피쉬지느러미
Fish fin.png)
지느러미는 Actinopterygii와 Chondrichthyes 물고기의 몸에서 튀어나온 가시나 가오리로 구성된 독특한 해부학적 특징입니다.그것들은 피부로 덮여 있고 대부분의 경골 어류에서 볼 수 있는 것처럼 물갈퀴가 있는 방식으로 결합되어 있거나 상어에서 볼 수 있는 것처럼 지느러미와 유사합니다.꼬리나 꼬리지느러미를 제외하고, 물고기 지느러미는 척추와 직접적인 연결이 없고 근육으로만 지탱됩니다.그들의 주된 기능은 물고기가 헤엄칠 수 있도록 도와주는 것입니다.
동물의 다른 위치에 위치한 지느러미는 앞으로 나아가거나, 돌아서거나, 똑바로 서 있거나, 멈추거나 하는 다양한 목적을 수행합니다.대부분은 수영할 때 지느러미를 사용하고, 날치는 가슴지느러미를 사용하여 활공하고, 개구리는 기어 다닐 때 사용합니다.지느러미는 다른 목적으로도 사용될 수 있습니다; 수컷 상어와 모기치는 정자를 전달하기 위해 변형된 지느러미를 사용하고, 탈곡기 상어는 먹이를 기절시키기 위해 꼬리 지느러미를 사용하고, 암초 돌 물고기는 독을 주입하는 가시를 등지느러미에 가지고 있고, 아귀는 먹이를 유인하기 위해 낚싯대처럼 등지느러미의 첫번째 척추를 사용합니다.그리고 트리거피쉬는 산호 틈에 쪼여들고, 지느러미의 가시를 이용하여 자신을 제자리에 가둠으로써 포식자를 피합니다.
핀 종류
핀은 쌍으로 구성할 수도 있고 쌍으로 구성할 수도 없습니다.가슴지느러미와 골반지느러미는 짝을 이루지만 등지느러미와 항문지느러미, 꼬리지느러미는 짝을 이루지 않고 몸의 중간선을 따라 위치합니다.모든 종류의 지느러미에 대해, 진화하는 동안 이 특정 지느러미가 사라진 많은 어종이 있습니다(예: 보바사트라니아의 골반 지느러미, 바다 개복치의 꼬리 지느러미).일부 분류에서는 진화 과정에서 짝을 이루지 못한 지느러미가 추가로 발견되기도 했습니다(예: 추가 등지느러미, 아디포스 지느러미).일부 아칸토디("가시상어")에서는 가슴지느러미와 골반지느러미 사이에 하나 이상의 "중간" 또는 "전골" 척추 쌍이 존재하지만, 이들은 지느러미와 관련이 없습니다.
| 가슴지느러미 |  | 한 쌍의 가슴지느러미는 양쪽에 위치하며, 보통 오퍼큘럼 바로 뒤에서 접힌 상태로 유지되며, 사지동물의 앞다리와 상동입니다.
|
|---|---|---|
| 골반지느러미 (ventral fin) |  | 한 쌍의 골반 또는 복측 지느러미는 일반적으로 가슴지느러미 아래와 뒤에 위치하지만, 많은 어족에서는 가슴지느러미(예: 대구) 앞에 위치할 수 있습니다.그들은 사지동물의 뒷다리와 상동입니다.골반지느러미는 물고기가 물을 통해 오르락내리락하고, 급격하게 회전하고, 빠르게 멈출 수 있도록 도와줍니다. |
| 등지느러미 |  상어등지느러미 |  등지느러미는 등 쪽에 위치합니다.물고기는 등지느러미를 3개까지 가질 수 있습니다.등지느러미는 구르는 것으로부터 물고기를 보호하고 갑작스러운 회전과 정지를 돕는 역할을 합니다. |
| 항문/클로아칼 지느러미 |  | 항문/클로아칼 핀은 항문/클로아카 뒤쪽 복측 표면에 위치합니다. |
| 지방지느러미 |  송어지느러미 | 지방지느러미는 등지느러미 뒤쪽과 꼬리지느러미 바로 앞에서 발견되는 부드럽고 살이 많은 지느러미입니다.많은 어족에서는 발견되지 않지만 31개의 유테오스테안목 중 9개 목(Percopsiformes, Myctophormes, Aulopiformes, Stomiiformes, Salmiformes, Osmeriformes, Characiformes, Siluriformes, Argentiniformes)에서 발견됩니다.[3]연어, 차라시드, 메기 등이 대표적입니다. 지방지느러미의 기능은 신비로운 것입니다.2005년의 자료에 따르면 지방지느러미를 제거한 송어의 꼬리박 빈도가 8% 더 높은 것으로 나타났지만, 부화한 물고기를 표시하기 위해 자주 잘라냅니다.[4][5]2011년에 발표된 추가적인 정보는 그 지느러미가 촉각, 소리 그리고 압력의 변화와 같은 자극을 감지하고 반응하는데 필수적일 수 있다는 것을 암시했습니다.캐나다 연구원들은 지느러미에서 신경망을 확인했는데, 이는 그것이 감각 기능을 가지고 있을 가능성이 있지만, 그것을 제거하는 것의 결과가 정확히 무엇인지는 아직도 확신하지 못하고 있습니다.[6][7] 2013년의 비교 연구에 따르면 지방 지느러미는 두 가지 다른 방식으로 발달할 수 있습니다.하나는 연어형 방식으로, 지방지느러미가 유충-지느러미 접힘부에서 동시에 그리고 다른 중앙지느러미와 동일하게 직접적으로 발달하는 방식입니다.다른 하나는 유충-지느러미 접힘이 줄어들고 다른 중앙지느러미가 발달한 후 지방지느러미가 늦게 발달하는 차라시폼(characiform)형 방식입니다.그들은 차라시폼 형태의 발달이 존재한다는 것은 지방지느러미가 "유충의 지느러미 접힌 나머지"가 아니라는 것을 암시하며 지방지느러미의 기능이 부족하다는 견해와 일치하지 않는다고 주장합니다.[3] 2014년 발표된 연구에 따르면 지방지느러미는 다른 계통에서 반복적으로 진화해왔습니다.[8] |
| 꼬리지느러미 (꼬리지느러미) |   하반신발지느러미 (Heterocercal caudal fin)  꼬리지느러미 (C) | 꼬리지느러미는 꼬리지느러미(꼬리를 뜻하는 라틴어 cauda에서 유래)로 꼬리지느러미의 끝에 위치합니다.대부분의 택시에서 추진에 사용됩니다(몸쪽 꼬리 지느러미 운동도 참조).꼬리지느러미는 축방향 골격의 척추뼈와 족저공(직경)에 의해 지지됩니다.축 골격과의 관계에 따라 4가지 유형의 꼬리지느러미(A-D)가 구별됩니다. (A) - 헤테로시컬(Heterocercal)은 척추뼈가 꼬리의 윗쪽 엽까지 확장되는 것을 의미하며, 종종 아랫쪽 엽보다 더 길게 만듭니다 (상어, 플라코더미, 대부분의 줄기 Actinopterygii, 철갑상어와 패들피시).그러나, 이종각 꼬리 지느러미의 외형은 대칭적으로 보일 수도 있습니다(예: Birgeria, Bobasatrania).자궁경부는 자궁경부의 반대입니다.
(B) - 원추는 척추뼈가 꼬리 끝까지 뻗어 있고 꼬리는 대칭적이지만 확장되지는 않는 것을 의미합니다 (첫 번째 물고기와 사이클로스톰처럼, 랜슬릿에서 더 원시적인 전구물질입니다). (C) - 일반적으로 지느러미가 표면적으로 대칭적으로 보이지만 실제로 척추뼈가 지느러미의 상엽까지 매우 짧은 거리만큼 뻗어 있는 동족골.그러나 동족지느러미는 비대칭적으로 보일 수도 있습니다(예: 푸른 날치).대부분의 현생 어류(teleost)는 동족의 꼬리를 가지고 있습니다.다양한 모양으로 제공되며 다음과 같이 나타날 수 있습니다. (D) - 두배골이란 척추뼈가 꼬리 끝까지 뻗어 있고 꼬리는 대칭적이고 확장되어 있는 것을 의미합니다(비치르, 폐어, 램프리, 실러캔스, 타라시상과 같이).대부분의 고생대 어류들은 이종의 2배체 꼬리를 가지고 있었습니다.[11]
|
| 카우달 용골 핀틀릿 | _diagram_cropped.GIF) | 빠르게 헤엄치는 어떤 종류의 물고기는 꼬리지느러미 바로 앞에 수평의 꼬리지느러미를 가지고 있습니다.배의 용골과 마찬가지로, 이것은 꼬리 지느러미에 안정성과 지지력을 제공하는 꼬리(아래 참조)로 구성된 꼬리 지느러미의 측면 능선입니다.한 쌍의 용골이 각 면에 하나씩 있을 수도 있고 위와 아래에 두 쌍이 있을 수도 있습니다. 지느러미는 작은 지느러미로, 일반적으로 등지느러미와 항문지느러미 뒤에 있습니다(비처에서는 등지느러미만 있고 등지느러미는 없습니다).참치나 꽁치와 같은 일부 물고기에서는 가오리가 없고, 물러설 수 없으며, 마지막 등지느러미와 항문지느러미 사이에서 발견됩니다. |
경골어류

