에너지 흐름(에콜로지)
Energy flow (ecology)에너지 흐름은 생태계 [1]내의 생물들을 통한 에너지의 흐름이다.모든 생물은 생산자와 소비자로 구성될 수 있고, 그 생산자와 소비자는 먹이사슬로 [2][3]구성될 수 있다.먹이사슬 내의 각 레벨은 영양 [1]레벨입니다.각 영양 수준에서 유기체의 양을 보다 효율적으로 보여주기 위해, 이러한 먹이 사슬은 영양 [1]피라미드로 조직된다.먹이사슬의 화살표는 에너지 흐름의 방향을 나타내는 화살촉과 함께 에너지 흐름이 단방향임을 보여줍니다;[2][3] 에너지는 도중에 각 단계에서 열로 손실됩니다.
에너지의 단방향 흐름과 먹이 거미줄을 타고 올라갈 때의 연속적인 에너지 손실은 열역학의 지배를 받는 에너지 흐름의 패턴으로,[4][5] 시스템 간의 에너지 교환 이론입니다.영양역학은 [1]열역학(태양에서 태양 복사를 통해 외부에서 발생하는)의 유기체 간 전달과 변환을 다루기 때문에 열역학과 관련이 있습니다.

에너지학과 탄소 순환

에너지학의 첫 번째 단계는 광합성이며, 이 광합성은 공기 중의 물과 이산화탄소가 태양으로부터 온 에너지로 흡수되어 [7]산소와 포도당으로 전환된다.세포 호흡은 산소와 당이 흡수되어 이산화탄소와 물로 다시 전환되면서 에너지를 방출하는 역반응이다.호흡에 의해 생성된 이산화탄소와 물은 식물에 재활용될 수 있다.
에너지 손실은 효율성(에너지가 다음 단계로 얼마나 이동하는지) 또는 바이오매스(정립 [1]작물로 측정한 한 시점에 해당 수준에 존재하는 생물 물질의 양)로 측정할 수 있습니다.생산자 영양 수준에서의 모든 순 1차 생산량 중 일반적으로 10%만이 다음 단계인 1차 소비자에게 전달되고, 그 10%만이 다음 영양 수준까지 전달되며, 식품 [1]피라미드에서도 마찬가지입니다.생태계의 [8][1]효율성 또는 비효율성에 따라 생태계의 효율은 5%에서 20%가 될 수 있습니다.이러한 효율의 감소는 유기체가 생존하기 위해 세포호흡을 해야 하고 세포호흡을 할 [1]때 에너지가 열로 손실되기 때문에 발생한다.그것이 또한 [1]생산자보다 3차 소비자가 적은 이유이기도 하다.
프라이머리 생산
생산자는 광합성을 [9]하는 유기체이다.생산자들은 태양으로부터 에너지를 저장 가능하고 사용 가능한 에너지, 포도당,[1] 그리고 산소의 화학적인 형태로 바꾸기 때문에 중요하다.생산자 스스로 포도당에 저장된 에너지를 세포 호흡을 위해 사용할 수 있습니다.또는 생산자가 다음 영양 수준에서 초식동물에 의해 소비된다면, 에너지의 일부는 피라미드 [1]위로 전달된다.생산자 안에 저장된 포도당은 소비자에게 음식으로 작용하기 때문에 소비자가 태양 에너지에 [1][7]접근할 수 있는 것은 생산자를 통해서만 가능하다.주요 생산자의 예로는 조류, 이끼, 그리고 풀, 나무, [1]관목과 같은 다른 식물들이 있다.
화학합성 박테리아는 광합성과 비슷한 과정을 수행하지만 태양 에너지 대신 [10][11]황화수소와 같은 화학 물질에 저장된 에너지를 사용한다.화학합성이라고 불리는 이 과정은 보통 열과 수소, 황화수소, [10]메탄과 같은 화학물질을 생산하는 열수 분출구의 깊은 바다 속에서 일어난다.화학합성 박테리아는 황화수소와 산소의 결합에 있는 에너지를 이산화탄소를 포도당으로 전환시켜 그 과정에서 물과 황을 방출한다.[11]화학 합성 박테리아를 소비하는 유기체는 포도당을 흡수하고 산소를 사용하여 세포 호흡을 수행할 수 있는데, 이는 초식동물이 생산자를 소비하는 것과 유사합니다.
