물고기 비늘
Fish scale
물고기 비늘은 물고기의 껍질에서 자라는 작고 단단한 판이다. 대부분의 물고기의 피부는 이러한 보호 비늘로 덮여 있는데, 이것은 또한 반사 및 착색 사용을 통해 효과적인 위장 기능을 제공할 수 있을 뿐만 아니라 가능한 유체역학적인 장점도 제공할 수 있다. 저울이라는 용어는 조개껍질 꼬투리나 껍데기를 뜻하는 구 프랑스어 "에스카일"에서 유래되었다.[1]
비늘은 새우잡이, 상어 등 어류의 강하고 단단한 갑옷 판에서부터 장어, 낚시어 등 어류의 미세하거나 없는 것까지 크기, 형태, 구조, 범위가 매우 다양하다. 저울의 형태학은 그것이 유래한 물고기의 종을 식별하는 데 사용될 수 있다.
대부분의 뼈 있는 물고기는 연어와 잉어의 사이클로이드 비늘이나 횃불의 씨노이드 비늘, 철갑상어와 가어의 가노이드 비늘로 덮여 있다. 수족구(상어와 광선)는 플래카로이드 비늘로 덮여 있다. 어떤 종은 대신 악어로 덮여 있고, 어떤 종은 피부 일부나 전체에 외부 덮개가 없다.
물고기 비늘은 물고기의 신경계의 일부로서 진피의 중간층으로부터 생산되는데, 이것은 파충류 비늘과 구별된다.[2][3] 포유류에서 치아와 모발 발달에 관여하는 동일한 유전자가 규모 발달에도 관여하고 있다. 수족류의 플라코이드 비늘은 피부 의치라고도 하며 구조적으로 척추동물 치아와 동질적이다. 대부분의 물고기들은 또한 박테리아, 곰팡이, 바이러스 같은 병원균으로부터 보호할 수 있고 물고기가 헤엄칠 때 표면 저항을 줄일 수 있는 점액이나 점액층으로 덮여 있다.
테로돈트 저울

화석 물고기의 가장 풍부한 형태인 로돈트의 뼈 있는 비늘은 잘 이해된다. 이 비늘은 유기체의 수명 내내 형성되어 벗겨졌고, 죽은 후에 빠르게 분리되었다.[4]
기계적 손상에 내성이 있고 비교적 화석화되기 쉬운 조직인 Bone은 내부 디테일을 보존하는 경우가 많아 비늘의 조직과 성장을 자세히 연구할 수 있다. 비늘은 덴틴으로 구성된 성장하지 않는 "크라운"으로 구성되며, 때로는 이름이 오르게 된 에나멜로이드 상부 표면과 아스피딘 베이스가 있다.[5] 그것의 성장 기반은 세포가 없는 뼈로 만들어지는데, 이것은 때때로 물고기의 측면에 그것을 고정시키기 위한 고정장치 구조를 개발했다.[6] 그 외에도, 뼈 성장에는 다섯 가지 유형이 있는데, 이는 테로돈트 내 5가지 자연 그룹 또는 최종 멤버 메타(또는 정형) 덴틴과 메소덴틴 조직 사이의 스펙트럼을 나타낼 수 있다.[7] 각각의 5개의 척도 형태는 더 파생된 물고기 군집들의 척도를 닮은 것으로 보이며, 이는 로돈트 군집들이 물고기 군집들의 뒤를 잇는 줄기 군집들일 수 있음을 암시한다.[6]
그러나, 종을 구별하기 위해 척도 형태학만을 사용하는 것은 몇 가지 함정이 있다. 각 유기체 내에서, 척도 모양은 신체 부위에 따라 크게 달라지는데,[8] 중간 형태는 서로 다른 영역 사이에 나타나며, 설상가상으로 척도 형태학은 한 영역 내에서 일정하지 않을 수도 있다. 사물을 더 혼란스럽게 하기 위해, 척도 형태학은 taxa에만 고유한 것이 아니며, 서로 다른 두 종의 동일한 영역에서 구별할 수 없을 수도 있다.[9]
비록 궁극적으로 그러한 수렴 형질을 사용하는 것은 오류가 발생하기 쉽지만, 로돈트의 형태학과 역사학은 그들의 다양성을 계량화하고 종을 구별하는 주요한 도구를 제공한다. 그럼에도 불구하고, 세 그룹으로 구성된 프레임워크는 규모 형태학 및 이력학에 기초하여 제안되었다.[7] 현대의 상어 종과의 비교를 통해 로돈트 비늘이 기능적으로 현대의 장상어의 비늘과 유사하다는 것을 보여주었고, 마찬가지로 생태학적 틈새의 광범위한 비교를 가능하게 했다.[10]
코스모이드 저울

코스모이드 비늘은 초기 폐어(딥노이 하위급) 중 일부를 포함한 고대 로브핀 어류와 변형된 형태의 살아있는 실러캔스를 포함한 크로소프테리지기(Crossoptygiii)에서만 발견된다(아래 엘라스모이드 비늘 참조). 그것들은 아마 플라코이드 비늘의 융합에서 유래되었을 것이다. 비늘의 안쪽은 이소포핀이라고 불리는 촘촘한 항성뼈로 이루어져 있다. 그 위에 혈관과 함께 공급되는 스펀지나 혈관뼈 층이 있고, 이어 바이트로덴틴의 피상적인 외부 코팅이 있는 코스미네라는 복잡한 틀니 모양의 층이 있다. 윗면은 케라틴이다. 우주비늘은 성층 골층의 성장을 통해 크기가 증가한다.[11]
엘라스모이드 비늘

엘라스모이드 비늘은 유스테놉테론에서처럼 주로 뼈로 구성된 결핵성 층인 아소페닌이라 불리는 촘촘한 성층 콜라겐 뼈로 이루어진 얇고 임브릭화된 비늘이다. 첫 번째 로브핀 생선에 있던 틀니 층은 대개 현존하는 쾰라칸스에서처럼 감소하거나, 아니면 완전히 존재하지 않는 폐어, 데보니아 유스테놉테론에서 감소한다.[12] 엘라스모이드 비늘은 물고기 진화의 과정에서 여러 번 나타났다. 그것들은 모든 현존하는 물고기들과 멸종된 폐피쉬들과 같은 일부 로브핀 어류들뿐만 아니라, 코즈미네이트가 부족하고 진짜 우주비늘보다 더 얇은 우주비늘을 변형한 실라칸스 어류에도 존재한다. 그것들은 또한 유스테놉테론, 아미이드, 텔레osts와 같은 일부 테트라포도모르프에도 존재하는데, 사이클로이드 비늘과 씨테노이드 비늘은 가장 미네랄이 적은 엘라스모이드 비늘을 나타낸다.
제브라피쉬 엘라스모이드 비늘은 뼈의 광물화 과정을 연구하기 위해 연구실에서 사용되며, 유기체 밖에서 배양(케프트)할 수 있다.[13][14]
가노이드 비늘
_(3149758934).jpg)