골어류(Actinopterygii, Sarcopterygii)는 Osteichthyes(또는 육지 척추동물도 포함하는 Euteleostomi)라고 불리는 분류군을 형성합니다; 그들은 대부분 뼈로 만들어진 골격을 가지고 있고, (그들의 이빨, 지느러미 가시, 그리고 상아질을 제외하고) 주로 연골로 만들어진 골격을 가지고 있는 연골어류(아래 참조)와 대조될 수 있습니다.
경골 어류는 가오리 지느러미와 엽지느러미로 나뉩니다.대부분의 살아있는 물고기들은 30,000 종 이상으로 구성된 매우 다양하고 풍부한 집단인 가오리 지느러미를 가지고 있습니다.오늘날 현존하는 척추동물 중 가장 큰 부류로 종의 50% 이상을 차지하고 있습니다.[13]먼 옛날에는, 엽지느러미 물고기가 많았습니다.오늘날 그들은 단지 8종의 살아있는 종들과 함께, 빈곤한 다양성을 보여주고 있습니다.
경골 어류는 레피도트리치아라고 불리는 지느러미와 가오리를 가지고 있습니다.그들은 일반적으로 지느러미를 사용할 필요 없이 물고기가 가라앉는 것과 떠다니는 것 사이에서 중립적인 균형을 이루도록 해주는 수영 물갈퀴를 가지고 있습니다.그러나, 수영 물집은 많은 물고기, 특히 폐어에 존재하지 않는데, 이 물고기들은 수영 물집이 진화한 경골 어류의 공통 조상에 존재하는 원시적인 폐를 보유한 유일한 물고기입니다.경골 어류들은 또한 헤엄치기 위해 지느러미를 사용할 필요 없이 숨을 쉴 수 있도록 도와주는 견갑골을 가지고 있습니다.
엽지느러미



엽지느러미를 가진 물고기는 Sarcopterygii라고 불리는 뼈가 많은 물고기의 한 종류를 형성합니다.그들은 일련의 뼈로 몸에 결합된 살이 많고, 엽상적이며 쌍을 이루는 지느러미를 가지고 있습니다.[14]엽지느러미를 가진 물고기의 지느러미는 몸에서 뻗어 나온 살이 많고 엽처럼 생긴 비늘 모양의 줄기 위에서 자란다는 점에서 다른 모든 물고기의 지느러미와 다릅니다.가슴지느러미와 골반지느러미는 사지동물과 비슷한 관절을 가지고 있습니다.이 지느러미들은 데본기에 최초의 사지 척추동물들(양서류)의 다리로 진화했습니다.또한 대부분의 가오리 지느러미 물고기들의 등지느러미가 하나인 것과는 달리, 사르코테리기인들은 별도의 밑부분이 있는 두 개의 등지느러미를 가지고 있습니다.꼬리지느러미는 이종간질(화석 분류군만 해당) 또는 이중간질입니다.
라티메리아(Latimeria)는 아직까지 존재하는 지느러미가 있는 물고기입니다.서인도양 실러캔스(Latimeria chalumnae)와 인도네시아 실러캔스(Latimeria menadoensis) 두 종이 있습니다.실러캔스는 데본기 초기인 약 4억 800만 년 전에 대략 현재의 형태로 진화한 것으로 생각됩니다.[15]
실러캔스의 이동은 그들 종류에 독특합니다.실러캔스는 이동하기 위해 가장 일반적으로 물살과 표류의 위나 아래를 이용합니다.그들은 물 속에서 움직임을 안정화시키기 위해 짝을 지은 지느러미를 사용합니다.해저에서 그들의 짝을 지은 지느러미는 어떤 종류의 움직임에도 사용되지 않습니다.실러캔스는 꼬리 지느러미를 사용함으로써 빠른 출발을 위한 추진력을 만들 수 있습니다.그들이 가지고 있는 많은 지느러미 때문에, 실러캔스는 높은 기동성을 가지고 있고 물 속에서 그들의 몸을 거의 모든 방향으로 향하게 할 수 있습니다.그들은 머리를 세우고 배를 위로 헤엄치는 것이 목격되었습니다.그들의 로스트랄 기관은 실러캔스가 장애물 주변을 이동하는 것을 돕는 전기 지각을 하도록 돕는 것으로 생각됩니다.[16]
폐어는 살아있는 엽지느러미 물고기이기도 하답니다.그들은 아프리카 (Protopterus), 호주 (Neoceratodus), 그리고 남아메리카 (Lepidosien)에서 발생합니다.폐어는 데본기 시대에 진화했습니다.유전학적 연구와 고생물학적 자료는 폐어가 육상 척추동물의 가장 가까운 살아있는 동족이라는 것을 확인시켜줍니다.[17]
지느러미 모양과 지느러미 모양은 지느러미 물고기에서 비교적 보수적입니다.그러나, Allenypterus, Rebelligatrix, Foreyia 또는 사족류와 같은 이상한 형태를 보여주는 화석 기록의 몇 가지 예가 있습니다.
지느러미 어류의 지느러미 다양성
-

-

점박이 폐어 Protopterus dolloi
-
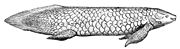
퀸즐랜드폐어 네오케라토더스 포스테리
-

-

-

-

-

-

서인도양 실러캔스 라티메리아 칼룸나에
-

_by_ObsidianSoul_02_flipped.png)





_(cropped).jpg)

.jpg)
가오리핀

가오리 지느러미를 가진 물고기는 Actinopterygii라고 불리는 뼈가 많은 물고기의 한 종류입니다.그들의 지느러미에는 가시나 가오리가 들어있습니다.지느러미는 가시광선, 연광선, 또는 둘의 조합만을 포함할 수 있습니다.만약 둘 다 존재한다면, 가시광선은 항상 앞쪽에 있습니다.가시는 일반적으로 뻣뻣하고 날카롭습니다.광선은 일반적으로 부드럽고 유연하며 분절적이며 가지가 있을 수 있습니다.광선의 이러한 분절은 가시와 가시를 구분하는 주요한 차이점입니다. 가시는 특정 종에서 유연할 수 있지만 결코 분절되지 않습니다.
척추는 다양한 용도를 가지고 있습니다.메기에서, 그들은 방어의 한 형태로 사용됩니다; 많은 메기들은 그들의 가시를 바깥쪽으로 잠글 수 있는 능력을 가지고 있습니다.방아쇠 물고기는 또한 가시가 뽑히는 것을 막기 위해 틈에 가두는 데 사용합니다.
레피도트리샤는 보통 뼈로 구성되어 있지만, Cheirolepis와 같은 초기 골인류에서는 상아질과 에나멜도 있었습니다.[18]이러한 디스크는 분할되어 여러 디스크 위에 쌓여 있습니다.그것들은 피부 비늘에서 유래되었을 수도 있습니다.[18]지느러미 선의 형성에 대한 유전적 근거는 특정 단백질의 생성을 위해 코딩된 유전자로 생각됩니다.엽지느러미 물고기에서 사지동물의 진화는 이러한 단백질의 손실과 관련이 있다고 제안되어 왔습니다.[19]
가오리지느러미 어류의 지느러미 다양성
-
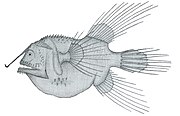
팬핀 낚시꾼 코로프리네 조르다니
-

팬케이크 배트피시 Halieutichys aculeatus
-

가느다란 개복치 란자니아 레비스
-

팬피쉬 카롤리누스속
-

무화과 해초 스테놉틱스 디아파나
-

Silverroughy Hoplostethus mediteraneus
-

볏가자미 로포넥테스갈루스
-

잭나이프피쉬 에쿠스 란솔라투스속
-

-

대서양난파치 폴리프리온아메리카누스
-

-
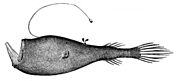
별을 바라보는 바다뱀붙이 세라티아 수라노스코프
-

리지헤드 뽀로미트라 유니콘스
-

열대양날개날다귀 외떡잎코끼리
-

쿠스크장어 벤토코메테스로부스투스
-

래트테일 술카투스속
-

트라이포드피쉬 배시프테로이스 그릴레이터
-

대왕노랑어 리갈레쿠스 글레스네
-

짧은부리저어 안구스티로스트리스
-

고스트나이프피쉬 스텔라르코린쿠스 옥시린쿠스
-

리모라 레모라브라히프테라
-

블루대시락스키퍼 근막골절도마뱀붙이
-

나일비치르 폴립테루스 비치르
-

해안절개송어 온코린쿠스 클라키
-

아프리카버터메기 실베미스터스
-

붕장어 렙토케팔루스 붕장어


.gif)