1차 생산을 제어하는 요인 중 하나는 생산자로 유입되는 에너지의 양이며,[12][13][1] 이는 생산성을 사용하여 측정할 수 있습니다.태양 에너지의 단 1%만이 생산자로 들어가고 나머지는 튕겨나가거나 [13]통과한다.1차 총생산성은 생산자가 실제로 [13][14]얻는 에너지의 양입니다.일반적으로 생산자에게 유입되는 에너지의 60%는 생산자의 [12]호흡에 사용됩니다.순 1차 생산성은 식물이 세포 호흡에 사용한 양을 [13]뺀 후 유지되는 양이다.1차 생산을 제어하는 또 다른 요인은 생산자가 [14]살고 있는 물이나 토양에서 유기/무기 영양소의 수준입니다.
이차 생산
2차 생산은 소비자가 자체 바이오매스로 전환한 식물에 저장된 에너지를 사용하는 것이다.에코시스템에 따라 소비자 수준이 다르며, 결국 하나의 상위 소비자로 끝납니다.대부분의 에너지는 식물의 유기물에 저장되며, 소비자는 식물을 먹을 때 이 에너지를 소비한다.초식동물과 잡식동물의 이 에너지는 육식동물에 의해 소비된다.또한 1차 생산 중이고 결국 쓰레기와 쓰레기로 끝나는 많은 양의 에너지가 있습니다.유해 먹이사슬에는 다량의 미생물, 대식충 동물, 마이오파우나, 곰팡이, 그리고 박테리아가 포함되어 있다.이러한 유기체는 잡식동물과 육식동물에 의해 소비되며 많은 양의 2차 생산을 [15]차지한다.2차 소비자는 소비 [16]효율에 있어 크게 다를 수 있습니다.소비자에게 전달되는 에너지 효율은 약 10%[16]로 추정됩니다.소비자를 통한 에너지 흐름은 수생 및 육상 환경에 따라 다르다.
수중 환경에서의 2차 생산
헤테로트로프는 2차 생산에 기여하며 1차 생산성과 순 1차 [16]생산물에 의존합니다.2차 생산은 초식동물과 분해자가 사용하는 에너지로 1차 [16]생산성에 의존합니다.주로 초식동물과 분해자는 수생 생태계의 두 가지 주요 유기원, 즉 자기와 [16]동종에서 나오는 모든 탄소를 소비한다.오토콘 탄소는 생태계 내에서 나오며 수생식물, 조류, 식물성 플랑크톤을 포함한다.생태계 외부에서 유입되는 알로치톤성 탄소는 대부분 물로 [16]유입되는 육상 생태계의 죽은 유기물이다.스트림 생태계에서는 연간 에너지 투입량의 약 66%를 다운스트림으로 방출할 수 있습니다.나머지 양은 [17]열로 소비되고 손실됩니다.