가노이드 비늘은 철갑상어, 노젓기, 가르, 보우핀, 비취르에서 발견된다. 그것들은 코스모이드 비늘에서 유래되었고 톱니 모양의 가장자리가 종종 있다. 코스메인 대신 딱딱한 에나멜 같은 덴틴 층을, 비트로덴틴 대신 가노인이라고 하는 무기 뼈 염분을 층층이 덮여 있다.
가노인은 가노이드 비늘의 특징적인 성분이다. 그것은 가르, 비취르, 퀼라칸스와 [15]같은 일부 비텔레그라운드의 두개골과 지느러미뿐만 아니라 비늘을 덮는 유리처럼 종종 다층 광물화 조직이다.[16][17] 그것은 막대 모양의 아파타이트 결정체로 구성되어 있다.[18] Ganoine은 예를 들어 줄기 그룹인 Actinoptrigian Cheirolepis의 저울에서 발견되는 광학고기의 고대 특징이다.[17] 흔히 광선 핀 어류의 시너포모픽 성격으로 여겨지지만, 멸종된 아칸토디에서도 가노인이나 가노인 같은 조직이 발견된다.[17] 그것은 가노인이 척추동물의[15] 치아 에나멜에 균질하거나 심지어 에나멜의 한 종류로 간주되는 것으로 제안되었다.[18]
 암블리프테루스 스트리투스 | 멸종된 카본리퍼스 어류의 가노이드 비늘, 암블리프테루스 선조체. (a) 비늘 4개의 외면을 나타내고, (b) 비늘 2개의 내면을 나타낸다. 암블리프테루스(Amblypterus)의 각 암블리프테루스(Rhomboidal) 모양의 가노이드 비늘은 내부 표면에 능선이 있으며, 이는 다음 비늘의 한 단에 맞는 돌출된 펙으로 만들어지는데, 이는 집의 지붕에 기와를 붙이는 방식과 유사하다. |  |
대부분의 가노이드 비늘은 회전목마(다이아몬드 모양)이며, 페그앤소켓 관절에 의해 연결된다. 그것들은 보통 두께가 두껍고 다른 비늘처럼 겹치기 보다는 조각톱처럼 서로 잘 맞는다.[19] 이와 같이 가노이드 비늘은 거의 관통할 수 없고 포식으로부터 뛰어난 보호가 된다.

.png)


철갑상어에서는 비늘이 옆면과 뒷면을 따라 무기고판으로 크게 확대되는 반면, 보우핀에서는 비늘의 두께가 크게 줄어 사이클로이드 비늘과 닮았다.

악어 가의 가노이드 비늘로 만든 귀걸이

가노이드 비늘을 가진 원시 레이핀의 화석


.jpg)
미국 원주민들과 카리브해 사람들은 악어 가의 단단한 가노이드 비늘을 화살촉, 가슴받이, 쟁기를 덮는 차폐물로 사용했다. 요즘은 이런 비늘로 장신구가 만들어진다.[20]
렙토이드 비늘
렙토이드(본리지) 비늘은 고차량의 뼈대 어류인 텔레osts(더 많이 파생된 가오리 껍질)에서 발견된다. 이 비늘의 바깥쪽은 뼈의 능선으로 부채꼴 모양으로 되어 있고 안쪽은 섬유질의 결합조직으로 교차되어 있다. 렙토이드 비늘은 다른 종류의 비늘에 비해 얇고 반투명하며, 에나멜처럼 굳어진 층이나 덴틴 층이 부족하다. 가노이드 비늘과 달리 물고기가 자랄수록 동심층에는 비늘이 더해진다.[21]
렙토이드 비늘은 기와 같이 머리부터 꼬리까지의 구성이 겹쳐져 있어 우주비늘이나 가노이드 비늘보다 유연성이 뛰어나다. 이렇게 배열하면 몸 위로 물이 더 부드럽게 흐를 수 있고, 드래그를 줄일 수 있다.[22] 어떤 종의 비늘은 에비뉴라고 불리는 불균일한 계절성장의 띠를 보여준다. 이 띠들은 물고기의 노화를 위해 사용될 수 있다.
렙토이드 비늘은 사이클로이드와 씨테노이드의 두 가지 형태로 나타난다.
사이클로이드 저울
사이클로이드(원형) 비늘은 질감이 매끄럽고 균일하며, 바깥쪽 가장자리나 여백이 매끈하다. 연어, 잉어 등 지느러미가 부드러운 물고기에게 가장 흔하다.

.jpg)









체노이드 비늘
Ctenoid (toothed) 비늘은 외측 또는 후측 가장자리를 따라 ctenii라고 불리는 작은 이빨이나 스핀들을 가지고 있다는 점을 제외하면 사이클로이드 비늘과 같다. 이러한 치아 때문에 비늘은 질감이 거칠다. 그것들은 보통 횃대 같은 지느러미가 있는 물고기에서 발견된다. 이 비늘은 거의 뼈가 없으며, 수산화칼슘과 탄산칼슘을 함유한 표면층과 콜라겐으로 구성된 더 깊은 층으로 구성되어 있다. 다른 척도 유형의 에나멜은 표면적인 능선과 cteni로 감소한다.


 |  |
즐린류의 물고기 비늘에 이빨의 사이즈 위치와, 금줄 Cetonurus crassiceps 쇼에서 이 비늘로 변경될 수 있다. | |

_by_M._L._Nievera_(colored).png)





Ctenoid 비늘, 다른 표피 구조에 비슷한, 원판에서 독특한 세포 분화들은 포피에서 비롯된다 다른 구조물에 독점을 만든다 유래되었다.[26] 개발 caudal 지느러미 근처에 물고기의 측면 라인을 따라 시작한다.[27] 그 개발 과정 섬유 아세포의 표피와 진피 사이에 축적과 함께 시작한다.[26] 콜라겐 원섬유는 광화 작용의 개시로 이어지는 진피 층에 스스로를 조직하기 시작한다.[26] 그 규모의 원주는 처음, 중복되는 사와 함께 mineralize 두께에 이어 자란다.[26]
Ctenoid 규모 추가적인 세가지 유형으로: 나눌 수 있다.
- 는 규모의 마진이 오목한 자국 및 돌출부 곰 Crenate 비늘,.
- 그 규모는 규모 자체에 지속적이면 가시 곰Spinoid 비늘,.
- 는 크기에 가시가 뚜렷한 구조물 진정한 빗 모양의 비늘,.
대부분의ray-finned 물고기들 빗 모양의 비늘이 있습니다. 반면에 다른 종으로 여성에서 남성과 원형의 비늘로 빗 모양의 비늘이 가자 미목 어류의 일부 종들 약점에eyed 측과 원형의 비늘에, 빗 모양의 비늘이 있습니다.
반사



많은 teleost 생선에는 거울로 기능과silvered 유리의 외형을 극히 반사율이 높은 비늘로 덮여 있다. silvering을 통해 반사 또는 지배적인 사람이 펼쳐진 바다 생선, 특히 맨 위 100미터에 살고 있는 널리 퍼져 있다. A투명성 효과 동물의 몸은 극히 반사율이 높은 만들기 위해 질산은으로 감광시킴으로써 이루어질 수 있다. 그래서 거울 수직 방향 측면에서 물고기와 같은 동물들 안 보이게 만든다 바다에서 중간 깊이에서, 빛 위로부터 나온다.[28]
바다도끼는 측면(옆으로) 극도로 납작하게 만들어 몸통 두께가 밀리미터에 불과하고, 몸통도 알루미늄 호일을 닮을 정도로 은빛을 띤다. 거울은 구조적인 색상을 제공하는 데 사용되는 것과 유사한 미세한 구조로 구성된다. 구아닌 결정의 5-10개의 스택은 파장 사이의 간격을 두어 구조적으로 간섭하고 거의 100% 반사를 달성한다. 도끼가 서식하는 깊은 물속에서는 파장 500나노미터의 푸른 빛만이 아래쪽으로 스며들어 반사하면 되기 때문에 125나노미터 떨어져 있는 거울이 좋은 위장 기능을 한다.[28]
상해의 대부분의 물고기는 은빛으로 위장된다. 얕은 물 속에 사는 청어와 같은 물고기에서는 거울이 파장의 혼합물을 반사해야 하며, 따라서 물고기는 다양한 스페이스의 결정 스택을 가지고 있다. 단면으로 둥근 몸을 가진 물고기의 또 다른 문제는 거울이 수평으로 반사되지 않기 때문에 피부에 평평하게 놓이면 효과가 없다는 것이다. 전체 미러 효과는 수직 방향의 많은 작은 반사경을 통해 달성된다.[28]
이런 성질을 가진 생선 비늘은 화장이나 립스틱에 반짝거리는 효과를 줄 수 있기 때문에 일부 화장품에 사용된다.[29]
플라코이드 비늘

