물고기 턱
Fish jaw
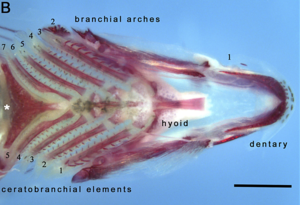
대부분의 뼈 있는 물고기는 주로 뼈로 만들어진 두 개의 턱을 가지고 있다. 1차 구강턱은 입을 벌리고 닫으며, 두 번째 인두턱 세트가 목 뒤쪽에 위치한다. 구강턱은 먹이를 물고 으스러뜨려 포획하고 조종하는 데 쓰인다. 인두 안쪽에 위치하기 때문에 소위 인두 턱은 음식을 더욱 가공하여 입에서 위장으로 옮기는 데 쓰인다.[2][3]
상어나 광선과 같은 수족구 물고기는 주로 연골로 만들어진 구강 턱 한 세트를 가지고 있다. 그들은 인두 턱을 가지고 있지 않다. 일반적으로 턱은 관절형이며, 위턱과 아래턱으로 구성되어 수직으로 반대하며, 순서가 정해진 수많은 치아를 가질 수 있다. 갑상선 어류는 여러 세트(폴리피오돈트)를 성장시키고 컨베이어 벨트 방식으로 새 치아를 안쪽 턱 표면에서 횡방향으로 움직여서 착용할 때 치아를 교체한다. 대부분의 뼈다귀 어류에서도 치아를 여러 번 교체하지만, 수족구와는 달리 새 치아는 오래된 치아가 빠진 후에야 분출한다.
죠스는 아마도 턱이 없는 물고기의 아가미를 지탱하는 인두 아치에서 유래되었을 것이다. 가장 초기 턱은 약 4억 3천만년 전 실루리아 해 동안 멸종된 플라코다름과 가시가 있는 상어에게 나타났다. 턱이 제공하는 원래 선택적 이점은 아마도 먹이를 주는 것이 아니라 호흡 효율의 증가와 관련이 있을 것이다. 턱은 아가미를 가로질러 물을 펌프질하기 위해 부칼 펌프에서 사용되었다. 먹이를 주기 위한 턱의 익숙한 사용은 많은 척추동물에서 주요한 기능이 되기 전에 2차적인 기능으로 발전했을 것이다. 인간의 턱을 포함한 모든 척추동물 턱은 초기 물고기 턱으로부터 진화했다. 초기 척추동물 턱의 외형은 "아마도 척추동물 역사상 가장 심오하고 급진적인 진화 단계"로 묘사되어 왔다.[4][5] 턱이 없는 물고기는 턱이 없는 물고기보다 생존에 어려움이 많았고 턱이 없는 물고기는 대부분 멸종했다.
죠스는 연결 메커니즘을 사용한다. 이러한 연결고리는 많은 전문화된 먹이 메커니즘을 진화시켜온 쇄골과 같은 뼈 있는 물고기의 머리에서 특히 흔하고 복잡할 수 있다. 특히 턱 돌출의 연계 메커니즘이 발달했다. 석션 공급의 경우 연결된 4바 연결 시스템이 입의 조정된 개방과 부칼공동의 3차원 확장을 담당한다. 또한 4봉 연결은 전축의 돌출에 대한 책임이 있으며,[6] 3개의 주요 4봉 연결 시스템이 어류의 부칼 공동의 횡방향과 전방의 확장을 일반적으로 기술하게 된다.[6][7] 동물들의 서로 다른 종류의 연결에 대한 가장 철저한 개요는 M에 의해 제공되었다. 특히 생물학적 시스템에 잘 맞는 새로운 분류 시스템을 설계하기도 한 뮬러.[8]
스컬

물고기의 두개골은 느슨하게 연결된 일련의 뼈로 형성된다. 램프리스와 상어는 위턱과 아래턱이 모두 분리된 원소인 카르딜라긴성 내분비만을 가지고 있다. 뼈다귀 물고기는 폐어나 보금자리에서 다소 일관성이 있는 두개골 지붕을 형성하는 추가적인 피부 뼈를 가지고 있다.