.gif)











.gif)
.gif)





연골어류


연골어류는 Chondrichthyes라고 불리는 물고기의 한 종류를 형성합니다.그들은 뼈가 아닌 연골로 만들어진 골격을 가지고 있습니다.이 수업에는 상어, 가오리, 키메라 등이 포함됩니다.
상어 지느러미 골격은 길쭉하고, 머리카락과 깃털의 각질을 닮은 탄력성 있는 단백질 필라멘트인 세라토트리치아(ceratotricia)라는 이름의 부드럽고 분절되지 않은 광선으로 지탱됩니다.[20]원래 피부 성분이 없는 가슴 거들과 골반 거들은 연결되지 않았습니다.나중의 형태에서, 스카풀로코라코이드와 치골봉이 진화했을 때, 각 지느러미 쌍은 중간에 양쪽으로 연결되었습니다.광선 속에서 가슴지느러미는 머리에 연결되어 있고 매우 유연합니다.대부분의 상어에게 존재하는 주요 특징 중 하나는 이동을 돕는 이종의 꼬리입니다.[21]대부분의 상어는 8개의 지느러미를 가지고 있습니다.상어들은 지느러미가 꼬리의 첫 번째 방향으로 움직이는 것을 허용하지 않기 때문에 상어들의 바로 앞에 있는 물체들로부터만 떠내려갈 수 있습니다.[22]
현대의 연골어류와는 달리, 줄기 연골어류(예: 클라이마티스과 디플라칸트과 같은)[23]의 구성원들은 가슴의 진피판과 쌍지느러미와 관련된 진피판을 가지고 있었습니다.이러한 특징을 보여주는 가장 오래된 종은 중국의 실루리안(에어로니안) 하류 지역에서 발견된 아칸토디안 판징샤니아 리노바타입니다[24].판징샤니아는 뼈판에 융합된 진피 비늘로 구성된 복합 가슴판과 뼈로 이루어진 지느러미 가시를 가지고 있습니다.등지느러미와 관련된 지느러미는 현존하는 연골어류 중에서는 드물지만, 예를 들어 헤테로돈투스(Heterodontus)나 스쿠알루스(Squalus)에 존재합니다.등지느러미 가시는 일반적으로 Hybodontiformes, Ctenacanthiformes 또는 Xenacanthida와 같은 많은 화석군에서 발달합니다.스테타칸투스에서, 척추-붓 복합체를 형성하면서, 첫 번째 등지느러미 척추가 변형되었습니다.
대부분의 물고기와 마찬가지로, 상어의 꼬리는 꼬리 모양에 따라 속도와 가속도를 좌우하며 추진력을 제공합니다.고래 지느러미 모양은 상어 종마다 상당히 다양한데, 이는 별도의 환경에서 진화하기 때문입니다.상어는 배쪽이 배쪽보다 눈에 띄게 큰 이종성 꼬리지느러미를 가지고 있습니다.이것은 상어의 척추가 등 쪽으로 뻗어 있어서 근육 부착을 위한 더 넓은 표면적을 제공하기 때문입니다.이것은 이 부정적으로 부력이 있는 연골어류들 사이에서 더 효율적인 이동을 가능하게 합니다.이와는 대조적으로, 대부분의 경골 어류는 동족지느러미를 가지고 있습니다.[25]
호랑이 상어는 큰 윗엽을 가지고 있는데, 이것은 천천히 순항하고 갑자기 속도를 낼 수 있게 해줍니다.호랑이 상어는 사냥을 할 때 물을 쉽게 비틀고 돌릴 수 있어야 다양한 먹이를 유지할 수 있습니다. 반면 고등어와 청어와 같은 학교에서 공부하는 물고기를 사냥하는 포비글 상어는 빠르게 헤엄치는 먹이와 보조를 맞추기 위해 큰 아래엽을 가지고 있습니다.[13]다른 꼬리 적응은 상어가 물고기와 오징어를 기절시키기 위해 힘있고 길쭉한 윗엽을 사용하는 것과 같이 더 직접적으로 먹이를 잡는 것을 도와줍니다.
반면에, 광선은 추진력을 위해 커진 가슴지느러미에 의존합니다.비슷한 크기로 커진 가슴지느러미는 멸종된 Petalodontiformes (예: Belantsea, Janassa, Menaspis), Holocephali (쥐치와 그 화석 동족) 또는 Aquilolamna (Selachimorpha)와 Squatinactis (Squatinactiformes)에서 발견됩니다.어떤 연골어류는 뱀장어와 같은 운동을 합니다. (예: Clamydoselachus, Thrinacoselache,[26] Phoebodus[27])
연골어류의 지느러미 다양성
-

작은점박이두툽상어 Skyliorhinus canicula
-

백상아리 카르카로돈 카르카리아스
-

-

큰 이빨톱어 Pristis peroteti
-

대리석 전기선 어뢰 마모라타
-

베넷의 가오리 헤미트리곤 베넷디
-

-

-

스테타칸투스속("Stethacanthus productus")은 미국선녀벌레과에 속하는 속씨식물 속입니다.
-

-

-
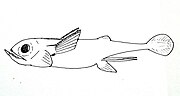
-

쿠바키메라 키마에라 쿠바나
-

미국코끼리물고기 Callorhinchus callorhynchus

.png)












상어지느러미
Human [28]Society International에 따르면, 상어 지느러미 때문에 매년 약 1억 마리의 상어가 죽임을 당합니다.지느러미가 잘린 후, 훼손된 상어들은 다시 물에 던져지고 죽게 내버려집니다.
아시아의 몇몇 나라에서는 상어 지느러미 수프와 같이 상어 지느러미가 요리의 진미입니다.[29]현재 상어의 지속가능성과 복지에 대한 국제적인 우려가 전세계적으로 상어지느러미 수프의 소비와 이용가능성에 영향을 미치고 있습니다.[30]상어 지느러미를 잡는 것은 많은 나라에서 금지되어 있습니다.
핀함수
추력발생
호일 모양의 핀은 움직일 때 추력을 발생시키고, 핀의 양력은 물이나 공기를 움직이게 하고 핀을 반대 방향으로 밀어냅니다.수중 동물들은 물속에서 지느러미를 앞뒤로 움직여서 상당한 추진력을 얻습니다.종종 꼬리지느러미가 사용되지만, 일부 수생 동물들은 가슴지느러미로부터 추진력을 발생시킵니다.[31]

공동 현상은 음의 압력으로 인해 액체에 기포(공동)가 형성되어 빠르고 격렬하게 붕괴될 때 발생합니다.큰 손상과 마모를 일으킬 수 있습니다.[32]돌고래와 참치와 같은 강력한 수영 해양 동물의 꼬리 지느러미에 공동 피해가 발생할 수 있습니다.캐비테이션은 주변 수압이 상대적으로 낮은 바다 표면 근처에서 발생할 가능성이 더 높습니다.돌고래들이 더 빨리 수영할 수 있는 힘이 있다 하더라도, 꼬리에 있는 붕괴하는 공동 거품이 너무 고통스럽기 때문에 돌고래들은 속도를 제한해야 할지도 모릅니다.[33]캐비테이션은 참치의 속도를 늦추기도 하지만, 다른 이유가 있습니다.돌고래와 달리, 이 물고기들은 신경의 끝이 없는 뼈가 있는 지느러미를 가지고 있기 때문에 거품을 느끼지 못합니다.그럼에도 불구하고, 그들은 지느러미 주변에 속도를 제한하는 증기 피막을 형성하기 때문에 더 빨리 수영할 수 없습니다.참치에서 공동 손상과 일치하는 병변이 발견되었습니다.[33]
스컴브리드 물고기(참치, 고등어, 가다랑어)는 특히 뛰어난 수영 실력을 자랑합니다.그들의 몸 뒤쪽 가장자리를 따라서 작은 선이 없고, 후퇴할 수 없는 지느러미들이 늘어서 있는데, 이를 핀릿이라고 합니다.이 핀틀의 기능에 대해서는 많은 추측이 있었습니다.나우엔과 로더가 2000년과 2001년에 수행한 연구는 "지느러미들이 꾸준한 수영을 하는 동안 국부적인 흐름에 유체역학적인 영향을 미친다"와 "가장 뒤에 있는 지느러미들은 수영하는 고등어의 꼬리에 의해 생성되는 추진력을 증가시킬 수 있는 꼬리 소용돌이로 흐름을 전환하도록 설계되어 있습니다"[34][35][36]라고 말했습니다.
물고기는 여러 개의 지느러미를 사용하기 때문에 주어진 지느러미가 다른 지느러미와 유체역학적 상호작용을 할 수 있습니다.특히, 코달(꼬리) 핀의 바로 상류에 있는 핀은 코달 핀에서의 유동 역학에 직접적인 영향을 줄 수 있는 근접 핀일 수 있습니다.2011년, 체적 영상 기술을 사용한 연구자들은 "자유롭게 헤엄치는 물고기들에 의해 생성되는 기상 구조의 최초의 순간적인 3차원 뷰"를 생성할 수 있었습니다.그들은 "연속적인 꼬리의 박자가 소용돌이 고리의 연결된 사슬을 형성하는 결과를 가져왔다"는 것과 "등지느러미와 항문지느러미의 각성은 대략 다음 꼬리 박자의 시간 내에 꼬리지느러미의 각성에 의해 빠르게 진행된다"는 것을 발견했습니다.[37]
제어동작
동작이 확립되면 다른 지느러미를 사용하여 동작 자체를 제어할 수 있습니다.[31][38]