플래코이드 비늘은 상어, 광선 등 수족관에서 발견된다. 그것들은 또한 피부 틀니클이라고도 불린다. 플라코이드 비늘은 구조적으로 척추동물 치아와 동일하며("덴티클"은 "작은 치아"를 의미하며, 혈관과 함께 공급되는 중심펄프강을 가지고 있고, 틀니 원뿔층으로 둘러싸여 있으며, 모두 진피 위에 놓여 있는 직사각형의 기저판 위에 놓여 있다. 가장 바깥쪽 층은 주로 무기질의 에나멜과 같은 물질인 비트로덴틴으로 구성되어 있다. 플라코이드 비늘은 크기가 커질 수 없지만, 오히려 물고기의 크기가 커질수록 비늘이 더 많이 추가된다.
의치 청어의 머리 밑에서도 비슷한 비늘을 발견할 수 있다. 광선에서는 체중계 커버리지의 양이 훨씬 적다.
상어피부

상어의 피부는 거의 전체적으로 작은 플래카이드 비늘로 덮여 있다. 비늘은 가시가 받쳐주는데, 가시가 역방향으로 쓰다듬으면 거친 느낌이 들지만, 물의 전방 이동에 의해 평평해지면 유체역학적으로 끌리는 것을 줄이고 난류를 줄이는 아주 작은 배틀을 만들어, 뼈대 물고기에 비해 수영이 효율적이고 조용해진다.[30] 연꽃 효과를 발휘하여 항풀링의 역할도 한다.[31]
모든 틀니클은 진피에 뿌리내린 신경과 동맥 공급이 있는 내부 펄프 캐비티로 구성되어 있어, 틀니클에 점액을 공급한다.[32] 틀니클은 눈금의 표면에서 튀어나온 리블트 구조를 가지고 있다. 현미경으로는 이 리블트는 눈금에서 나오는 갈고리나 굴곡처럼 보일 수 있다. 틀니에서 튀어나온 전체적인 모양은 상어의 종류에 따라 달라지며 일반적으로 두 가지 외관으로 설명할 수 있다.[33] 첫째는 능선이 상어 아래로 횡방향으로 내려와 물의 흐름과 평행하게 놓여 있는 눈금이다. 두 번째 형태는 상어의 후면을 겨냥하여 표면에서 갈고리 모양의 리블렛처럼 생긴 것이 매끄러운 눈금이다.[33] 두 개의 라블트 모양은 상어의 피부에서 더 멀리 떨어진 층을 흐르게 하는 난류 경계층을 만드는 데 도움이 된다.[34]
상어는 뼈가 있는 물고기와 달리 유연한 콜라겐 섬유로 만들어진 복잡한 피부 코르셋을 몸 주위를 둘러싼 나선망으로 배열하고 있다. 코르셋은 바깥쪽 골격으로 작용하여 수영 근육에 애착을 주어 에너지를 절약한다.[35] 이 플래코이드 비늘이 몸체에 있는 위치에 따라 유연하고 수동적으로 세울 수 있어 공격 각도를 바꿀 수 있다. 이 비늘은 또한 흐름의 방향으로 정렬된 라블렛을 가지고 있는데, 이 라블렛은 상어의 피부에 작용하는 드래그력을 감소시켜 높은 속도의 크로스 스트림 흐름을 억제한다.[36]
척도 형태학
비늘의 일반적인 해부학적 구조는 다르지만, 모두 세 부분으로 나눌 수 있다: 왕관, 목, 밑단. 척도 플라이어티는 척도 베이스의 크기와 관련이 있다. 유연성이 높은 저울은 기초가 작아 층하방체에 덜 단단하게 부착된다. 빠르게 헤엄치는 상어의 왕관에는 앞쪽에서 뒤쪽으로 이어지는 평행 리블렛이나 능선이 잇다.[37]
눈금의 세 가지 성분을 분석하면, 틀니클의 밑부분이 유체 흐름의 어떤 부분과도 접촉하지 않는다는 결론을 내릴 수 있다.[38] 하지만 의치의 왕관과 목은 중요한 역할을 하며 피부 표면 근처에서 발견되는 격동의 소용돌이와 에디를 만들어 내는 역할을 한다.[38] 틀니클은 매우 다양한 모양과 크기를 가지고 있기 때문에, 모든 형태가 같은 종류의 난류를 생성하지는 않을 것이라고 예상할 수 있다. 최근 한 연구 실험에서 초승달 모양의 초승달 구조를 가진 상어 틀티클의 생체모방 샘플이 트랙션 테이블을 미끄럼틀로 사용하여 물 탱크에서 테스트되었다. 실험 결과, 덴티클이 있는 표면은 부드러운 샘플 대비 전체적으로 10% 드래그 감소를 경험했다. 이러한 드래그 감소의 이유는 난류하는 항아리들이 틀니 사이에 끼여 층류 흐름에 대한 '쿠션 like' 장벽을 만들었기 때문이다.[39] 이와 같은 유형의 실험은 생체모방 샘플에 더 많은 변화를 구현한 다른 연구 그룹에 의해 수행되었다. 그러나 두 번째 그룹은 첫 번째 그룹과 같은 결론에 도달했는데, 그들의 실험에는 높은 수준의 실험 정확도를 달성할 수 있는 표본 내의 변동이 더 많이 포함되었기 때문이다. 결론적으로 그들은 더 실용적인 모양들이 복잡한 능선을 가진 모양들보다 더 오래 지속된다고 말했다. 실제 형태는 낮은 프로필이었고 사다리꼴 또는 반원형 수조 모양의 단면을 포함하고 있었으며, 효과는 낮았지만 항력을 6~7%[40] 줄였다.
드래그 감소

