original on 2021-06-30. Retrieved 2021-07-01.
- ^ Sipe, Timothy W.; Linnerooth, Amy R. (1995). "Intraspecific Variation in Samara Morphology and Flight Behavior in Acer saccharinum (Aceraceae)". American Journal of Botany. 82 (11): 1412–1419. doi:10.2307/2445868. ISSN 0002-9122. JSTOR 2445868. Archived from the original on 2021-07-09. Retrieved 2021-07-01.
- ^ Mount, Ann; Pickering, Catherine Marina (2009-10-01). "Testing the capacity of clothing to act as a vector for non-native seed in protected areas". Journal of Environmental Management. 91 (1): 168–179. doi:10.1016/j.jenvman.2009.08.002. hdl:10072/29656. ISSN 0301-4797. PMID 19717222. Archived from the original on 2021-06-29. Retrieved 2021-07-01.
- ^ D. Mause 2016, 페이지 248.
- ^ Bowler DE, Benton TG (May 2005). "Causes and consequences of animal dispersal strategies: relating individual behaviour to spatial dynamics". Biological Reviews of the Cambridge Philosophical Society. 80 (2): 205–25. doi:10.1017/S1464793104006645. PMID 15921049. S2CID 39351147.
- ^ a b Pijl 1972, p. 71.
- ^ a b Forget, Pierre-Michel (2005). Seed fate : predation, dispersal, and seedling establishment. Wallingford, Oxfordshire, UK: CABI Pub. p. 21. ISBN 0-85199-806-2. OCLC 54803650. Archived from the original on 2022-02-21. Retrieved 2021-07-01.
Seeds on plants can be dispersed via a variety of primary dispersal mechanisms, including abiotic factors, like wind or ballistic projection, or biotic factors, like fruit-eating birds.
- ^ a b c Norsworthy, Jason K.; Griffith, Griff; Griffin, Terry; Bagavathiannan, Muthukumar; Gbur, Edward E. (2014). "In-Field Movement of Glyphosate-Resistant Palmer Amaranth (Amaranthus palmen) and Its Impact on Cotton Lint Yield: Evidence Supporting a Zero-Threshold Strategy". Weed Science. 62 (2): 237–249. doi:10.1614/WS-D-13-00145.1. ISSN 0043-1745. JSTOR 43700653. S2CID 86013249. Archived from the original on 2021-07-09. Retrieved 2021-07-01 – via JSTOR.
Wind and water are common abiotic seed dispersal mechanisms, but there are several biotic dispersal mechanisms, such as movement via animals by adhesion (epizoochory) or ingestion (endozoochory), and even movement resulting from human activities (anthropochory).
- ^ a b D. Mause 2016, pp. 244–247.
- ^ Traveset, A.; Riera, N.; Mas, R. E. (2001). "Passage through bird guts causes interspecific differences in seed germination characteristics". Functional Ecology. 15 (5): 669–675. doi:10.1046/j.0269-8463.2001.00561.x. ISSN 1365-2435. Archived from the original on 2021-07-09. Retrieved 2021-07-01.
Many studies show that germination is more successful after seeds pass through the digestive tract of frugivores
- ^ Lengyel, Szabolcs; Gove, Aaron D.; Latimer, Andrew M.; Majer, Jonathan D.; Dunn, Robert R. (2010-02-20). "Convergent evolution of seed dispersal by ants, and phylogeny and biogeography in flowering plants: A global survey". Perspectives in Plant Ecology, Evolution and Systematics. 12 (1): 43–55. doi:10.1016/j.ppees.2009.08.001. ISSN 1433-8319. Archived from the original on 2021-07-02. Retrieved 2021-07-01.
- ^ Hämäläinen, Anni; Broadley, Kate; Droghini, Amanda; Haines, Jessica A.; Lamb, Clayton T.; Boutin, Stan; Gilbert, Sophie (2017). "The ecological significance of secondary seed dispersal by carnivores". Ecosphere. 8 (2): e01685. doi:10.1002/ecs2.1685. ISSN 2150-8925.
- ^ a b c d Howe, H. F. & Smallwood J. (1982). "Ecology of Seed Dispersal" (PDF). Annual Review of Ecology and Systematics. 13: 201–228. doi:10.1146/annurev.es.13.110182.001221. Archived from the original (PDF) on 2006-05-13.
- ^ Symon, D.E.; Whalen, M.A.; Mackay, D.A. (2000). "Diversity in Acaena (Rosaceae) in South Australia". Journal of the Adelaide Botanic Garden. 19: 55–73. ISSN 0313-4083. JSTOR 23874441. Archived from the original on 2021-06-28. Retrieved 2021-07-01 – via JSTOR.