.jpg)
암초 물고기의 몸은 종종 개복치와는 다른 모양을 하고 있습니다.오픈 워터 피쉬는 보통 속도를 위해 만들어지며, 물을 통과할 때 마찰을 최소화하기 위해 어뢰처럼 유선형으로 만들어집니다.산호초의 상대적으로 좁은 공간과 복잡한 수중 경관에서 암초 물고기가 활동합니다.이러한 기동성은 직선 속도보다 더 중요하기 때문에 산호초 물고기들은 활쏘기와 방향 전환 능력을 최적화하는 몸을 개발했습니다.그들은 산호초의 틈으로 피하거나 숨어서 산호초 주위를 찾음으로써 포식자들을 능가합니다.[42]나비 물고기, 담쟁이 물고기, 천사 물고기와 같은 많은 암초 물고기의 가슴과 골반 지느러미는 브레이크 역할을 하고 복잡한 기동을 할 수 있도록 진화했습니다.[44]나비 물고기, 담쟁이 물고기 그리고 천사 물고기와 같은 많은 암초 물고기들은 팬케이크처럼 깊고 옆으로 압축된 진화된 몸을 가지고 있고 바위의 틈에 들어갈 것입니다.골반과 가슴지느러미는 서로 다르게 진화했기 때문에 납작해진 몸과 함께 작동하여 기동성을 최적화합니다.[42]복어, 쥐치, 트렁크피쉬와 같은 몇몇 물고기들은 수영을 할 때 가슴지느러미에 의지하고 꼬리지느러미를 거의 사용하지 않습니다.[44]
생식


수컷 연골어류(상어와 가오리)와 일부 살아있는 가오리 지느러미 물고기의 수컷은 내부 수정이 가능한 생식 부속물인 침입 기관의 기능을 하도록 변형된 지느러미를 가지고 있습니다.가오리 지느러미 물고기에서는 고노포디아 또는 수각류라고 불리고, 연골어류에서는 클러퍼라고 불립니다.
고노포디아는 Anablepidae과와 Poeciliidae과의 일부 종의 수컷에서 발견됩니다.그것들은 움직일 수 있는 침입 기관의 기능을 하도록 변형된 항문 지느러미이며, 교미 중에 암컷에게 젖을 물리는 데 사용됩니다.수컷 항문지느러미의 세 번째, 네 번째, 다섯 번째 광선은 물고기의 정자가 분출되는 관 모양의 구조로 형성됩니다.[47]짝짓기를 할 준비가 되었을 때, 고노포디움은 직립하고 암컷을 향합니다.수컷은 암컷의 성구에 기관을 곧 삽입하는데, 물고기가 암컷을 붙잡아 임신을 확실히 할 수 있도록 하는 갈고리와 같은 적응을 합니다.만약 암컷이 움직이지 않고 있다가 그녀의 파트너가 그의 고노포디움과 그녀의 통풍구를 접촉한다면, 그녀는 수정될 것입니다.정자는 암컷의 난관에 보존됩니다.이것은 암컷들이 수컷들의 더 이상의 도움 없이 언제든지 스스로 수정할 수 있도록 해줍니다.어떤 종들에서는, 고노포디움이 전체 몸길이의 절반일 수도 있습니다.때로 지느러미가 너무 길어서 사용할 수 없을 때도 있는데, "리테일"종의 시포포루스 헬레리처럼 말입니다.호르몬 치료를 받은 여성들은 생식기계증에 걸릴 수도 있습니다.이것들은 번식에 쓸모가 없습니다.
비슷한 특징을 가진 유사한 기관은 다른 물고기에서 발견되는데, 예를 들어 헤미르함포돈의 안드로포디움이나 구데이대[48] 혹은 미들 트라이아스기 사우리치티스의 고노포디움은 가오리 지느러미 물고기의 생체성의 가장 오래된 예입니다.[49]
쥐는 것은 연골어류의 수컷에서 발견됩니다.그것들은 골반 지느러미의 뒤쪽 부분으로 또한 침범 기관의 기능을 하도록 변형되었고, 교배 동안 정액을 암컷의 클로카로 보내는 데 사용됩니다.상어의 짝짓기 행위는 보통 특정한 오리피스를 통해 물이 사이펀으로 들어갈 수 있도록 하는 걸쇠 중 하나를 들어올리는 것을 포함합니다.클라스퍼는 클로카에 삽입되고, 클라스퍼는 우산처럼 열려 위치를 고정시킵니다.그러면 사이펀은 물과 정자를 내보내는 수축을 시작합니다.[50][51]
기타기능
지느러미의 다른 용도로는 걷기와 해저에 걸터앉기, 물 위를 미끄러지기, 체온을 식히기, 먹잇감을 놀라게 하기, 전시(포수의 겁주기, 구애), 방어(독소 지느러미, 산호 사이의 자물쇠), 먹잇감을 유혹하기, 부착 구조 등이 있습니다.
인도 태평양 돛새치는 두드러진 등지느러미를 가지고 있습니다.스컴브로이드나 다른 부리 물고기처럼, 그들은 수영을 할 때 등지느러미를 몸의 홈으로 밀어 넣음으로써 자신을 능률적으로 만듭니다.[52]돛새치의 거대한 등지느러미, 즉 돛은 대부분의 경우 뒤로 젖혀집니다.돛새치들은 작은 물고기 무리를 무리 지어 다니고 싶다면 그들을 기르며, 또한 아마도 열을 식히기 위해 활동 기간이 지난 후에 기릅니다.[52][53]
_in_Waikiki_Aquarium.JPG)




동양날다람쥐는 보통 몸에 고정하는 큰 가슴지느러미를 가지고 있고 포식자에게 위협을 받으면 확장합니다.그 이름에도 불구하고, 그것은 날치가 아닌 피하의 물고기이고, 바다의 바닥을 걸을 때 골반 지느러미를 사용합니다.[55][56]
지느러미는 성적 장식품으로서 적응적인 의미를 가질 수 있습니다.구애하는 동안 암컷 시클리드인 골반아크로미스 태니아투스는 크고 눈에 띄는 보라색 골반지느러미를 보여줍니다."연구원들은 수컷들이 분명히 더 큰 골반 지느러미를 가진 암컷을 더 좋아했고 골반 지느러미가 암컷 물고기의 다른 지느러미들보다 더 불균형적인 방식으로 자라났다는 것을 발견했습니다."[57][58]