상어는 여러 가지 다른 방법을 통해 드래그와 전체 운송 비용을 줄인다. 압박 항력은 상어를 지나쳐 앞으로 나아가기 위해 밀어내는 부피의 양 때문에 상어의 앞면과 뒷면의 압력차에서 생성된다.[41] 이런 종류의 끌림도 층류 흐름에 정비례한다. 물고기 주위에 층류 흐름이 증가하면 압력 끌림도 증가한다.[42] 마찰 항력은 상어의 피부에 대한 액체의 상호작용의 결과로, 물고기의 표면에 대해 경계층이 어떻게 변하느냐에 따라 달라질 수 있다.[41]
라블렛은 점성 하위 계층에서 스트림과 현상의 교차 스트림 변환을 방해한다. 그 메커니즘은 복잡하고 아직 완전히 이해되지 않았다. 기본적으로 라블렛은 라블렛에 의해 형성된 계곡에 볼텍스가 들어맞지 못하기 때문에 표면 근처의 볼텍스 형성을 억제한다. 이것은 소용돌이를 표면에서 더 위로 밀어 올려, 오직 라블렛 끝과 상호작용을 하면서, 계곡에 어떠한 높은 속도 흐름도 일으키지 않는다. 이 고속 흐름은 이제 표면적이 매우 작은 라블트팁과만 상호작용하기 때문에, 드래그를 일으키는 운동량 전달은 이전보다 훨씬 낮아져 드래그를 효과적으로 감소시킨다. 또한, 이것은 교차 스트림 속도 변동을 감소시켜 모멘텀 전달에도 도움이 된다.[37]
최근 연구에 따르면, 붕괴 전/후 시스템이 근방 경계층에 있다는 것이 밝혀졌는데, 이 경계 층은 감소하는 속도로 두꺼워졌다가 마침내 붕괴되기 전에 갑자기 격동의 격변을 겪게 된다. 이 시스템은 완전히 자율적으로 조절하고 성장과 부패 주기를 매개한다; 균류는 성장 기간 동안 축적되어 갑자기 벽에서 들어올리는 머리핀 통풍구들의 스트라우할 배열로 정화된다. 리프팅 포티스는 상어의 표면으로부터 경계층을 밀어내고 멀리 떨어져서 물고기가 경험하는 전체적인 드래그를 감소시키는 것이다.[43]
거칠고 사포처럼 생긴 상어와 광택피부의 질감이 그 질감과 결합되어 샤그린이라 불리는 라위드 가죽의 원천으로 평가받게 되었다. 상어 샤그린이 역사적으로 많이 사용된 것 중 하나는 칼을 위한 손잡이를 만드는 것이었다. 나무판에 상어 껍질 조각을 붙여 오로시키라는 그라테를 만드는 일본 요리에도 거친 피부결이 사용된다. 비늘이 작아서 음식을 아주 곱게 간다.
기술 적용
.jpg)
해양 산업에는 시장이 매우 크고, 주름 방지 표면이 필요하다. 비전문가로 말하면, 반칙은 바나클, 조류, 녹색 슬러지 같은 주변 환경의 물질로 무언가가 감싸지는 과정으로 알려져 있다. 상어가 저울에 쌓이거나 자라지 않는 유일한 물고기라는 사실 때문에, 피부 틀티클은 이런 종류의 적용에 있어 매우 유망한 연구 분야다. 미 해군의 연구에 따르면 생체모방 물질을 설계할 수 있다면 최대 [44]45%의 군함 연료 절감 효과를 잠재적으로 유발할 수 있다고 한다.
상어를 포함한 수생 생물체의 구조를 바탕으로 한 생체모방 물질과 표면의 예가 많다. 그러한 애플리케이션은 공기, 물, 기름과 같은 유동 매체를 통해 보다 효율적인 이동을 가능하게 한다.
상어의 피부를 모방한 표면도 미생물과 해조류가 잠수함이나 선박의 선체에 코팅되지 않도록 하기 위해 사용되었다. 한 품종은 "상어"로 거래된다.[45][46]
상어 피부를 복제하는 많은 새로운 방법에는 몰드를 만들기 위해 폴리디메틸실록산(PDMS)을 사용하는 것이 포함된다. 보통 그 과정은 상어 가죽의 평평한 조각을 가져다가 PDMS로 덮어서 틀을 만들고 상어 피부 복제본을 얻기 위해 PDMS를 다시 그 틀에 붓는 것을 포함한다. 이 방법은 초저공포성 성질을 가진 생체모방 표면을 만들어 연꽃 효과를 나타내는 데 사용되어 왔다.[45] 한 연구에서는 이러한 생체모방 표면이 최대 9%[36]까지 항력을 감소시키는 반면 펄럭이는 움직임 항력 감소는 12.3%[47]에 달한다는 것을 발견했다.
해양 응용과 함께 항공우주 산업도 이러한 생체모방 설계의 혜택을 누릴 수 있다. 파라메트릭 모델링은 로우 및 하이프로필 보텍스 생성기와 같은 광범위한 설계 변형을 가진 상어 틀니클에서 수행되었다.[48] 이러한 생체모형은 다양한 비행기의 날개에 의치처럼 생긴 구조물을 응용하는 효과를 보기 위해 설계되고 분석되었다. 시뮬레이션 도중 샘플이 낮은 각도와 높은 각도의 공격 반응 방식을 변화시켰다는 점에 주목했다. 테스트한 낮은 샘플과 높은 프로필의 샘플 중 낮은 프로필의 소용돌이 발생기는 현재의 부드러운 날개 구조를 323% 능가했다. 이 같은 성능 증가는 피부 마찰로 경계층에서 손실된 운동량을 보충하는 틀티클 웨이크 및 스트림 현상의 분리 거품 때문이다.[48]
스쿠테스
_Waka,_Japan.jpeg)
scute는 저울과 비슷하고 같은 기능을 한다. 표피에서 형성되는 물고기의 비늘과 달리 피부의 아래쪽 혈관층에 소액이 형성되고 표피 원소는 윗면만 된다. 살아있는 진피에서 형성되는 이 악취는 표면적으로는 비늘과 비슷한 뿔 같은 외층을 생성한다.
Scute는 방패를 위해 라틴어에서 왔으며, 다음과 같은 형태를 취할 수 있다.
- 외부 방패 모양의 뼈판, 또는
- 변형되고 두꺼워진 저울은 종종 구부러지거나 가시가 돋거나 또는
- 일반적으로 측면 선과 관련된 돌출된 (경직되고 강한) 척도 또는 카우달 켈을 형성하는 카우달 페달 또는 복측 종단을 따라 돌출된 (경직되고 강한) 척도.
파인코네피쉬와 같은 어떤 물고기는 완전히 또는 부분적으로 소변으로 덮여 있다. 강 청어와 실핀은 보호에 사용되는 높은 비늘과 뾰족한 뾰족한 비늘로 이루어진 복줄의 소쿠르를 가지고 있다. 어떤 잭들은 양쪽 측면의 가로선을 따라 한 줄로 늘어선 악어들을 가지고 있다.
규모 개발
비늘은 일반적으로 물고기의 발달에 늦게 나타난다. 제브라피쉬의 경우 수정 후 30일이면 비늘을 형성하기 시작하는 데 필요한 층이 달라져 정리된다. 이를 위해서는 중생방 통합이 일어나고, 그 후 형태생성이 유도되며, 마지막으로 분화 또는 후기 변성의 과정이 발생한다.[49][50]
- 중간합성: 중핵의 결합이나 구조는 진피의 발달 과정에서 비롯된다. 이 과정은 그 물고기가 수레인지 뼈인지에 달려있다. 갑상선 어류의 경우 구조는 두 개의 층의 형성을 통해 발생한다. 첫째는 피상적이고 넓으며 둘째는 가늘고 콤팩트하다. 이 두 층은 중피세포에 의해 분리된다. 뼈 있는 물고기는 콜라겐 섬유에 의해 수직으로 조직된 무세포 기질을 생성한다. 그 후, 두 물고기의 경우 섬유블라스트가 길어진다. 이들은 눈금이 형성되기 전에 통합되어 진피판을 시작하기 위해 중간막의 콤팩트한 층을 관통한다.[49][50][51]
- 형태생성 유도: 형태생식은 표피와 진피와 결합하여 생기는 표피의 형성에 기인한다. 형태생식은 섬유화합물이 콤팩트 메센치메스의 상부로 옮겨지는 시점에 시작된다. 이 과정을 거치면서 상피의 기저세포가 구분을 이루는 층을 형성하는데, 이 층은 중핵의 상부에 위치한다. 이후 이 세포들은 스케일 프리모듐이 발생할 영역에서 분화한다.[49][50][51]
- 분화 또는 후기 변성: 이러한 분화는 형성되는 규모의 유형에 따라 두 가지 다른 형태에 의해 생성된다. 엘라스모이드 비늘(사이클로이드, ctenoids)의 형성은 표피질의 매트릭스 사이의 공간 형성을 통해 발생한다. 이 공간에는 콜라겐 섬유가 들어 있다. 이 공간 주위에서 탄소성체는 분화하며, 스케일 형성에 필요한 물질을 생성하는데 책임이 있다. 이후 매트릭스 광물화가 일어나 이를 식별하는 강체 특성을 척도로 획득할 수 있다.[49][50][51]
가노이드 비늘은 엘라스모이드 비늘과 달리 광물화 콜라겐과 비미네랄화 콜라겐으로 부위별로 구성돼 있다. 이러한 형성은 메센키메스의 표면 세포가 매트릭스에 들어가면서 발생하며, 후자는 콜라겐 섬유로 구성되어 있으며 혈관 모세혈관 주위에 위치하여 혈관 공동이 생기게 된다. 이때 엘라스모브레스는 골육상세포로 대체되어 뼈를 형성한다. 오순화되지 않은 눈금 행렬의 패치는 메센치메와 결합을 유지할 수 있도록 압축된 콜라겐으로 구성되어 있다. 이것은 샤르피 섬유로 알려져 있다.[49][50][51]
물고기의 체중계 형성의 발달을 조절하는 유전자 중 하나가 소닉 고슴도치(shh) 유전자로, 유기생식에 관여하고 세포 통신 과정에서 (shh) 단백질을 통해 체중계 형성을 가능하게 한다.[52][53] 트리글리세리드와 콜레스테롤의 운반과 신진대사가 가능한 아폴리포프로테인 E(ApoE)는 쉿과 상호작용을 하는데, 이는 아포E가 쉿 신호 경로에 콜레스테롤을 제공하기 때문이다. 세포 분화와 상호작용 과정에서 아포E 전사 수준이 높아 이 단백질이 저울의 늦은 발달을 위해 중요하다는 결론을 내린 것으로 나타났다.[52][53]
수정척도