- ^ a b Vittoz, Pascal; Engler, Robin (7 February 2008). "Seed dispersal distances: a typology based on dispersal modes and plant traits" (PDF). Botanica Helvetica. 117 (2): 109–124. doi:10.1007/s00035-007-0797-8. S2CID 2339616. Archived (PDF) from the original on 27 June 2021. Retrieved 1 July 2021.
- ^ Pijl 1972, pp. 24–25.
- ^ a b Hyslop, Jade; Trowsdale, Sam (2012). "A review of hydrochory (seed dispersal by water) with implications for riparian rehabilitation". Journal of Hydrology (New Zealand). 51 (2): 137–152. ISSN 0022-1708. JSTOR 43945038. Archived from the original on 2021-07-09. Retrieved 2021-07-01.
- ^ a b Feldkamp, Susan (2006). Modern Biology. United States: Holt, Rinehart, and Winston. p. 618.
- ^ Lewis, Susan K. (April 17, 2007). "Flowers Modern & Ancient". PBS Online. Archived from the original on September 7, 2009. Retrieved 2010-08-30.
- ^ NOVA. 2007-04-17. PBS. WGBH. First Flower.
- ^ Thomson, Helen (Aug 17, 2015). "Fossilised remains of world's oldest flower discovered in Spain". The Guardian. Archived from the original on 2017-03-03.
- ^ Chinese Academy of Sciences (18 December 2018). "Flowers originated 50 million years earlier than previously thought". EurekAlert!. Archived from the original on 18 December 2018. Retrieved 18 December 2018.
- ^ Soltis, Douglas E.; Soltis, Pamela S. (2004). "Amborella not a "basal angiosperm"? Not so fast". American Journal of Botany. 91 (6): 997–1001. doi:10.3732/ajb.91.6.997. PMID 21653455.
- ^ "South Pacific plant may be missing link in evolution of flowering plants". Eurekalert.org. 2006-05-17. Archived from the original on 2011-05-14. Retrieved 2010-08-30.
- ^ Gabbott, Sarah (1 August 2017). "Did the first flower look like this?". BBC News. Archived from the original on 1 August 2017. Retrieved 1 August 2017.
- ^ Sauquet, Hervé; et al. (1 August 2017). "The ancestral flower of angiosperms and its early diversification". Nature Communications. 8: 16047. Bibcode:2017NatCo...816047S. doi:10.1038/ncomms16047. PMC 5543309. PMID 28763051.
- ^ "Oily Fossils Provide Clues To The Evolution Of Flowers". Sciencedaily.com. 2001-04-05. Archived from the original on 2010-08-19. Retrieved 2010-08-30.
- ^ "Age-Old Question On Evolution Of Flowers Answered". Unisci.com. 2001-06-15. Archived from the original on 2010-06-10. Retrieved 2010-08-30.
- ^ May 2005, Robert Roy Britt 26 (2005-05-26). "Human Affection Altered Evolution of Flowers". livescience.com. Archived from the original on 2021-09-06. Retrieved 2021-09-06.
- ^ a b c d Sharma 2009, 21쪽.
- ^ Sharma 2009, 8쪽.
- ^ a b Sharma 2009, 10쪽.
- ^ a b Sharma 2009, 22쪽.
- ^ Sharma 2009, pp. 24-27.
- ^ a b Sharma 2009, 11쪽.
- ^ Sharma 2009, 96쪽.
- ^ "The Meanings of Flowers". Flower Magazine. 2018-05-28. Archived from the original on 2020-07-31. Retrieved 2018-12-12.
- ^ Audet, Marye. "Roses and Their Meaning". Archived from the original on 26 August 2014. Retrieved 22 August 2014.
- ^ Frownfelter, Andrea. "Flower Symbolism as Female Sexual Metaphor". Eastern Michigan University. Archived from the original on 2014-08-10.
{{cite journal}}: 저널 인용 요구사항journal=(도와주세요) - ^ Dee, Nerys (1995). Your dreams & what they mean: how to understand the secret language of sleep. London: Thornsons. p. 142. ISBN 978-0-7225-3218-8. OCLC 33208041. Archived from the original on 2022-02-21. Retrieved 2021-06-15.
- ^ "Vishnu". Bbc.co.uk. 2009-08-24. Archived from the original on 2010-10-28. Retrieved 2010-08-30.
- ^ "God's Favorite Flower". Hinduism Today. Archived from the original on 2009-04-13. Retrieved 2010-08-30.
- ^ "The Lotus". Theosociety.org. Archived from the original on 2017-09-10. Retrieved 2010-08-30.
- ^ Fogden, Michael; Fogden, Patricia (2018). The Natural History of Flowers. Texas A&M University Press. p. 1. ISBN 978-1623496449.