진화
쌍지느러미의 진화
물고기에서 쌍지느러미의 진화 모델로 역사적으로 논의되어 온 두 가지 유력한 가설이 있습니다: 아가미 아치 이론과 측면 지느러미 접이 이론.일반적으로 "게겐바우어 가설"이라고 불리는 전자는 1870년에 확립되었으며 "쌍을 이룬 지느러미는 아가미 구조에서 파생된 것"이라고 제안합니다.[60]이것은 1877년에 처음으로 제안된 옆지느러미 접기 이론에 찬성하여 인기를 잃었는데, 이 이론은 한 쌍의 지느러미가 아가미 바로 뒤의 표피를 따라 세로, 가로로 접힌 부분에서 싹을 틔운다고 제안합니다.[61]화석 기록과 발생학에서 두 가설 모두에 대한 지지가 약합니다.[62]하지만, 발달 패턴에 의한 최근의 통찰은 짝을 이룬 지느러미의 기원을 더 잘 설명하기 위해 두 이론을 재고하게 만들었습니다.
고전이론
칼 게겐바우어의 개념인 "아키프테리움"은 1876년에 소개되었습니다.[63]그것은 아가미 아치에서 뻗어 나온 아가미, 즉 "관절 연골 줄기"로 묘사되었습니다.아치를 따라 그리고 중앙 아가미 광선에서 추가 광선이 발생했습니다.게겐바우르는 변형 상동성의 모델을 제시했습니다. – 모든 척추동물 쌍의 지느러미와 사지가 아치페테리움의 변형이라는 것입니다.이 이론에 근거하면 가슴지느러미와 골반지느러미와 같은 짝을 이룬 부속물은 가지형 아치와 구별되어 뒤쪽으로 이동했을 것입니다.그러나 형태적으로나 계통적으로 화석 기록에서 이 가설을 지지하는 것은 제한적이었습니다.[62]또한 골반 지느러미의 전-후 이동에 대한 증거는 거의 없었습니다.[64]아가미 아치 이론의 그러한 단점들은 성자에 의해 제안된 측면 지느러미 접기 이론을 선호하여 그것의 초기 종말로 이어졌습니다. 조지 잭슨 미바트, 프랜시스 밸푸어, 제임스 킹슬리 태처.
옆지느러미 접기 이론은 물고기의 몸 벽을 따라 옆으로 접힌 지느러미가 발달했다는 가설을 세웠습니다.[61]중앙 지느러미 주름의 분절과 싹이 중앙 지느러미를 발생시켰듯이, 옆 지느러미 주름으로부터의 유사한 핀 싹의 분절과 신장 메커니즘이 한 쌍의 가슴지느러미와 골반지느러미를 발생시켰다고 제안되었습니다.하지만, 화석 기록에는 옆으로 접힌 지느러미 모양의 전환에 대한 증거는 거의 없었습니다.[65]게다가, 가슴지느러미와 골반지느러미가 뚜렷한 진화론적, 기계론적 기원에서 나온다는 것이 나중에 계통발생적으로 증명되었습니다.[62]
진화발달생물학
짝을 이룬 동물의 존재와 진화에 대한 최근의 연구들은 램프리와 같은 지느러미가 없는 척추동물들과 짝을 이룬 지느러미가 있는 가장 기초적인 살아있는 척추동물인 연골무과를 비교했습니다.[66]2006년, 연구원들은 중앙 지느러미의 분절과 발달에 관련된 같은 유전적 프로그래밍이 두툽상어의 짝을 이루는 부속지의 발달에서도 발견되었습니다.[67]이러한 발견이 측면 지느러미 접기 가설을 직접적으로 지지하지는 않지만, 공유 중앙값 쌍을 이룬 지느러미 진화적 발달 메커니즘의 원래 개념은 여전히 관련성이 있습니다.
오래된 이론의 유사한 개조는 콘드리치안 아가미 아치와 쌍으로 된 부속물의 개발 프로그래밍에서 찾을 수 있습니다.2009년, 시카고 대학의 연구원들은 콘드리치안 아가미 아치와 짝을 이룬 지느러미의 초기 개발에 공유된 분자 패턴 메커니즘이 있다는 것을 증명했습니다.[68]이와 같은 연구 결과는 한 때 발견된 아가미 아치 이론을 재고하게 만들었습니다.[65]
지느러미에서 사지까지
물고기는 모든 포유류, 파충류, 조류 그리고 양서류의 조상입니다.[69]특히 육상 사지동물(네발 동물)은 어류에서 진화해 4억년 전 육지로 첫 진출을 했습니다.[70]그들은 이동을 위해 가슴지느러미와 골반지느러미가 한 쌍으로 형성된 것을 사용했습니다.가슴지느러미는 앞다리(인간의 경우 팔)로, 골반지느러미는 뒷다리로 발달했습니다.[71]사지동물에서 걷는 사지를 형성하는 유전자 기계의 많은 부분은 이미 물고기의 헤엄치는 지느러미에 존재합니다.[72][73]
아리스토텔레스는 유사 구조와 상동 구조의 구별을 인식하고 다음과 같은 예언적 비교를 했습니다: "새는 어떤 면에서 물고기를 닮았습니다.왜냐하면 새들은 몸의 윗부분에 날개를 가지고 있고 물고기들은 몸의 앞부분에 두개의 지느러미를 가지고 있기 때문입니다.새들은 아랫부분에 발이 있고 대부분의 물고기들은 아랫부분과 앞지느러미 근처에 두 번째 지느러미가 있습니다."
– Aristotle, De incessu animalium[74]


2011년 호주의 모나시 대학의 연구원들은 원시적이지만 여전히 살아있는 폐어를 "사족류의 하중을 받는 뒷다리가 어떻게 진화했는지를 알아내기 위해 골반 지느러미 근육의 진화를 추적하기 위해" 사용했습니다.[75][76]시카고 대학의 추가적인 연구는 바닥을 걷는 폐어류가 육상 사지동물의 걸음걸이의 특성을 이미 진화시켰다는 것을 발견했습니다.[77][78]
융합적 진화의 고전적인 예에서 익룡, 새, 박쥐의 가슴 팔다리는 독립적인 경로를 따라 하늘을 나는 날개로 더욱 진화했습니다.날갯짓을 해도 걷는 다리와 유사한 점이 많으며 가슴지느러미의 유전적 청사진의 핵심적인 측면이 남아 있습니다.[79][80]
최초의 포유류는 페름기(2억9890만년 전에서 2억5217만년 전 사이)에 출현했습니다.고래류(고래, 돌고래, 상괭이)를 포함한 이 포유류의 여러 그룹이 바다로 돌아가기 시작했습니다.최근 DNA 분석은 고래류가 짝수발 유제류 안에서 진화했으며 하마와 공통 조상을 공유하고 있음을 시사합니다.[81][82]약 2천 3백만년 전에 육지 포유동물과 비슷한 또 다른 그룹이 바다로 돌아오기 시작했습니다.이것들은 물개들이었습니다.[83]고래와 바다표범에서 걸어 다니는 수족이 된 것은 독립적으로 새로운 형태의 수영지느러미로 진화했습니다.앞다리는 물갈퀴가 된 반면 뒷다리는 유실되거나 물갈퀴로 변형되었습니다.고래류에서, 꼬리는 마지막에 두 개의 지느러미를 얻었는데, 이것은 요행이라고 불립니다.[84]물고기 꼬리는 보통 수직이고 좌우로 움직입니다.고래류의 가시는 다른 포유류와 같은 방식으로 구부러지기 때문에, 고래류는 수평적이고 위아래로 움직입니다.[85][86]

어룡은 돌고래와 닮은 고대 파충류입니다.그들은 약 2억 4천 5백만 년 전에 처음 나타났고 약 9천만 년 전에 사라졌습니다.
"지상의 조상을 가진 이 바다를 다니는 파충류는 물고기들에게 매우 강하게 수렴하여 실제로 향상된 수중 운동을 위해 등지느러미와 꼬리지느러미를 진화시켰습니다.이 구조물들은 무에서 진화했기 때문에 더욱 주목할 만합니다. 조상의 육상 파충류는 전구 역할을 하기 위해 등에 혹이나 꼬리에 날이 없었습니다."[87]
생물학자 스티븐 제이 굴드는 이 어룡이 그가 가장 좋아하는 수렴 진화의 예라고 말했습니다.[88]
다양한 형태와 다양한 위치(다리, 몸통, 꼬리)의 지느러미 또는 지느러미는 또한 펭귄(날개에서 변형된), 바다 거북(앞다리가 지느러미로 변형된), 모사사우르스(지느러미로 변형된), 바다 뱀(세로로 확장된 납작한 꼬리 지느러미)과 같은 다른 네발동물 그룹에서 진화했습니다.
로봇 지느러미