다른 그룹의 물고기들은 다양한 기능을 제공하기 위해 많은 변형 비늘을 진화시켰다.
- 거의 모든 물고기는 물 움직임을 감지하는 기계수용체 시스템인 횡선을 가지고 있다. 뼈가 있는 물고기에서는 가로선을 따라 있는 비늘이 중앙의 모공을 가지고 있어 물이 감각 세포와 접촉할 수 있다.
- 도그피쉬 상어와 치마라의 등지느러미 가시, 가오리의 따끔따끔한 꼬리 가시, 톱니나 톱니 모양의 톱니 등이 융합되어 변형된 플라코이드 비늘이다.
- 외과 어류는 외과적 발톱의 양쪽에 변형된 비늘인 메스 모양의 날을 가지고 있다.[54]
- 어떤 청어, 멸치, 반달가슴살은 낙엽성 비늘을 가지고 있는데, 이것은 쉽게 벗겨지고 포식자 탈출에 도움이 된다.
- 남성 퍼시나 대럴터스는 골반 지느러미와 항문 사이에 확대된 지적 비늘이 줄지어 있다.
- 고슴도치는 큰 외부 가시로 변형된 비늘을 가지고 있다.
- 반면 복어는 고슴도치보다 가시가 얇고 숨은 가시가 많아 물고기가 부풀어야 눈에 띈다. 고슴도치 물고기와 달리 이 가시들은 변형된 비늘이 아니라 다른 척추동물에서 깃털과 털을 생산하는 유전자의 같은 네트워크의 지배하에 발달한다.[55][56]
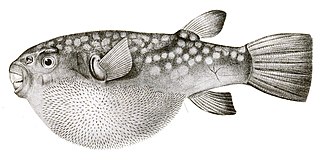
복어 가시는 변형된 눈금이 아니라 독립적인 유전자 네트워크에 의해 개발된다.


비늘 없는 물고기

만다린피쉬는 비늘이 부족하고 냄새나고 쓴 슬라임이 겹겹이 쌓여 스스로를 보호한다.

비늘이 없는 물고기는 단단한 가죽 껍질이나 뼈 판과 같이 비늘이 제공할 수 있는 보호 비늘의 대안으로 진화한다.
- 턱이 없는 물고기(등고기와 해파지)는 비늘이 없고 피부뼈가 없는 매끄러운 피부를 가지고 있다.[57] 램프리스는 가죽이 질긴 피부로부터 약간의 보호를 받는다. 개고기는 위협을 받으면 많은 양의 점액이나 점액을 배출한다.[58] 그들은 스스로를 손으로 묶을 수 있고, 가면서 슬라임을 긁어내고 포식자로부터 해방시킬 수 있다.[59]
- 비록 몇몇 종들은 아주 부드러운 사이클로이드 비늘로 덮여있지만 대부분의 뱀장어는 암이 없다.
- 대부분의 메기는 비늘이 부족하지만, 몇몇 집단은 피부판이나 어떤 종류의 악어의 형태로 신체용 갑옷을 가지고 있다.[60]
- 만다린피쉬는 비늘이 부족하고 냄새나고 쓴 슬라임 층이 있어 질병을 차단하고 포식자를 억제할 수 있어 그들의 밝은 색채가 무감각하다는 것을 암시한다.[61]
- 앵글러피쉬는 살갗이 헐렁하고 가는 경우가 많으며 갈퀴가 있는 피부 따갑기나 덩굴로 덮여있지만 일정한 비늘은 가지고 있지 않다. 그들은 포식자의 주의를 피하기 위해 위장술에 의존하는 반면, 그들의 느슨한 피부는 포식자들이 포식자를 잡기 어렵게 만든다.
관상어, 해어, 복어, 밀렵꾼, 그리고 스틱백어 가족을 포함한 많은 뼈다귀 집단은 구조적으로 플래코이드 비늘과 비슷한 외부 뼈 접시를 포식자에 대한 보호용 무기로 개발했다.
- 해마는 비늘이 부족하지만 몸길이에 고리로 배열된 뼈판 갑옷 위로 늘어진 얇은 피부를 가지고 있다.
- 복어에서는 판들이 서로 융합되어 전신을 감싸고 있는 단단한 껍질이나 외골격을 형성한다. 이 뼈판들은 변형된 비늘이 아니라 오식화된 피부다. 이 무거운 갑옷 박스피쉬 때문에 느린 동작으로 제한되지만, 다른 물고기들은 거의 성인을 먹을 수 없다.
 | |
뱀장어는 칼이 없는 것처럼 보이지만, 어떤 종은 아주 매끄러운 사이클로이드 비늘로 덮여 있다. | |


호키나 황어와 같은 어떤 물고기는 비늘을 가지고 태어나지만 자라면서 비늘을 벗긴다.
파일피쉬는 작은 뾰족탑과 함께 겹치지 않는 거친 비늘을 가지고 있어서 파일피쉬라고 불린다. 어떤 대파리는 비늘이 너무 작기 때문에 메스 없이 보인다.
두드러진 스케일링은 가로선과 코르슬렛을 따라 참치에만 나타나며, 어깨 부위에는 굵고 확대된 스케일 보호 띠가 있다. 그들의 몸통 참치는 대부분 비늘이 너무 작아서, 무심코 살펴보니 그들은 메스껍지 않은 것처럼 보인다.[62]