지상 환경에서의 2차 생산
이차 생산은 종종 영양 수준의 관점에서 설명되며, 이것은 관계를 설명하는 데 유용할 수 있지만, 더 드문 상호작용을 지나치게 강조한다.소비자들은 종종 여러 [18]영양 수준에서 음식을 먹는다.세 번째 영양 수준 이상으로 전달되는 에너지는 상대적으로 [18]중요하지 않습니다.동화효율은 소비자가 섭취한 음식의 양, 동화되는 양, 대변이나 소변으로 배출되는 양으로 표현될 수 있다.에너지의 일부는 호흡에 사용되는 반면, 에너지의 다른 일부는 전기 [16]소비 장치의 바이오매스로 사용됩니다.먹이사슬은 크게 두 가지가 있습니다.1차 먹이사슬은 자가영양에서 나와 소비자에게 전달되는 에너지이고, 2차 먹이사슬은 육식동물이 자기영양 [16]에너지를 소비하는 초식동물이나 분해체를 먹는 것입니다.소비자는 1차 소비자와 2차 소비자와 3차 소비자로 나뉜다.육식동물은 에너지 동화가 훨씬 더 높고, 약 80%이며, 초식동물은 약 20-50%[16]의 훨씬 낮은 효율을 가지고 있습니다.시스템의 에너지는 동물의 이주/이주에 의해 영향을 받을 수 있습니다.생물들의 움직임은 육지 [17]생태계에서 중요하다.육생 생태계에서 초식동물의 에너지 소비는 3~7%[17]로 낮다.에너지의 흐름은 많은 지상 환경에서 비슷하다.초식동물의 순 1차 소비량 변동은 일반적으로 낮다.이는 방목민의 소비가 약 33%[17]로 훨씬 높은 호수 및 연못의 수중 환경과 큰 대조를 이룬다.외부온도와 내부온도는 매우 다른 [16]동화효율을 가지고 있다.
유해동물
유해동물은 부패하고 있는 유기물을 섭취하고 육식동물에 [16]의해 소비된다.포식자의 생산성은 먹잇감의 생산성과 관련이 있습니다.이를 통해 생태계의 주요 생산성이 [19]이후의 모든 생산성에 영향을 미친다는 것을 확인할 수 있습니다.
이물질은 생태계에서 유기물질의 많은 부분을 차지한다.온대 숲의 유기 물질은 대부분 죽은 식물들로 구성되어 있으며, 약 62%[18]입니다.
수생태계에서는 하천에 떨어진 잎사귀가 젖어 유기물이 거머리처럼 움직이기 시작한다.이것은 다소 빠르게 일어나며 미생물들과 무척추동물을 유인할 것이다.그 잎들은 거친 입자 [15]유기물이라고 불리는 큰 조각들로 분해될 수 있다.CPOM은 미생물에 의해 빠르게 서식하고 있다.Meiofauna는 하천 [15]생태계에서 2차 생산에 매우 중요합니다.이 잎을 분해하고 군집화하는 미생물은 유해생물에게 매우 중요하다.디테리토보어는 조직으로부터 화합물을 방출함으로써 잎을 더 먹을 수 있게 만든다; 그것은 궁극적으로 [15]잎을 부드럽게 하는데 도움을 준다.잎의 셀룰로오스와 리그닌이 분해되기 어렵기 때문에 질소가 부패하기 때문에 질소가 감소합니다.따라서, 군집화 미생물은 분해를 돕기 위해 질소를 들여온다.잎의 분해는 초기 질소 함량, 계절, 그리고 나무의 종류에 따라 달라질 수 있습니다.나뭇잎이 떨어질 때 나무의 종류는 다양할 수 있다.따라서 잎의 분해는 미생물 집단의 [15]모자이크라고 불리는 다른 시기에 일어난다.
생태계의 종 효과와 다양성은 그 성능과 [20]효율성을 통해 분석될 수 있습니다.또한 하천의 2차 생산은 하천으로 떨어지는 이물질의 영향을 크게 받을 수 있다. 즉, 쓰레기 제거 및 [19]배제에 대한 연구 중에 해저 동물군 바이오매스의 생산과 풍부성이 47-50% 감소하였다.
생태계 전체의 에너지 흐름
연구에 따르면 1차 생산자들은 생태계 전체에서 [14]비슷한 비율로 탄소를 고정하는 것으로 나타났습니다.일단 탄소가 실행 가능한 에너지원으로 시스템에 도입되면, 더 높은 영양 수준으로 에너지의 흐름을 제어하는 메커니즘은 생태계에 따라 달라집니다.수생 및 육생 생태계 중에서 이러한 변동을 설명할 수 있는 패턴이 확인되었으며, 두 가지 주요 제어 경로인 하향식 [21][22]및 상향식으로 구분되었다.각 경로 내의 작용 메커니즘은 궁극적으로 생태계 내의 군집과 영양 수준 구조를 다양한 정도로 [23]조절합니다.상향식 제어에는 자원 품질과 가용성에 기초한 메커니즘이 포함됩니다.이 메커니즘은 1차 생산성과 그에 따른 에너지 및 바이오매스의 흐름을 제어하여 영양 [22]수준을 높입니다.하향식 제어에는 [23][22]소비자의 소비에 기반한 메커니즘이 포함됩니다.이러한 메커니즘은 초식동물이나 포식자들이 더 낮은 영양 [21]레벨을 먹고 살기 때문에 한 영양 레벨에서 다른 영양 레벨로 에너지가 전달되는 속도를 조절합니다.