보다 단순한 구조는 턱이 없는 물고기에서 발견되는데, 두개골은 뇌를 부분적으로만 감싸고 있는 장막 원소의 수조 같은 바구니로 표현되며, 내이 및 단 하나의 콧구멍을 위한 캡슐과 관련된다.[9]
상어와 같은 수족관 물고기들도 단순한 두개골을 가지고 있다. 두개골은 뇌 주위에 케이스를 형성하는 단일 구조로, 아래쪽 표면과 옆면을 감싸고 있지만, 큰 폰타넬로서 윗부분은 항상 부분적으로 열려 있다. 두개골의 가장 앞쪽에는 연골의 앞판, 후각을 감싸는 로스트룸, 캡슐 등이 있다. 이들 뒤에는 궤도를 그리며, 이어 내이 구조를 감싸고 있는 캡슐 한 쌍이 추가로 있다. 마지막으로, 후방을 향한 두개골 테이퍼, 포아멘 마그넘이 첫 번째 척추와 관절을 이루며 하나의 콘딜 바로 위에 놓여 있다. 게다가, 두개골 전체에 걸쳐 다양한 지점들에는 두개골 신경들을 위한 더 작은 포아미나가 있다. 턱은 거의 항상 적절한 두개골과는 구별되는 별도의 연골 후프들로 구성된다.[9]
광어류에서는 원시적 패턴에서 상당한 수정이 있었다. 두개골의 지붕은 일반적으로 잘 형성되어 있으며, 테트라포드(tetrapods)의 뼈와 정확한 관계가 불분명하지만, 대개는 편의를 위해 비슷한 이름을 붙인다. 그러나 두개골의 다른 요소들은 축소될 수 있다; 확대된 궤도의 뒤에는 광대 부분이 거의 없고, 그 사이에 뼈가 있다면 거의 없다. 위턱은 주로 프리맥실라에서 형성되는 경우가 많으며, 맥실라 자체는 더 뒤쪽에 위치하며, 추가적인 뼈인 동정골은 턱과 나머지 두개골을 연결한다.[9]
비록 화석화된 로브핀 물고기의 두개골은 초기 4엽 물고기의 두개골과 비슷하지만, 살아있는 폐고기의 두개골도 마찬가지라고는 말할 수 없다. 두개골 지붕은 완전히 형성되지 않았으며, 테트라포드의 뼈와 직접적인 관계가 없는, 다소 불규칙하게 생긴 여러 개의 뼈로 구성되어 있다. 위턱은 피테리고이드와 구토만으로 형성되는데, 모두 이를 품고 있다. 두개골의 대부분은 연골에서 형성되며 전체적인 구조는 축소된다.[9]
구강턱
더 낮게

척추동물의 경우 아래턱(필수 또는 턱뼈)[10]은 두개골과 두개골을 이루는 뼈다. 로브핀 물고기와 초기 화석 테트라포드에서는 포유류의 하악골에 해당하는 뼈는 아래턱에 있는 몇 개의 뼈 중 단지 가장 크다. 의치골이라고 하며, 턱의 바깥 표면의 몸체를 형성한다. 아래에는 여러 개의 비장골과 접하고 있으며, 턱의 각도는 바로 위 각골과 직사각형 뼈로 이루어져 있다. 턱의 안쪽 표면은 전골에 의해 정렬되어 있고, 관절뼈는 두개골과 함께 관절 모양을 형성한다. 마침내 세 개의 좁은 코로노이드 뼈 세 개가 전골 위에 놓여 있다. 이름에서 알 수 있듯이 치아의 대다수는 의치에 붙어있지만, 흔히 코로노이드 뼈에도 치아가 있으며, 때로는 전치골에도 치아가 있다.[11]
그러나 이 복잡한 원시 패턴은 뼈가 융합되거나 완전히 사라졌기 때문에 척추동물의 대부분에서 여러 가지 정도로 단순화되었다. 텔레오스트에서는 틀니, 관절, 각진 뼈만 남는다.[11] 상어와 같은 카르딜라겐성 물고기는 다른 척추동물의 아래턱에서 발견된 뼈 중 어떤 것도 가지고 있지 않다. 대신, 그들의 아래턱은 다른 그룹의 메켈의 연골과 동음이의인 카르딜라겐 구조로 구성되어 있다. 이것은 철갑상어와 같은 몇몇 원시적인 뼈의 물고기에서도 턱의 중요한 요소로 남아 있다.[11]
상부
위턱, 즉 맥실라[12][13](maxilla)는 위턱을 형성하는 구개열을 따라 두 개의 뼈가 융합된 것이다. 이것은 맨디블(하악 턱)과 비슷한데, 이 역시 맨디블 심포석에서 두 반쪽이 융합된 것이다. 뼈 있는 물고기에서 맥시야는 "상피맥시야"라고 불리며, 하악맥시야는 "하악맥시야"이다. 맥실라의 치경공정은 윗니를 잡고 있으며, 이를 맥실아치라고 한다. 대부분의 척추동물에서, 위턱의 가장 앞부분인 포유류에서 근치들이 붙어있는 부분은 별도의 한 쌍의 뼈인 전축류로 구성되어 있다. 뼈만 앙상한 생선에서는 맥실라와 프리맥실라 모두 비교적 판처럼 생긴 뼈로, 위턱의 옆구리와 얼굴 일부만을 이루고 있으며 프리맥실라 역시 콧구멍의 아래쪽 경계를 이루고 있다.[14] 상어나 광선과 같은 수족구류도 진정한 최대치가 부족하다. 그들의 위턱은 대신 다른 척추동물에서 발견된 뼈와 동질적이지 않은 수족관에서 형성된다.[14]
어떤 물고기는 로스트럼이라고 불리는 위턱뼈가 영구적으로 돌출되어 있다. 빌피쉬(마린, 황새치, 돛새치)는 로스트럼(빌)을 사용해 먹이를 베고 기절시킨다. 패들피쉬, 도깨비 상어, 망치머리 상어는 약한 전기장을 감지하여 먹이의 존재를 알리는 전기 감응기로 가득 차 있다. 톱샤크와 멸종위기에 처한 톱새우에는 전기 감응성이 뛰어나고 슬래싱에 사용되는 로스트럼(톱)이 있다.[15] 로스트룸은 물고기 앞에서 통풍으로 뻗어 있다. 해머헤드의 경우 로스트룸(망치)은 통풍과 측면(측면)으로 모두 확장된다.