레비티쿠스
레비티쿠스의 한 유명한 구절은 "바다에 있는 모든 것 중... 바다와 강에서" 지느러미와 비늘이 모두 없는 것들은 "너에게 혐오스러운 것"이어야 하며, 먹히지 않을 수도 있다.[63] 이것은 모든 수생 무척추동물을 혐오와 부정으로 제거하고 비늘이 없는 물고기(지느러미가 없는 물고기는 없는 것 같다)도 제거한다.
토라와 탈무드의 초크나 신의 율례에 따르면, 고기가 코셔로 선언되려면 비늘과 지느러미가 있어야 한다.[64] "척도"의 정의는 코셔 물고기의 비늘이 눈에 보여야 하며, 손이나 스케일링 칼로 피부에서 쉽게 제거할 수 있다는 점에서 생물학에서 제시된 정의와 다르다.[64] 정교회 연합의 코셔 인증 기관에 따르면, 피부는 찢어지지 않고 비늘을 제거할 수 있다면 물고기는 코셔라고 한다.[65] 따라서 잉어와 연어는 코셔인 반면 비늘이 미세한 상어는 몸 밖으로 잘리지 않고는 쉽게 제거할 수 없는 철갑상어는 모두 코셔가 아니다. 다른 비코셔 어류로는 메기, 장어, 민물 대구, 뱀 고등어, 복어 등이 있다.[64]
레피도파기