수생생태계와 육생태계
각 유형의 생태계 내에서 에너지 흐름의 많은 변화가 발견되어 생태계 유형 간의 변화를 식별하기가 어렵습니다.일반적으로 에너지 흐름은 온도, 물 가용성 및 빛 [24]가용성과 함께 일차 생산성의 함수입니다.예를 들어, 수생 생태계 중에서, 깊은 호수나 맑은 [24]물줄기보다 큰 강과 얕은 호수에서 더 높은 생산률을 볼 수 있다.육지 생태계 중 습지, 늪지, 열대우림이 1차 생산률이 가장 높은 반면 툰드라, 고산 생태계는 [24]가장 낮다.1차 생산과 환경 조건 간의 관계는 생태계 유형 내의 변화를 설명하는데 도움이 되었으며, 생태학자들은 다양한 상향식 및 하향식 제어가 [22]작용하기 때문에 에너지가 지상 생태계보다 수생 생태계를 통해 더 효율적으로 흐른다는 것을 입증할 수 있었다.
상향식
에너지 흐름에 대한 상향식 제어의 강도는 생태계 [14][21]내 1차 생산자의 영양 품질, 크기 및 성장률에 따라 결정됩니다.광합성 물질은 전형적으로 질소(N)와 인(P)이 풍부하고 모든 [25]생태계에 걸쳐 N과 P에 대한 높은 초식동물 수요를 보충합니다.수생 1차 생산은 주로 광합성 물질로 구성된 작은 단세포 식물 플랑크톤에 의해 지배되며,[21] 초식동물들에게 이러한 영양소의 효율적인 원천을 제공합니다.이와는 대조적으로, 다세포 육생 식물은 탄소수는 높지만 영양가는 [21]낮은 많은 큰 지지 셀룰로오스 구조를 가지고 있다.이러한 구조적 차이 때문에, 수생 1차 생산자들은 육생 생태계의 [21]숲과 초원보다 수생 생태계에 저장된 광합성 조직당 바이오매스가 적다.수생 생태계의 광합성 물질에 비해 바이오매스가 낮기 때문에 육상 [21]생태계에 비해 회전율이 더 효율적입니다.식물성 플랑크톤이 초식동물에 의해 소비됨에 따라, 그들의 향상된 성장과 번식 속도는 손실된 바이오매스를 충분히 대체하고 영양소 밀도가 높은 품질과 함께 더 큰 2차 [21]생산을 지원합니다.
1차 생산에 영향을 미치는 추가 요인에는 수생 [21]생태계에서 더 큰 규모로 발생하는 N과 P의 투입이 포함된다.이 영양소들은 식물의 성장을 촉진하는 데 중요하며, 더 높은 영양 수준으로 전달되면, 소비자의 [22][24]바이오매스와 성장률을 높입니다.만약 이 영양소들 중 하나가 부족하면, 그것들은 전반적인 1차 [15]생산을 제한할 수 있다.호수 내에서는 P가 더 제한적인 영양소인 반면, N과 P는 모두 [22]강에서 1차 생산을 제한한다.이러한 제한 효과로 인해 영양소 투입은 잠재적으로 수생 [23]생태계의 순 1차 생산 제한을 완화할 수 있다.수생생태계에 씻긴 알로치톤성 물질은 N, P와 더불어 1차 [15]생산자가 쉽게 흡수하는 탄소 분자 형태의 에너지를 도입한다.투입량이 많아지고 영양소 농도가 높아지면 1차 순생산률이 높아져 2차 [25]생산량이 많아진다.