| 외부영상 | |
|---|---|
수중 동물의 추진을 위한 지느러미의 사용은 현저하게 효과적일 수 있습니다.어떤 물고기들은 90%[31] 이상의 추진 효율을 얻을 수 있다고 계산되었습니다.물고기는 배나 잠수함보다 훨씬 더 효과적으로 가속하고 조종할 수 있고 물의 교란과 소음을 덜 낼 수 있습니다.이것은 수중 동물의 움직임을 모방하려는 수중 로봇의 생체 모방 연구로 이어졌습니다.[90]예를 들면, 해면 운동을 분석하고 수학적으로 모델링하기 위해 필드 로보틱스 연구소에서 만든 로봇 참치가 있습니다.[91]2005년, 씨 라이프 런던 아쿠아리움은 에식스 대학의 컴퓨터 과학과에서 만든 세 마리의 로봇 물고기를 전시했습니다.그 물고기는 진짜 물고기처럼 주변을 헤엄치고 장애물을 피하면서 자율적으로 설계되었습니다.그들의 창작자는 그가 "참치의 속도, 파이크의 가속도, 그리고 뱀장어의 항해 기술"을 결합하려고 노력하고 있었다고 주장했습니다.[92][93][94]
독일의 페스토(Festo)가 개발한 아쿠아펭귄은 펭귄의 앞지느러미 모양과 추진력을 모방합니다.[95][96]Festo는 또한 AquaRay,[97] AquaJelly[98], AiraCuda를 각각 [99]만타선, 해파리, 바라크라의 움직임을 모방하여 개발했습니다.
2004년 MIT의 휴 허는 개구리 다리의 근육을 로봇에 외과적으로 이식한 다음 전기로 근육 섬유를 움직이면서 로봇을 수영하게 함으로써 살아있는 작동기를 가진 생체기계 로봇 물고기의 원형을 만들었습니다.[100][101]
로봇 물고기는 물고기 디자인의 개별 부분을 다른 물고기들과 분리하여 조사할 수 있는 능력과 같은 몇 가지 연구상의 이점을 제공합니다.하지만, 이것은 생물학을 지나치게 단순화시킬 위험이 있기 때문에 동물 디자인의 중요한 측면들은 간과되고 있습니다.로봇 물고기는 또한 연구원들이 유연성이나 특정 동작 제어와 같은 하나의 파라미터를 변화시킬 수 있게 해줍니다.연구원들은 직접 힘을 측정할 수 있는데, 이것은 살아있는 물고기에서 하기가 쉽지 않습니다."로봇 장치는 또한 기관차 표면의 위치를 정확하게 알 수 있기 때문에 3차원 운동학적 연구와 연관된 유체 역학 분석을 용이하게 합니다.또한 자연 운동의 개별 구성 요소(예를 들어, 돌출부 대 펄럭이는 부속물의 뇌졸중)는 별도로 프로그래밍될 수 있는데, 이것은 살아있는 동물과 작업할 때 확실히 달성하기 어렵습니다."[102]
참고 항목
참고문헌
인용문
- ^ Standen, EM (2009). "Muscle activity and hydrodynamic function of pelvic fins in trout (Oncorhynchus mykiss)". The Journal of Experimental Biology. 213 (5): 831–841. doi:10.1242/jeb.033084. PMID 20154199.
- ^ Gene Helfman, Bruce Collette, Douglas Facey, & Brian Bowen. (2009) 어류의 다양성: 생물학, 진화, 생태학존 와일리 앤 선즈.
- ^ a b Bender, Anke; Moritz, Timo (1 September 2013). "Developmental residue and developmental novelty – different modes of adipose-fin formation during ontogeny". Zoosystematics and Evolution. 89 (2): 209–214. doi:10.1002/zoos.201300007. ISSN 1860-0743.
- ^ Tytell, E. (2005). "The Mysterious Little Fatty Fin". Journal of Experimental Biology. 208 (1): v–vi. doi:10.1242/jeb.01391.
- ^ Reimchen, T E; Temple, N F (2004). "Hydrodynamic and phylogenetic aspects of the adipose fin in fishes". Canadian Journal of Zoology. 82 (6): 910–916. doi:10.1139/Z04-069.
- ^ Temple, Nicola (18 July 2011). "Removal of trout, salmon fin touches a nerve". Cosmos. Archived from the original on 12 January 2014.
- ^ Buckland-Nicks, J. A.; Gillis, M.; Reimchen, T. E. (2011). "Neural network detected in a presumed vestigial trait: ultrastructure of the salmonid adipose fin". Proceedings of the Royal Society B: Biological Sciences. 279 (1728): 553–563. doi:10.1098/rspb.2011.1009. PMC 3234561. PMID 21733904.
- ^ Stewart, Thomas A.; Smith, W. Leo; Coates, Michael I. (2014). "The origins of adipose fins: an analysis of homoplasy and the serial homology of vertebrate appendages". Proceedings of the Royal Society B: Biological Sciences. 281 (1781): 20133120. doi:10.1098/rspb.2013.3120. PMC 3953844. PMID 24598422.
- ^ Hyman, Libbie (1992). Hyman's Comparative Vertebrate Anatomy (3 ed.). The University of Chicago Press. p. 210. ISBN 978-0226870137. Retrieved 18 October 2018.
- ^ a b Brough, James (1936). "On the evolution of bony fishes during the Triassic Period". Biological Reviews. 11 (3): 385–405. doi:10.1111/j.1469-185X.1936.tb00912.x. S2CID 84992418.
- ^ 폰 지텔 KA, 우드워드 AS와 슐로저 M (1932) 고생물학 2권, 맥밀런과 회사의 교과서.13페이지.
- ^ Kogan , Romano (2016). "Redescription of Saurichthys madagascariensis Piveteau, 1945 (Actinopterygii, Early Triassic), with implications for the early saurichthyid morphotype". Journal of Vertebrate Paleontology. 36 (4): e1151886. Bibcode:2016JVPal..36E1886K. doi:10.1080/02724634.2016.1151886. S2CID 87234436.
- ^ a b Nelson, Joseph S. (1994). Fishes of the World. New York: John Wiley and Sons. ISBN 978-0-471-54713-6. OCLC 28965588.
- ^ Clack, J. A. (2002) Gaining Ground.인디애나 대학교
- ^ Johanson, Zerina; Long, John A.; Talent, John A.; Janvier, Philippe; Warren, James W. (2006). "Oldest Coelacanth, from the Early Devonian of Australia". Biology Letters. 2 (3): 443–46. doi:10.1098/rsbl.2006.0470. PMC 1686207. PMID 17148426.
- ^ Fricke, Hans; Reinicke, Olaf; Hofer, Heribert; Nachtigall, Werner (1987). "Locomotion of the Coelacanth Latimeria Chalumnae in Its Natural Environment". Nature. 329 (6137): 331–33. Bibcode:1987Natur.329..331F. doi:10.1038/329331a0. S2CID 4353395.
- ^ Takezaki, N.; Nishihara, H. (2017). "Support for lungfish as the closest relative of tetrapods by using slowly evolving ray-finned fish as the outgroup". Genome Biology and Evolution. 9 (1): 93–101. doi:10.1093/gbe/evw288. PMC 5381532. PMID 28082606.
- ^ a b Zylberberg, L.; Meunier, F. J.; Laurin, M. (2016). "A microanatomical and histological study of the postcranial dermal skeleton of the Devonian actinopterygian Cheirolepis canadensis". Acta Palaeontologica Polonica. 61 (2): 363–376. doi:10.4202/app.00161.2015.
- ^ Zhang, J.; Wagh, P.; Guay, D.; Sanchez-Pulido, L.; Padhi, B. K.; Korzh, V.; Andrade-Navarro, M. A.; Akimenko, M. A. (2010). "Loss of fish actinotrichia proteins and the fin-to-limb transition". Nature. 466 (7303): 234–237. Bibcode:2010Natur.466..234Z. doi:10.1038/nature09137. PMID 20574421. S2CID 205221027.
- ^ 햄릿 1999, 페이지 528.
- ^ 상어에서 이성질체 꼬리의 기능: 꾸준한 수평 수영과 수직 이동 동안의 정량적 웨이크 역학 - 실험 생물학 저널 205, 2365–2374 (2002)
- ^ "A Shark's Skeleton & Organs". Archived from the original on 5 August 2010. Retrieved 14 August 2009.
- ^ Burrow, Carole (2021). Acanthodii, Stem Chondrichthyes. Verlag Dr. Friedrich Pfeil. ISBN 978-3-89937-271-7. OCLC 1335983356.
- ^ Andreev, Plamen S.; Sansom, Ivan J.; Li, Qiang; Zhao, Wenjin; Wang, Jianhua; Wang, Chun-Chieh; Peng, Lijian; Jia, Liantao; Qiao, Tuo; Zhu, Min (September 2022). "Spiny chondrichthyan from the lower Silurian of South China". Nature. 609 (7929): 969–974. Bibcode:2022Natur.609..969A. doi:10.1038/s41586-022-05233-8. ISSN 1476-4687. PMID 36171377. S2CID 252570103.
- ^ Michael, Bright. "Jaws: The Natural History of Sharks". Columbia University. Archived from the original on 24 December 2011. Retrieved 29 August 2009.