레피도파기는 다른 물고기의 비늘을 먹는 것을 포함하는 물고기의 특별한 먹이 활동이다.[67] 레피도파기는 적어도 5개의 담수가족과 7개의 해양가족에서 독립적으로 진화해 왔다.[68]
생선 비늘은 케라틴과 에나멜 층과는 별개로 피부 부분과 단백질이 풍부한 점액층을 함유하고 있어 영양이 풍부할 수 있다. 그것들은 인산칼슘의 풍부한 공급원이다.[68] 그러나 스트라이크를 하기 위해 소비되는 에너지와 스트라이크 당 소비되는 비늘의 양은 미지근한 물고기의 크기에 제한을 두며, 그들은 보통 그들의 먹이보다 훨씬 작다.[68] 체중계 먹는 행동은 대개 음식의 부족과 극단적인 환경 조건 때문에 진화한다. 비늘을 먹는 것과 비늘을 둘러싼 피부는 틈새의 다른 곳에서는 구할 수 없는 단백질이 풍부한 영양분을 제공한다.[69]
물고기 턱은 보통 양쪽 대칭을 나타낸다. 체중계 섭취 시클리드 페리스소두스 마이크로레피스에서 예외가 발생한다. 이 물고기의 턱은 두 가지 뚜렷한 형태학적 형태로 나타난다. 하나의 형태는 턱이 왼쪽으로 비틀어져 있어 희생자의 오른쪽 옆구리에 있는 비늘을 더 쉽게 먹을 수 있다. 다른 형태는 턱이 오른쪽으로 꼬여 있어 희생자의 왼쪽 옆구리에 있는 비늘을 쉽게 먹을 수 있다. 모집단에서 두 형태에 대한 상대적 풍부함은 주파수 의존적 선택에 의해 조절된다.[66][70][71]
참고 항목
참조
- ^ 스케일 에티몬라인. 2019년 4월 28일 회수
- ^ Mongera, A.; Nüsslein-Volhard, C. (2013). "Scales of fish arise from mesoderm". Current Biology. 23 (9): R338–R339. doi:10.1016/j.cub.2013.02.056. PMID 23660349.
- ^ Sharpe, P. T. (2001). "Fish scale development: Hair today, teeth and scales yesterday?". Current Biology. 11 (18): R751–R752. doi:10.1016/S0960-9822(01)00438-9. PMID 11566120. S2CID 18868124.
- ^ Turner, S.; Tarling, D. H. (1982). "Thelodont and other agnathan distributions as tests of Lower Paleozoic continental reconstructions". Palaeogeography, Palaeoclimatology, Palaeoecology. 39 (3–4): 295–311. Bibcode:1982PPP....39..295T. doi:10.1016/0031-0182(82)90027-X.
- ^ Märss, T. (2006). "Exoskeletal ultrasculpture of early vertebrates". Journal of Vertebrate Paleontology. 26 (2): 235–252. doi:10.1671/0272-4634(2006)26[235:EUOEV]2.0.CO;2.
- ^ Jump up to: a b Janvier, Philippe (1998). "Early vertebrates and their extant relatives". Early Vertebrates. Oxford University Press. pp. 123–127. ISBN 978-0-19-854047-2.
- ^ Jump up to: a b Turner, S. (1991). "Monophyly and interrelationships of the Thelodonti". In M. M. Chang; Y. H. Liu; G. R. Zhang (eds.). Early Vertebrates and Related Problems of Evolutionary Biology. Science Press, Beijing. pp. 87–119.
- ^ Märss, T. (1986). "Squamation of the thelodont agnathan Phlebolepis". Journal of Vertebrate Paleontology. 6 (1): 1–11. doi:10.1080/02724634.1986.10011593.
- ^ Botella, H.; J. I. Valenzuela-Rios; P. Carls (2006). "A New Early Devonian thelodont from Celtiberia (Spain), with a revision of Spanish thelodonts". Palaeontology. 49 (1): 141–154. doi:10.1111/j.1475-4983.2005.00534.x.
- ^ Ferrón, Humberto G.; Botella, Héctor (2017). "Squamation and ecology of thelodonts". PLOS ONE. 12 (2): e0172781. Bibcode:2017PLoSO..1272781F. doi:10.1371/journal.pone.0172781. PMC 5328365. PMID 28241029.
- ^ Michael ALLABY "cosmoid scale." 동물학 사전. Encyclopedia.com. 2019년 10월 29일 <https://www.encyclopedia.com>.
- ^ 질버버그, L, 메이어, F.J., 로린, M. (2010) 악타 팔래ontologica Polonica 55: 459–470의 데보니아 사르콥터지안 유스테놉테론 포오르디에서 후두피 골격에 대한 미생물학적 및 역사학적 연구.
- ^ Bergen, Dylan J. M.; Kague, Erika; Hammond, Chrissy L. (2019). "Zebrafish as an Emerging Model for Osteoporosis: A Primary Testing Platform for Screening New Osteo-Active Compounds". Frontiers in Endocrinology. 10: 6. doi:10.3389/fendo.2019.00006. ISSN 1664-2392. PMC 6361756. PMID 30761080.
- ^ de Vrieze, E.; van Kessel, M. A. H. J.; Peters, H. M.; Spanings, F. A. T.; Flik, G.; Metz, J. R. (1 February 2014). "Prednisolone induces osteoporosis-like phenotype in regenerating zebrafish scales". Osteoporosis International. 25 (2): 567–578. doi:10.1007/s00198-013-2441-3. ISSN 1433-2965. PMID 23903952. S2CID 21829206.
- ^ Jump up to: a b Zylberberg, L.; Sire, J. -Y.; Nanci, A. (1997). "Immunodetection of amelogenin-like proteins in the ganoine of experimentally regenerating scales of Calamoichthys calabaricus, a primitive actinopterygian fish". The Anatomical Record. 249 (1): 86–95. doi:10.1002/(SICI)1097-0185(199709)249:1<86::AID-AR11>3.0.CO;2-X. PMID 9294653.
- ^ Sire, Jean-Yves; Donoghue, Philip C. J.; Vickaryous, Matthews K. (2009). "Origin and evolution of the integumentary skeleton in non-tetrapod vertebrates". Journal of Anatomy. 214 (4): 409–440. doi:10.1111/j.1469-7580.2009.01046.x. ISSN 0021-8782. PMC 2736117. PMID 19422423.
- ^ Jump up to: a b c Richter, M. (1995). "A microstructural study of the ganoine tissue of selected lower vertebrates". Zoological Journal of the Linnean Society. 114 (2): 173–212. doi:10.1006/zjls.1995.0023.
- ^ Jump up to: a b Bruet, B. J. F.; Song, J.; Boyce, M. C.; Ortiz, C. (2008). "Materials design principles of ancient fish armour". Nature Materials. 7 (9): 748–756. Bibcode:2008NatMa...7..748B. doi:10.1038/nmat2231. PMID 18660814.
- ^ Jump up to: a b Sherman, Vincent R.; Yaraghi, Nicholas A.; Kisailus, David; Meyers, Marc A. (1 December 2016). "Microstructural and geometric influences in the protective scales of Atractosteus spatula". Journal of the Royal Society Interface. 13 (125): 20160595. doi:10.1098/rsif.2016.0595. ISSN 1742-5689. PMC 5221522. PMID 27974575.
- ^ "Missouri Alligator Gar Management and Restoration Plan" (PDF). Missouri Department of Conservation Fisheries Division. January 22, 2013. Archived from the original (PDF) on May 6, 2016. Retrieved April 12, 2019.
- ^ 라글러, K. F., J. E. Bardach, R. R. Miller(1962) 이크시학. 뉴욕: John Wiley & Sons.
- ^ Ballard, Bonnie; Cheek, Ryan (2 July 2016). Exotic Animal Medicine for the Veterinary Technician. John Wiley & Sons. ISBN 978-1-118-92421-1.
- ^ Pouyaud, L.; Sudarto, Guy G. Teugels (2003). "The different colour varieties of the Asian arowana Scleropages formosus (Osteoglossidae) are distinct species: morphologic and genetic evidences". Cybium. 27 (4): 287–305.
- ^ Ismail, M. (1989). Systematics, Zoogeography, and Conservation of the Freshwater Fishes of Peninsular Malaysia (Doctoral Dissertation ed.). Colorado State University.
- ^ E.J. Brill (1953). The Fishes of the Indo-Australian Archipelago. E.J. Brill. pp. 306–307.
- ^ Jump up to: a b c d 카와사키, 켄타 C, 「시클리드 스케일 형태학의 유전자 분석」(2016). Masters Theses 2014년 5월 - 현재. 425. http://scholarworks.umass.edu/masters_theses_2/425
- ^ Helfman, Gene (2009). The Diversity of Fishes Biology, Evolution, and Ecology. Wiley-Blackwell.
- ^ Jump up to: a b c Herring, Peter (2002). The Biology of the Deep Ocean. Oxford: Oxford University Press. pp. 193–195. ISBN 9780198549567.
- ^ "There Are Probably Fish Scales In Your Lipstick". HuffPost India. 23 April 2015. Retrieved 6 May 2019.
- ^ Martin, R. Aidan. "Skin of the Teeth". Retrieved 28 August 2007.
- ^ Fürstner, Reiner; Barthlott, Wilhelm; Neinhuis, Christoph; Walzel, Peter (1 February 2005). "Wetting and Self-Cleaning Properties of Artificial Superhydrophobic Surfaces". Langmuir. 21 (3): 956–961. doi:10.1021/la0401011. ISSN 0743-7463. PMID 15667174.
- ^ Lauder, George V.; Wainwright, Dylan K.; Domel, August G.; Weaver, James C.; Wen, Li; Bertoldi, Katia (2016). "Structure, biomimetics, and fluid dynamics of fish skin surfaces". Physical Review Fluids. 1 (6): 060502. Bibcode:2016PhRvF...1f0502L. doi:10.1103/PhysRevFluids.1.060502. S2CID 18118663.
- ^ Jump up to: a b Feld, Katrine; Kolborg, Anne Noer; Nyborg, Camilla Marie; Salewski, Mirko; Steffensen, John Fleng; Berg-Sørensen, Kirstine (24 May 2019). "Dermal Denticles of Three Slowly Swimming Shark Species: Microscopy and Flow Visualization". Biomimetics. 4 (2): 38. doi:10.3390/biomimetics4020038. ISSN 2313-7673. PMC 6631580. PMID 31137624.
- ^ Fletcher, Thomas; Altringham, John; Peakall, Jeffrey; Wignall, Paul; Dorrell, Robert (7 August 2014). "Hydrodynamics of fossil fishes". Proceedings of the Royal Society B: Biological Sciences. 281 (1788): 20140703. doi:10.1098/rspb.2014.0703. ISSN 0962-8452. PMC 4083790. PMID 24943377.
- ^ Martin, R. Aidan. "The Importance of Being Cartilaginous". ReefQuest Centre for Shark Research. Retrieved 29 August 2009.
- ^ Jump up to: a b Hage, W.; Bruse, M.; Bechert, D. W. (1 May 2000). "Experiments with three-dimensional riblets as an idealized model of shark skin". Experiments in Fluids. 28 (5): 403–412. Bibcode:2000ExFl...28..403B. doi:10.1007/s003480050400. ISSN 1432-1114. S2CID 122574419.