톱다운
하향식 메커니즘은 수생 식품 [23]거미줄 내의 소비자 이동으로 인해 수생 1차 생산자를 더 잘 통제한다.소비자들 사이에서, 초식동물들은 1차 생산자에서 더 높은 영양 [26]레벨의 포식자로의 에너지 흐름을 연결함으로써 영양 계단식의 영향을 중재할 수 있다.생태계 전반에 걸쳐 초식동물 성장과 생산자 영양 [25]품질 사이에는 일관된 연관성이 있습니다.그러나 수생 생태계에서 1차 생산자는 육생 [21]생태계보다 4배 빠른 속도로 초식동물에 의해 소비된다.비록 이 주제가 매우 논란이 되고 있지만, 연구원들은 초식동물 조절의 차이를 생산자 대 소비자 크기 비율과 초식동물 [7]선택성을 포함한 여러 이론의 탓으로 돌렸다.
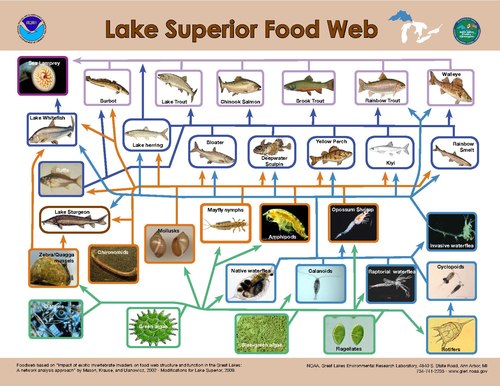
1차 생산자에 대한 하향식 제어 모델링은 1차 생산자에 대한 소비자 대 1차 생산자의 크기 비율이 가장 [28]높을 때 에너지 흐름에 대한 가장 큰 제어가 발생함을 시사한다.수생계에서 단일 영양 수준 내에서 발견되는 유기체의 크기 분포는 [21]육생계보다 훨씬 좁다.육지에서는 곤충과 같이 소비하는 식물보다 작은 소비자 크기부터 유제류처럼 상당히 큰 소비자 크기까지 다양하지만, 수생 시스템에서는 영양 수준 내의 소비자 신체 크기가 훨씬 덜 달라지고 영양적 [21]위치와 강한 상관 관계가 있다.그 결과 생산자와 소비자의 크기 차이는 육지보다 수생 환경에서 일관되게 커지기 때문에 수생 1차 [21]생산자에 대한 초식동물 방제가 강화된다.
초식동물들은 유기물이 먹이망을 [26]통해 순환되면서 잠재적으로 유기물의 운명을 통제할 수 있다.초식동물들은 구조적 방어기구가 [21]있는 식물을 피하면서 영양분이 풍부한 식물을 선택하는 경향이 있다.지지구조와 마찬가지로 방어구조는 영양소 부족, 고탄소 [26]셀룰로오스로 구성됩니다.영양가 있는 식품원에 대한 접근은 초식 대사와 에너지 수요를 증가시켜 1차 [14]생산자의 이동을 증가시킨다.수생 생태계에서 식물성 플랑크톤은 영양가가 높고 일반적으로 방어 메커니즘이 [26]부족하다.이렇게 하면 소비된 식물 물질이 불안정한 유기 [15][26]폐기물로 빠르게 시스템으로 다시 배출되기 때문에 하향식 제어가 강화됩니다.지상 생태계에서 1차 생산자는 영양학적으로 밀도가 낮고 방어 [21]구조를 포함할 가능성이 높다.초식동물들은 영양학적으로 밀도가 높은 식물을 선호하고 방어구조가 있는 식물이나 식물의 일부를 피하기 때문에,[26] 생태계 내에서 더 많은 양의 식물 물질이 소비되지 않고 남겨진다.초식동물의 질 낮은 식물 물질 회피는 지상 시스템이 [21]에너지 흐름에 대한 하향식 제어가 약한 이유일 수 있다.
「 」를 참조해 주세요.