- ^ Grogan, Eileen D.; Lund, Richard (2008). "A basal elasmobranch, Thrinacoselache gracia n. gen and sp., (Thrinacodontidae, new family) from the Bear Gulch Limestone, Serpukhovian of Montana, USA". Journal of Vertebrate Paleontology. 28 (4): 970–988. Bibcode:2008JVPal..28..970G. doi:10.1671/0272-4634-28.4.970. S2CID 84735866.
- ^ Frey, Linda; Coates, Michael; Ginter, Michał; Hairapetian, Vachik; Rücklin, Martin; Jerjen, Iwan; Klug, Christian (2019). "The early elasmobranch Phoebodus: phylogenetic relationships, ecomorphology and a new time-scale for shark evolution". Proceedings of the Royal Society B: Biological Sciences. 286 (1912). doi:10.1098/rspb.2019.1336. PMC 6790773. PMID 31575362. S2CID 203619135.
- ^ 상어 피닝.국제휴먼 소사이어티.
- ^ Vannuccini S (1999). "Shark utilization, marketing and trade". FAO Fisheries Technical Paper. 389.
- ^ "In China, victory for wildlife conservation as citizens persuaded to give up shark fin soup - The Washington Post". www.washingtonpost.com. Retrieved 20 January 2017.
- ^ a b c Sfakiotakis, M; Lane, DM; Davies, JBC (1999). "Review of Fish Swimming Modes for Aquatic Locomotion" (PDF). IEEE Journal of Oceanic Engineering. 24 (2): 237–252. Bibcode:1999IJOE...24..237S. CiteSeerX 10.1.1.459.8614. doi:10.1109/48.757275. S2CID 17226211. Archived from the original (PDF) on 24 December 2013.
- ^ Franc, Jean-Pierre and Michel, Jean-Marie (2004) Cavitation Springer의 기본.ISBN 9781402022326.
- ^ a b Brahic, Catherine (28 March 2008). "Dolphins swim so fast it hurts". New Scientist. Retrieved 31 March 2008.
- ^ Nauen, JC; Lauder, GV (2001a). "Locomotion in scombrid fishes: visualization of flow around the caudal peduncle and finlets of the Chub mackerel Scomber japonicus". Journal of Experimental Biology. 204 (Pt 13): 2251–63. doi:10.1242/jeb.204.13.2251. PMID 11507109.
- ^ Nauen, JC; Lauder, GV (2001b). "Three-dimensional analysis of finlet kinematics in the Chub mackerel (Scomber japonicus)". The Biological Bulletin. 200 (1): 9–19. doi:10.2307/1543081. JSTOR 1543081. PMID 11249216. S2CID 28910289.
- ^ Nauen, JC; Lauder, GV (2000). "Locomotion in scombrid fishes: morphology and kinematics of the finlets of the Chub mackerel Scomber japonicus" (PDF). Journal of Experimental Biology. 203 (Pt 15): 2247–59. doi:10.1242/jeb.203.15.2247. PMID 10887065.
- ^ Flammang, BE; Lauder, GV; Troolin, DR; Strand, TE (2011). "Volumetric imaging of fish locomotion". Biology Letters. 7 (5): 695–698. doi:10.1098/rsbl.2011.0282. PMC 3169073. PMID 21508026.
- ^ Fish, FE; Lauder, GV (2006). "Passive and active flow control by swimming fishes and mammals". Annual Review of Fluid Mechanics. 38 (1): 193–224. Bibcode:2006AnRFM..38..193F. doi:10.1146/annurev.fluid.38.050304.092201. S2CID 4983205.
- ^ Magnuson JJ (1978) "스콤브리드 피쉬에 의한 로코모션: 수역학, 형태와 행동" 어류 생리학, 제7권: 로코모션, WS Hoar and DJ Randall (Eds) 학술 출판부240-308페이지ISBN 9780123504074.
- ^ 해상에서의 선박의 움직임 2011년 11월 25일 웨이백 머신에서 2012년 11월 22일 회수.
- ^ Rana and Joag (2001) Classical Mechanics 391페이지, Tata McGraw-Hill EducationISBN 9780074603154.
- ^ a b c Alevizon WS (1994) "캐리비안 리프 생태를 위한 물고기 가이드" 걸프 출판사 ISBN 1-55992-077-7
- ^ Lingham-Soliar, T. (2005). "Dorsal fin in the white shark,Carcharodon carcharias: A dynamic stabilizer for fast swimming". Journal of Morphology. 263 (1): 1–11. doi:10.1002/jmor.10207. PMID 15536651. S2CID 827610.
- ^ a b 어류학 플로리다 자연사 박물관.2012년 11월 22일 회수.
- ^ Masterson, J. "Gambusia affinis". Smithsonian Institution. Retrieved 21 October 2011.
- ^ Kuntz, Albert (1913). "Notes on the Habits, Morphology of the Reproductive Organs, and Embryology of the Viviparous Fish Gambusia affinis". Bulletin of the United States Bureau of Fisheries. 33: 181–190.
- ^ Kapoor BG and Khanna B (2004) 어류학 핸드북 pp. 497–498, Springer Science & Business Media.ISBN 9783540428541.
- ^ Helfman G, Collette BB, Facey DH and Bowen BW (2009) 물고기의 다양성: 생물학, 진화, 생태학 p. 35, Wiley-Blackwell.ISBN 978-1-4051-2494-2
- ^ Maxwell; et al. (2018). "Re‐evaluation of the ontogeny and reproductive biology of the Triassic fish Saurichthys (Actinopterygii, Saurichthyidae)". Palaeontology. 61: 559–574. doi:10.5061/dryad.vc8h5.
- ^ "System glossary". FishBase. Retrieved 15 February 2013.
- ^ Heinicke, Matthew P.; Naylor, Gavin J. P.; Hedges, S. Blair (2009). The Timetree of Life: Cartilaginous Fishes (Chondrichthyes). Oxford University Press. p. 320. ISBN 978-0191560156.
- ^ a b 세계의 수생생활 pp. 332-333, Marshall Cavendish Corporation, 2000ISBN 9780761471707.
- ^ Dement J 종의 스포트라이트: 애틀랜틱 세일피시 (Istiopphorus albicans) 2010년 12월 17일 웨이백 머신 littoralsociety.org 에서 보관.2012년 4월 1일 회수.
- ^ Bertelsen E and Pietsch TW (1998). Encyclopedia of Fishes. San Diego: Academic Press. pp. 138–139. ISBN 978-0-12-547665-2.
- ^ 퍼플 플라잉 거나드, 닥틸롭테나 오리엔탈리스 (Cubier, 1829) 호주 박물관업데이트: 2012년 9월 15일.검색됨: 2012년 11월 2일.
- ^ Froese, Rainer; Pauly, Daniel (eds.) (2012).피시베이스의 "닥틸롭테나 오리엔탈리스".2012년 11월 버전.
- ^ 암컷 물고기들이 짝을 유혹하기 위해 지느러미를 뽐내고 있습니다. 2010년 10월 8일.
- ^ Baldauf, SA; Bakker, TCM; Herder, F; Kullmann, H; Thünken, T (2010). "Male mate choice scales female ornament allometry in a cichlid fish". BMC Evolutionary Biology. 10: 301. doi:10.1186/1471-2148-10-301. PMC 2958921. PMID 20932273.
- ^ 슐츠, 켄 (2011) 켄 슐츠의 소금물 물고기 현장 가이드 250페이지, 존 와일리 & 선즈ISBN 978111803985.
- ^ 굿리치, 에드윈 S. 1906."메모리즈:물고기의 중앙 지느러미와 쌍지느러미의 발달, 구조 및 기원에 관한 노트." 세포 과학 저널 s2-50 (198): 333–76
- ^ a b Brand, Richard A (2008). "Origin and Comparative Anatomy of the Pectoral Limb". Clinical Orthopaedics and Related Research. 466 (3): 531–42. doi:10.1007/s11999-007-0102-6. PMC 2505211. PMID 18264841.
- ^ a b c Coates, M. I. (2003). "The Evolution of Paired Fins". Theory in Biosciences. 122 (2–3): 266–87. doi:10.1078/1431-7613-00087.
- ^ 게겐바우어, C., F.J. 벨, 그리고 E.레이 랭커스터 1878년비교 해부학의 요소. 칼 게겐바우어 작... F에 의한. 제프리 벨... E가 쓴 번역서와 서문. 레이 랭커스터...런던,: 맥밀런 앤 컴퍼니,
- ^ 굿리치, 에드윈 S. 1906."메모리즈:물고기의 중앙 지느러미와 쌍지느러미의 발달, 구조 및 기원에 관한 노트." 세포 과학 저널 s2-50 (198): 333–76
- ^ a b Begemann, Gerrit (2009). "Evolutionary Developmental Biology". Zebrafish. 6 (3): 303–4. doi:10.1089/zeb.2009.0593.
- ^ Cole, Nicholas J.; Currie, Peter D. (2007). "Insights from Sharks: Evolutionary and Developmental Models of Fin Development". Developmental Dynamics. 236 (9): 2421–31. doi:10.1002/dvdy.21268. PMID 17676641. S2CID 40763215.