- ^ Jump up to: a b Motta, Philip; Habegger, Maria Laura; Lang, Amy; Hueter, Robert; Davis, Jessica (1 October 2012). "Scale morphology and flexibility in the shortfin mako Isurus oxyrinchus and the blacktip shark Carcharhinus limbatus". Journal of Morphology. 273 (10): 1096–1110. doi:10.1002/jmor.20047. ISSN 1097-4687. PMID 22730019. S2CID 23881820.
- ^ Jump up to: a b Dou, Zhaoliang; Wang, Jiadao; Chen, Darong (1 December 2012). "Bionic Research on Fish Scales for Drag Reduction". Journal of Bionic Engineering. 9 (4): 457–464. doi:10.1016/S1672-6529(11)60140-6. ISSN 1672-6529. S2CID 137143652.
- ^ "Experimental investigations on drag-reduction characteristics of bionic surface with water-trapping microstructures of fish scales" (PDF).
- ^ Palmer, Colin; Young, Mark T. (14 January 2015). "Surface drag reduction and flow separation control in pelagic vertebrates, with implications for interpreting scale morphologies in fossil taxa". Royal Society Open Science. 2 (1): 140163. Bibcode:2015RSOS....240163P. doi:10.1098/rsos.140163. ISSN 2054-5703. PMC 4448786. PMID 26064576.
- ^ Jump up to: a b Lauder, George V.; Wainwright, Dylan K.; Domel, August G.; Weaver, James C.; Wen, Li; Bertoldi, Katia (18 October 2016). "Structure, biomimetics, and fluid dynamics of fish skin surfaces". Physical Review Fluids. 1 (6): 060502. Bibcode:2016PhRvF...1f0502L. doi:10.1103/PhysRevFluids.1.060502.
- ^ Muthuramalingam, Muthukumar; Villemin, Leo S.; Bruecker, Christoph (29 April 2019). "Streak formation in flow over Biomimetic Fish Scale Arrays". The Journal of Experimental Biology. 222 (Pt 16): jeb205963. arXiv:1904.12752. Bibcode:2019arXiv190412752M. doi:10.1242/jeb.205963. PMID 31375542. S2CID 139103148.
- ^ Bandyopadhyay, Promode R.; Hellum, Aren M. (23 October 2014). "Modeling how shark and dolphin skin patterns control transitional wall-turbulence vorticity patterns using spatiotemporal phase reset mechanisms". Scientific Reports. 4: 6650. Bibcode:2014NatSR...4E6650B. doi:10.1038/srep06650. ISSN 2045-2322. PMC 4206846. PMID 25338940.
- ^ Magin, Chelsea M.; Cooper, Scott P.; Brennan, Anthony B. (1 April 2010). "Non-toxic antifouling strategies". Materials Today. 13 (4): 36–44. doi:10.1016/S1369-7021(10)70058-4. ISSN 1369-7021.
- ^ Jump up to: a b Liu, Yunhong; Li, Guangji (15 December 2012). "A new method for producing "Lotus Effect" on a biomimetic shark skin". Journal of Colloid and Interface Science. 388 (1): 235–242. Bibcode:2012JCIS..388..235L. doi:10.1016/j.jcis.2012.08.033. ISSN 0021-9797. PMID 22995249.
- ^ "Sharklet Discovery Sharklet Technologies, Inc". www.sharklet.com. Retrieved 26 September 2018.
- ^ Lauder, George V.; Oeffner, Johannes (1 March 2012). "The hydrodynamic function of shark skin and two biomimetic applications". Journal of Experimental Biology. 215 (5): 785–795. doi:10.1242/jeb.063040. ISSN 1477-9145. PMID 22323201.
- ^ Jump up to: a b Domel, August G.; Saadat, Mehdi; Weaver, James C.; Haj-Hariri, Hossein; Bertoldi, Katia; Lauder, George V. (28 February 2018). "Shark skin-inspired designs that improve aerodynamic performance". Journal of the Royal Society Interface. 15 (139): 20170828. doi:10.1098/rsif.2017.0828. PMC 5832729. PMID 29436512.
- ^ Jump up to: a b c d e Sire, J.Y.; Huysseune, A.N.N. (2003). "Formation of dermal skeletal and dental tissues in fish: a comparative and evolutionary approach". Biological Reviews. 78 (2): 219–249. doi:10.1017/S1464793102006073. PMID 12803422. S2CID 19556201.
- ^ Jump up to: a b c d e Le Guellec, D.; Morvan-Dubois, G.; Sire, J.Y. (2004). "Skin development in bony fish with particular emphasis on collagen deposition in the dermis of the zebrafish (Danio rerio)". International Journal of Developmental Biology. 48 (2–3): 217–231. doi:10.1387/ijdb.15272388. PMID 15272388.
- ^ Jump up to: a b c d Sire, J.Y. (2001). "Teeth outside the mouth in teleost fishes: how to benefit from a developmental accident". Evolution & Development. 3 (2): 104–108. doi:10.1046/j.1525-142x.2001.003002104.x. PMID 11341672. S2CID 13353402.
- ^ Jump up to: a b Sire, J.Y.; Akimenko, M.A. (2003). "Scale development in fish: a review, with description of sonic hedgehog (shh) expression in the zebrafish (Danio rerio)". International Journal of Developmental Biology. 48 (2–3): 233–247. doi:10.1387/ijdb.15272389. PMID 15272389.
- ^ Jump up to: a b Monnot, M.J.; Babin, P.J.; Poleo, G.; Andre, M.; Laforest, L.; Ballagny, C.; Akimenko, M.A. (1999). "Epidermal expression of apolipoprotein E gene during fin and scale development and fin regeneration in zebrafish". Developmental Dynamics. 214 (3): 207–215. doi:10.1002/(SICI)1097-0177(199903)214:3<207::AID-AJA4>3.0.CO;2-5. PMID 10090147.
- ^ Sorenson, L.; Santini, F.; Carnevale, G.; Alfaro, M.E. (2013). "A multi-locus timetree of surgeonfishes (Acanthuridae, Percomorpha), with revised family taxonomy". Molecular Phylogenetics & Evolution. 68 (1): 150–160. doi:10.1016/j.ympev.2013.03.014. PMID 23542000.
- ^ 어떻게 복어가 이상한 가시를 얻게 되었는지 2019년 7월 25일.
- ^ Shono, T.; Thiery, A.P.; Cooper, R.L.; Kurokawa, D.; Britz, R.; Okabe, M.; Fraser, G.J. (2019). "Evolution and Developmental Diversity of Skin Spines in Pufferfishes". iScience. 19: 1248–1259. doi:10.1016/j.isci.2019.06.003. PMC 6831732. PMID 31353167.
- ^ Coolidge E, Hedrick MS 및 Milsom WK(2011) "Ventilatory Systems". 인: McKenzie DJ, Farrell AP, Brauner CJ (Eds) 물고기 생리학: 원시 물고기, 엘스비에, 182–213페이지. ISBN 9780080549521
- ^ Rothschild, Anna (1 April 2013). "Hagfish slime: The clothing of the future?". BBC News. Retrieved 2 April 2013.
- ^ Yong, Ed (23 January 2019). "No One Is Prepared for Hagfish Slime". The Atlantic. Retrieved 26 January 2019.
- ^ Friel, J P; Lundberg, J G (1996). "Micromyzon akamai, gen. et sp. nov., a small and eyeless banjo catfish (Siluriformes: Aspredinidae) from the river channels of the lower Amazon basin". Copeia. 1996 (3): 641–648. doi:10.2307/1447528. JSTOR 1447528.
- ^ Sadovy, Y.; Randall, J. E.; Rasotto, Maria B. (May 2005). "Skin structure in six dragonet species (Gobiesociformes; Callionymidae): Interspecific differences in glandular cell types and mucus secretion". Journal of Fish Biology. 66 (5): 1411–1418. doi:10.1111/j.0022-1112.2005.00692.x.
- ^ 참치는 비늘이 있니? 북동 수산 과학 센터 NOAA 어업. 2019년 8월 4일에 접속.
- ^ 레비티쿠스 11:9–10
- ^ Jump up to: a b c 아례 씨트론, "코셔 피쉬에 관한 모든 것"
- ^ Kosher Fish OU Kosher 인증 확인. 2019년 8월 9일 회수.
- ^ Jump up to: a b Lee, H. J.; Kusche, H.; Meyer, A. (2012). "Handed Foraging Behavior in Scale-Eating Cichlid Fish: Its Potential Role in Shaping Morphological Asymmetry". PLOS ONE. 7 (9): e44670. Bibcode:2012PLoSO...744670L. doi:10.1371/journal.pone.0044670. PMC 3435272. PMID 22970282.
- ^ Froese, R. and D. Pauly. Editors. "Glossary: Lepidophagy". FishBase. Retrieved 12 April 2007.
- ^ Jump up to: a b c Janovetz, Jeff (2005). "Functional morphology of feeding in the scale-eating specialist Catoprion mento" (PDF). The Journal of Experimental Biology. 208 (Pt 24): 4757–4768. doi:10.1242/jeb.01938. PMID 16326957. S2CID 15566769.
- ^ Martin, C.; P.C. Wainwright (2011). "Trophic novelty is linked to exceptional rates of morphological diversification in two adaptive radiations of Cyprinodon pupfish". Evolution. 65 (8): 2197–2212. doi:10.1111/j.1558-5646.2011.01294.x. PMID 21790569. S2CID 23695342.
- ^ Hori, M. (1993). "Frequency-dependent natural selection in the handedness of scale-eating cichlid fish". Science. 260 (5105): 216–219. Bibcode:1993Sci...260..216H. doi:10.1126/science.260.5105.216. PMID 17807183. S2CID 33113282.
- ^ Stewart, T. A.; Albertson, R. C. (2010). "Evolution of a unique predatory feeding apparatus: functional anatomy, development and a genetic locus for jaw laterality in Lake Tanganyika scale-eating cichlids". BMC Biology. 8 (1): 8. doi:10.1186/1741-7007-8-8. PMC 2828976. PMID 20102595.
추가 읽기
- Helfman, G.S., B.B. Collette and D.E. Facey (1997). The Diversity of Fishes. Blackwell Science. pp. 33–36. ISBN 978-0-86542-256-8.CS1 maint: 여러 이름: 작성자 목록(링크)
- Schultze, H.P. (2016). "Scales, enamel, cosmine, ganoine, and early osteichthyans". Comptes Rendus Palevol. 15 (1–2): 83–102. doi:10.1016/j.crpv.2015.04.001.

외부 링크
| 위키미디어 커먼즈에는 피쉬 스케일과 관련된 미디어가 있다. |