- 로스트럼이 있는 생선(상악뼈 확장)

톱피쉬는 전기에 민감한 로스트럼(톱)을 가지고 있는데, 먹이를 베는 데도 사용된다.



턱 돌출
텔레오스트에는 움직일 수 있는 프리맥시야(상턱 끝에 있는 뼈)와 이에 상응하는 턱 근육의 변화가 있어 입 밖으로 턱이 튀어나올 수 있다. 이것은 그들이 먹이를 잡아 입으로 끌어당길 수 있는 큰 장점이 있다. 보다 파생된 텔레오스트에서는 확대된 프리맥시야가 주 이빨을 가진 뼈로, 아래턱에 붙어 있는 맥시야가 지렛대 역할을 하여 입이 열리고 닫힐 때 프리맥시라를 밀고 당긴다. 이 돌기 있는 턱은 적어도 다섯 번은 독립적으로 진화한 텔레오스트의 진화적인 신기한 것이다.[16]
프리맥시야는 신경낭(뇌통)에 붙어 있지 않으며, 입을 내밀어 원형 개구부를 만드는 역할을 한다. 이것은 입 안의 압력을 낮추어 먹잇감을 안으로 빨아들인다. 이어 아래턱과 맥실라(턱의 주요 상부 고정골)를 뒤로 당겨 입을 닫게 하고, 물고기가 먹이를 잡을 수 있게 된다. 이와는 대조적으로 턱을 닫는 것만으로도 음식을 입에서 밀어낼 위험이 있다. 보다 진보된 텔레오스트에서는 프리맥시야가 확대되어 이빨이 있는 반면, 맥시야는 이빨이 없다. 맥실라는 프리맥실라와 아래턱을 모두 앞으로 밀어주는 기능을 한다. 입을 열기 위해 인덕터 근육이 맥실라 윗부분을 뒤로 당겨 아래턱을 앞으로 밀어낸다. 또 맥실라는 약간 회전하는데, 이것은 프리맥실라와 연동되는 뼈의 과정을 앞으로 밀어낸다.[17]
텔레osts는 두개골 내의 인대 연결과 관련된 네 가지 다른 메커니즘 중 하나를 사용하여 이 턱 돌출에 도달한다.[18]


- 하악성 우울증 메커니즘: 아래턱의 우울증(필수)은 위턱에 연결된 인대와 힘줄을 통한 힘 전달을 통해 프리막시야를 돌출부로 당기거나 밀어 넣는다(예: 시프린우스, 라브루스).[18] 이것은 가장 일반적으로 사용되는 메커니즘이다.
- 트위스트 맥실라 메커니즘: 하악골의 수축은 맥실라가 세로축을 중심으로 비틀어 전축(예: 무길)이 돌출되는 원인이 된다.[18]
- 분리 메커니즘: 프리맥실라의 돌출은 신경크라늄의 고도를 통해 이루어지며 프리맥실라가 전방으로 이동하게 된다. 신경크라늄의 움직임은 위턱의 운동학(예: Spathodus erythrodon)과 결합되지 [18][19]않아 먹이 포획과 조작 중에 턱의 다용성과 모듈성을 더 높일 수 있다.