- ^ Freitas, Renata; Zhang, GuangJun; Cohn, Martin J. (2006). "Evidence That Mechanisms of Fin Development Evolved in the Midline of Early Vertebrates". Nature. 442 (7106): 1033–37. Bibcode:2006Natur.442.1033F. doi:10.1038/nature04984. PMID 16878142. S2CID 4322878.
- ^ Gillis, J. A.; Dahn, R. D.; Shubin, N. H. (2009). "Shared Developmental Mechanisms Pattern the Vertebrate Gill Arch and Paired Fin Skeletons". Proceedings of the National Academy of Sciences. 106 (14): 5720–24. Bibcode:2009PNAS..106.5720G. doi:10.1073/pnas.0810959106. PMC 2667079. PMID 19321424.
- ^ "원시 물고기는 초보적인 손가락을 가지고 있었다" 사이언스 데일리, 2008년 9월 23일.
- ^ Laurin, M. (2010). How Vertebrates Left the Water. Berkeley, California, USA.: University of California Press. ISBN 978-0-520-26647-6.
- ^ Hall, Brian K (2007) 사지에 핀인: 시카고 대학의 진화, 개발, 변혁 대학 출판부.ISBN 9780226313375.
- ^ 슈빈, 닐 (2009) 당신의 내면의 물고기: 35억 인류의 역사를 향한 여행 빈티지 북스ISBN 9780307277459.UCTV 인터뷰
- ^ Clack, Jennifer A (2012) "지느러미에서 발까지" 6장 187-260페이지: Gaining Ground, 제2판: 테트라포드의 기원과 진화, 인디애나 대학 출판부ISBN 9780253356758.
- ^ Moore, John A (1988). "[www.sicb.org/dl/saawok/449.pdf "Understanding nature—form and function"] Page 485". American Zoologist. 28 (2): 449–584. doi:10.1093/icb/28.2.449.
- ^ 룽피쉬는 육지에서의 삶에 대한 통찰력을 제공합니다: '인간은 단지 변형된 물고기일 뿐입니다' 사이언스 데일리, 2011년 10월 7일.
- ^ Cole, NJ; Hall, TE; Don, EK; Berger, S; Boisvert, CA; et al. (2011). "Development and Evolution of the Muscles of the Pelvic Fin". PLOS Biology. 9 (10): e1001168. doi:10.1371/journal.pbio.1001168. PMC 3186808. PMID 21990962.
- ^ 폐어를 위한 작은 발걸음, 걷는 것의 진화를 위한 큰 걸음" 사이언스 데일리, 2011년 12월 13일.
- ^ King, HM; Shubin, NH; Coates, MI; Hale, ME (2011). "Behavioral evidence for the evolution of walking and bounding before terrestriality in sarcopterygian fishes". Proceedings of the National Academy of Sciences. 108 (52): 21146–21151. Bibcode:2011PNAS..10821146K. doi:10.1073/pnas.1118669109. PMC 3248479. PMID 22160688.
- ^ Shubin, N; Tabin, C; Carroll, S (1997). "Fossils, genes and the evolution of animal limbs" (PDF). Nature. 388 (6643): 639–648. Bibcode:1997Natur.388..639S. doi:10.1038/41710. PMID 9262397. S2CID 2913898. Archived from the original (PDF) on 16 September 2012.
- ^ 척추동물 비행: 캘리포니아 대학의 세 가지 솔루션입니다.2005년 9월 29일 업데이트.
- ^ "Scientists find missing link between the dolphin, whale and its closest relative, the hippo". Science News Daily. 25 January 2005. Archived from the original on 4 March 2007. Retrieved 18 June 2007.
- ^ Gatesy, J. (1 May 1997). "More DNA support for a Cetacea/Hippopotamidae clade: the blood-clotting protein gene gamma-fibrinogen". Molecular Biology and Evolution. 14 (5): 537–543. doi:10.1093/oxfordjournals.molbev.a025790. PMID 9159931.
- ^ Flynn JJ, Finarelli JA, Zehr S, Hsu J, Nedbal MA (2005). "Molecular phylogeny of the carnivora (mammalia): assessing the impact of increased sampling on resolving enigmatic relationships". Systematic Biology. 54 (2): 317–337. doi:10.1080/10635150590923326. PMID 16012099.
- ^ Felts WJL "고래과 물갈퀴의 일부 기능적 및 구조적 특징" 255-275쪽: Norris KS (ed.) 고래, 돌고래, 그리고 돌고래, 그리고 돌고래, 그리고 돌고래, 캘리포니아 대학 출판부.
- ^ 고래의 진화 캘리포니아 대학 박물관2012년 11월 27일 회수.
- ^ Thewissen, JGM; Cooper, LN; George, JC; Bajpai, S (2009). "From Land to Water: the Origin of Whales, Dolphins, and Porpoises" (PDF). Evo Edu Outreach. 2 (2): 272–288. doi:10.1007/s12052-009-0135-2. S2CID 11583496.
- ^ Martill D.M. (1993)."수프 기판:독일의 포시도니아 셰일(Lower Jurassic)의 이크티오사우루스(Ichthyosaurus)의 예외적인 보존을 위한 매개체".카우피아 - Darmstädter Beiträgezur Naturgeschichte, 2:77-97
- ^ 굴드, 스티븐 제이(1993)는 여덟 마리의 아기 돼지에서 "모양이 안 좋아졌다"고 말했습니다. 자연사에 대한 성찰.노튼, 179-94ISBN 9780393311396.
- ^ "Charlie: CIA's Robotic Fish — Central Intelligence Agency". www.cia.gov. Archived from the original on 16 August 2013. Retrieved 12 December 2016.
- ^ Richard Mason. "What is the market for robot fish?". Archived from the original on 4 July 2009.
- ^ Witoon Juwarahawong. "Fish Robot". Institute of Field Robotics. Archived from the original on 4 November 2007. Retrieved 25 October 2007.
- ^ "Robotic fish powered by Gumstix PC and PIC". Human Centred Robotics Group at Essex University. Retrieved 25 October 2007.
- ^ "Robotic fish make aquarium debut". cnn.com. CNN. 10 October 2005. Retrieved 12 June 2011.
- ^ Walsh, Dominic (3 May 2008). "Merlin Entertainments tops up list of London attractions with aquarium buy". thetimes.co.uk. Times of London. Retrieved 12 June 2011.
- ^ Festo의 경우, 2009년 5월 18일, 자연은 Way Control Engineering을 보여줍니다.
- ^ 바이오닉 펭귄들은 물속을 날아다닙니다. 2009년 4월 27일, 에어 기즈매그.
- ^ 페스토 아쿠아레이 로봇 기술, 2009년 4월 20일.
- ^ 2012년 7월 12일 페스토 엔지니어링 TV의 아쿠아 젤리 로봇 젤리피쉬.
- ^ 경량 로봇: 페스토의 플라잉 서커스 2011년 7월 18일 웨이백 머신 더 엔지니어에서 2015년 9월 19일 보관.
- ^ Huge Herr, D. Robert G (October 2004). "A Swimming Robot Actuated by Living Muscle Tissue". Journal of NeuroEngineering and Rehabilitation. 1 (1): 6. doi:10.1186/1743-0003-1-6. PMC 544953. PMID 15679914.
- ^ 바이오메카트로닉스 작동 방법/2012년 11월 22일 검색.
- ^ Lauder, G. V. (2011). "Swimming hydrodynamics: ten questions and the technical approaches needed to resolve them" (PDF). Experiments in Fluids. 51 (1): 23–35. Bibcode:2011ExFl...51...23L. doi:10.1007/s00348-009-0765-8. S2CID 890431.
서지학
- Hamlett, William C. (1999). Sharks, skates, and rays: the biology of elasmobranch fishes (1st ed.). The Johns Hopkins University Press. p. 56. ISBN 978-0-8018-6048-5.
추가열람
- Hall, Brian K (2007) 사지에 핀인: 시카고 대학의 진화, 개발, 변혁 대학 출판부.ISBN 9780226313375.
- 헬프만 G, 콜렛 BB, 페이시 DE 및 보웬 BW(2009) "이동과 급전의 기능 형태학" 8장 101-116쪽.인: 물고기의 다양성: 생물학, 존 와일리 & 선즈ISBN 9781444311907.
- Lauder, GV; Nauen, JC; Drucker, EG (2002). "Experimental Hydrodynamics and Evolution: Function of Median Fins in Ray-finned Fishes". Integr. Comp. Biol. 42 (5): 1009–1017. doi:10.1093/icb/42.5.1009. PMID 21680382.
- Lauder, GV; Drucker, EG (2004). "Morphology and experimental hydrodynamics of fish fin control surfaces" (PDF). Journal of Oceanic Engineering. 29 (3): 556–571. Bibcode:2004IJOE...29..556L. doi:10.1109/joe.2004.833219. S2CID 36207755.
외부 링크
- 골편어류의 핀피도트리키아 상동성
- 물고기의 핀 어스라이프 거미줄
- 로봇 물고기가 오염을 발견할 수 있을까요?스탭의 작동 방식.2012년 1월 30일 접속.