레퍼런스
- ^ a b c d e f g h i j k l m n Lindeman RL (1942). "The Trophic-Dynamic Aspect of Ecology" (PDF). Ecology. 23 (4): 399–417. doi:10.2307/1930126. JSTOR 1930126. Archived from the original (PDF) on 2017-03-29. Retrieved 2020-12-04.
- ^ a b Briand F, Cohen JE (November 1987). "Environmental correlates of food chain length". Science. New York, N.Y. 238 (4829): 956–60. Bibcode:1987Sci...238..956B. doi:10.1126/science.3672136. PMID 3672136.
- ^ a b Vander Zanden MJ, Shuter BJ, Lester N, Rasmussen JB (October 1999). "Patterns of Food Chain Length in Lakes: A Stable Isotope Study". The American Naturalist. 154 (4): 406–416. doi:10.1086/303250. PMID 10523487. S2CID 4424697.
- ^ Sharma JP (2009). Environmental studies (3rd ed.). New Delhi: University Science Press. ISBN 978-81-318-0641-8. OCLC 908431622.
- ^ Van Ness HC (1969). Understanding thermodynamics (Dover ed.). New York: Dover Publications, Inc. ISBN 978-1-62198-625-6. OCLC 849744641.
- ^ "Carbon Cycle". Archived from the original on 12 August 2006.
- ^ a b c d Whitmarsh JGovindjee(1999년)."그 광합성 과정".Singhal GS, Renger G, Sopory SK, Irrgang의 KD, Govindjee(eds.)에서.광생물학의 개념:광합성과 광형 태형성.보스턴:Kluwer 학술 출판사.를 대신하여 서명함. 11–51.아이 에스비엔 978-0-7923-5519-9..carbon/year의 4×1018 kJ/yr에 해당합니다 100×1015그램 광합성 생물에 의해 고정되어, 음 4×1021 J/yr 자유 에너지 감소 탄소 저장.
- ^ Teal JM (1962). "Energy flow in the salt marsh ecosystem of Georgia". Ecology. 43 (4): 614–624. doi:10.2307/1933451. JSTOR 1933451.
- ^ Morris J, Hartl DL, Knoll AH, Lue R, Michael M (2019). Biology: How Life Works (3rd ed.). W. H. Freeman. ISBN 978-1319017637.
- ^ a b Kellermann MY, Wegener G, Elvert M, Yoshinaga MY, Lin YS, Holler T, Mollar XP, Knittel K, Hinrichs KU (November 2012). "Autotrophy as a predominant mode of carbon fixation in anaerobic methane-oxidizing microbial communities". Proceedings of the National Academy of Sciences of the United States of America. 109 (47): 19321–6. Bibcode:2012PNAS..10919321K. doi:10.1073/pnas.1208795109. PMC 3511159. PMID 23129626.
- ^ a b Cavanaugh CM, Gardiner SL, Jones ML, Jannasch HW, Waterbury JB (July 1981). "Prokaryotic Cells in the Hydrothermal Vent Tube Worm Riftia pachyptila Jones: Possible Chemoautotrophic Symbionts". Science. New York, N.Y. 213 (4505): 340–2. Bibcode:1981Sci...213..340C. doi:10.1126/science.213.4505.340. PMID 17819907.
- ^ a b Amthor JS, Baldocchi DD (2001). "Terrestrial higher plant respiration and net primary production". Terrestrial Global Productivity: 33–59. doi:10.1016/B978-012505290-0/50004-1. ISBN 9780125052900.
- ^ a b c d Sigman DM, Hain MP (2012). "The biological productivity of the ocean" (PDF). Nature Education Knowledge. 3 (6): 1–6.
- ^ a b c d e Cebrian J (October 1999). "Patterns in the Fate of Production in Plant Communities". The American Naturalist. 154 (4): 449–468. doi:10.1086/303244. PMID 10523491. S2CID 4384243.
- ^ a b c d e f g h Allan JD, Castillo MM (2007). Stream ecology: structure and function of running waters (2nd ed.). Dordrecht: Springer. ISBN 978-1-4020-5582-9. OCLC 144222191.