- ^ Wilson, Adelaide B (1967). Flowers for your church. Place of publication not identified: Barrows & Co. p. 26. OCLC 500430357. Archived from the original on 2021-06-28. Retrieved 2021-06-15.
- ^ "VISITING A HINDU TEMPLE; A BEGINNER'S GUIDE". Hinduism Today. April 1991. Archived from the original on 2021-04-16. Retrieved 2018-11-27.
- ^ Fairtrade Foundation, 화훼농가 및 근로자, 2023년 3월 23일 접속
- ^ Wood, Zoe (30 June 2017). "Blooming tasty – edible flowers are summer's hottest food trend". The Guardian. Archived from the original on 27 November 2018. Retrieved 27 November 2018.
- ^ Bradley, Sue (8 May 2015). "How to crystallise flowers like the Victorians". The Telegraph. Archived from the original on 2022-01-11.
- ^ Wong, James (7 May 2017). "Grow your own herbal teas". The Guardian. Archived from the original on 27 November 2018. Retrieved 27 November 2018.
- ^ "Stone Age mourners 'placed flowers on graves'". The Telegraph. 10 May 2015. Archived from the original on 2022-01-11.
- ^ "Burial Rites". Hellenica World. Archived from the original on 29 November 2018. Retrieved 28 November 2018.
- ^ Hays, Christopher B. (2011). Death in the Iron Age II and in First Isaiah. Mohr Siebeck. p. 302. ISBN 978-3-16-150785-4. Archived from the original on 2019-12-28. Retrieved 2018-11-28.
- ^ Day, Frances Ann (2003). Latina and Latino Voices in Literature. Greenwood. p. 72. ISBN 978-0-313-32394-2.
- ^ By (2015-08-11). "The Fascinating Tradition of Giving Flowers". Flowers of the Field Las Vegas. Archived from the original on 2019-03-16. Retrieved 2019-03-15.
- ^ "The Cross-Cultural Rhetoric Blog: Flowers in Russia". web.stanford.edu. Archived from the original on 2019-04-13. Retrieved 2019-03-15.
- ^ "Folk Art in Russia and Ukraine News & Info". Archived from the original on 2019-03-24. Retrieved 2019-03-15.
원천
- De Craene, Ronse; P., Louis (2010). Floral Diagrams. Cambridge: Cambridge University Press. doi:10.1017/cbo9780511806711. ISBN 978-0-511-80671-1.
- Knuth, Paul; Müller, Hermann; Ainsworth Davis, J. R. (1906). Handbook of flower pollination: based upon Hermann Müller's work 'The fertilisation of flowers by insects'. Vol. 1. Clarendon Press. doi:10.5962/bhl.title.54973. OCLC 1841036. Archived from the original on 2022-02-21. Retrieved 2021-06-15.
- Fritsch, Felix Eugene; Salisbury, E. J. (Edward James) (1920). An introduction to the structure and reproduction of plants. Cornell University Library. London: G. Bell and Sons Ltd.
- Walker, Timothy (2020). Pollination: The Enduring Relationship Between Plant and Pollinator. Princeton University Press. ISBN 978-0-691-20375-1.
- D. Mauseth, James (2016). Botany: An Introduction to Plant Biology (6th ed.). Jones & Bartlett Learning. ISBN 978-1-284-07753-7.
- Pijl, L. van der (1972). Principles of Dispersal in Higher Plants (2nf ed.). Berlin Heidelberg: Springer-Verlag. ISBN 978-3-642-96108-3. Archived from the original on 2021-07-09. Retrieved 2021-07-01.
- Sharma, O. P. (2009). Plant Taxonomy (2nd ed.). New Delhi, India: Tata McGraw Hill Education Private Limited. ISBN 978-0070141599.
더보기
- Buchmann, Stephen (2016). The Reason for Flowers: Their History, Culture, Biology, and How They Change Our Lives. Scribner. ISBN 978-1-4767-5553-3.
- Esau, Katherine (1965). Plant Anatomy (2nd ed.). New York: John Wiley & Sons. ISBN 978-0-471-24455-4.
- Greyson, R.I. (1994). The Development of Flowers. Oxford University Press. ISBN 978-0-19-506688-3.
- Leins, P. & Erbar, C. (2010). Flower and Fruit. Stuttgart: Schweizerbart Science Publishers. ISBN 978-3-510-65261-7.
- Sattler, R. (1973). Organogenesis of Flowers. A Photographic Text-Atlas. University of Toronto Press. ISBN 978-0-8020-1864-9.
외부 링크
 Wiki 인용문에서 꽃과 관련된 인용문
Wiki 인용문에서 꽃과 관련된 인용문- 네이티브 플랜트 정보망