- ^ a b c d e f g h i j k Smith TM, Smith RL (2015). Elements of ecology (9th ed.). Boston. ISBN 978-1-292-07741-3. OCLC 914328590.
- ^ a b c d Fisher SG, Likens GE (February 1973). "Energy Flow in Bear Brook, New Hampshire: An Integrative Approach to Stream Ecosystem Metabolism". Ecological Monographs. 43 (4): 421–439. doi:10.2307/1942301. JSTOR 1942301.
- ^ a b c Hairston Jr NG, Hairston Sr NG (September 1993). "Cause-Effect Relationships in Energy Flow, Trophic Structure, and Interspecific Interactions". The American Naturalist. 142 (3): 379–411. doi:10.1086/285546. hdl:1813/57238. S2CID 55279332.
- ^ a b Wallace JB, Eggert SL, Meyer JL, Webster JR (November 1999). "Effects of resource limitation on a detrital‐based ecosystem". Ecological Monographs. 69 (4): 409–42. doi:10.1890/0012-9615(1999)069[0409:eorloa]2.0.co;2.
- ^ Cardinale BJ, Srivastava DS, Duffy JE, Wright JP, Downing AL, Sankaran M, Jouseau C (October 2006). "Effects of biodiversity on the functioning of trophic groups and ecosystems". Nature. 443 (7114): 989–92. Bibcode:2006Natur.443..989C. doi:10.1038/nature05202. PMID 17066035. S2CID 4426751.
- ^ a b c d e f g h i j k l m n o p Shurin JB, Gruner DS, Hillebrand H (January 2006). "All wet or dried up? Real differences between aquatic and terrestrial food webs". Proceedings. Biological Sciences. 273 (1582): 1–9. doi:10.1098/rspb.2005.3377. PMC 1560001. PMID 16519227.
- ^ a b c d e f La Pierre K, Hanley T (2015). Trophic Ecology: Bottom-Up and Top-Down Interactions Across Aquatic and Terrestrial Systems. Cambridge University Press. pp. 55–85. ISBN 9781316299692.
- ^ a b c d Gruner DS, Smith JE, Seabloom EW, Sandin SA, Ngai JT, Hillebrand H, et al. (July 2008). "A cross-system synthesis of consumer and nutrient resource control on producer biomass". Ecology Letters. 11 (7): 740–55. doi:10.1111/j.1461-0248.2008.01192.x. PMID 18445030.
- ^ a b c d Ricklefs RE, Miller GL (2000). Ecology (4th ed.). New York: W.H. Freeman & Co. ISBN 0-7167-2829-X. OCLC 40734932.
- ^ a b c Cebrian J, Lartigue J (2004). "Patterns of Herbivory and Decomposition in Aquatic and Terrestrial Ecosystems". Ecological Monographs. 74 (2): 237–259. doi:10.1890/03-4019.
- ^ a b c d e f Schmitz OJ (December 2008). "Herbivory from Individuals to Ecosystems". Annual Review of Ecology, Evolution, and Systematics. 39 (1): 133–152. doi:10.1146/annurev.ecolsys.39.110707.173418.
- ^ Krause AE, Frank KA, Mason DM, Ulanowicz RE, Taylor WW (November 2003). "Compartments revealed in food-web structure". Nature. 426 (6964): 282–5. Bibcode:2003Natur.426..282K. doi:10.1038/nature02115. hdl:2027.42/62960. PMID 14628050. S2CID 1752696.
- ^ Shurin JB, Seabloom EW (2005). "The strength of trophic cascades across ecosystems: predictions from allometry and energetics". Journal of Animal Ecology. 74 (6): 1029–1038. doi:10.1111/j.1365-2656.2005.00999.x. ISSN 1365-2656.
추가 정보
- Podolinsky S (2004). "Socialism and the Unity of Physical Forces". Organization & Environment. 17 (1): 61–75. doi:10.1177/1086026603262092. S2CID 144332347.
- Weiner DR (2000). Models of Nature: Ecology, Conservation and Cultural Revolution in Soviet Russia. U.S.: University of Pittsburgh Press.



{kind=link}