- 서스펜션 납치 메커니즘: 현수막의 측면 팽창(팔라틴, 프테리고이드 시리즈, 사방골의 조합)은 전축이 앞쪽으로 튀어나오게 하는 인대를 잡아당긴다(예:[18][19] 페트로틸라피아 삼지디거).
일부 텔레ost는 이러한 메커니즘 중 하나 이상을 사용한다(예: 페트로틸라피아).[18]
쇄골은 턱 구조 때문에 물고기에게 먹이를 주는 생물역학에서 주요 연구 종으로 자리잡았다. 그들은 긴 입을 가지고 있고, 보통 바깥으로 튀어나온 별도의 턱 이빨을 가지고 있다.[20] 많은 종들이 두꺼운 입술로 쉽게 알아볼 수 있는데, 그 속은 가끔 신기하게 접혀 있는데, 이것은 "립피쉬"(Lip-fisesche)라는 독일식 이름을 갖게 한 특이점이다.[21]
코뼈와 하악골은 후두부에서 경질신경크라니움으로 연결되며, 맥실라의 상하관절은 각각 이 두 뼈의 앞끝과 연결되어 움직이는 관절에 의해 연결된 4개의 경골의 고리가 형성된다. 이 "4-bar 연결"은 주어진 기계적 결과(빠른 턱 돌출 또는 강력한 물림)를 달성하기 위해 수많은 배치를 허용함으로써 형태학을 기능에서 분리하는 특성을 가지고 있다. 쇄골의 실제 형태학은 이를 반영하는데, 많은 선들이 비슷하거나 동일한 생태적 틈새에서 동일한 기능적 출력을 내는 다른 턱 형태학을 보여준다.[20]
물고기에서 발견되는 가장 극단적인 턱 돌출은 슬링조(slingjaw wrasse), 에피불루스(Epibulus insidiator)에서 발생한다. 이 물고기는 턱을 머리 길이의 65%까지 뻗을 수 있다.[22] 이 종은 그것의 빠르고 극단적인 턱 돌출부를 이용하여 작은 물고기와 갑각류를 잡아낸다. 이 종에 속하는 속은 독특한 인대 1개(보메로-간체)와 두 개의 확대된 인대(간체외-만두근, 프리맥실라-맥실라)를 가지고 있는데, 두개골의 형태에 약간의 변화와 함께 극도의 턱 돌출이 가능하다.
인두턱

위턱은 1차(구강) 턱과 구별되는 2차 턱 세트다. 그것들은 대부분의 뼈 있는 물고기의 목, 즉 인두 속에 들어 있다. 그것들은 구강 턱과 비슷한 방식으로, 더 이상 호흡 기능이 없는 5번째 길 아치를 변형시키는 것으로서 유래된 것으로 여겨진다. 처음 네 개의 아치는 여전히 아가미로 기능한다. 인두턱은 구강턱과 달리 턱관절이 없고 대신 근육의 슬링으로 받쳐진다.
_Schlundzaehne_1x_Fig_147_(Matschie_et_al._1909).svg)
눈에 띄는 예는 모레이 뱀장어에게서 나타난다. 대부분의 물고기의 위턱은 이동성이 없다. 병아리의 위턱은 이동성이 매우 높으며, 아마도 그들이 서식하는 굴의 수축된 성질에 적응한 것으로서 다른 물고기들이 입에 음압을 만들어 삼키는 것처럼 삼키는 능력을 억제한다. 대신, 이 병아리가 먹이를 물면, 먼저 구강턱으로 정상적으로 물어서 먹이를 잡는다. 그 직후, 인두턱은 앞으로 나와 먹이를 물어 잡아 쥐게 하고, 다시 수축하여, 먹이를 모레이 뱀장어의 굴레 아래로 끌어내려 삼켜지게 한다.[23]
모든 척추동물은 인두를 가지고 있는데, 이는 먹이를 주는 것과 호흡하는 데 모두 사용된다. 인두는 머리 측면에 인두 아치라고 불리는 여섯 개 이상의 아웃포켓팅을 통해 발달하는 동안 발생한다. 인두 아치는 척추동물마다 다른 방식으로 골격, 근육, 순환계에 많은 다른 구조를 만들어 낸다. 인두 아치는 화음을 통해 염기성 중수체에게까지 거슬러 올라간다. 그들은 또한 인두성 기구의 내피 소매치기를 공유한다. 유사한 유전자 발현 패턴은 암피옥소와 헤미코드의 발달하는 인두에서 발견될 수 있다. 그러나 척추동물의 인두는 신경 파고 세포의 기여를 통해 내골격계 지지를 발생시킨다는 점에서 특이하다.[24]
수족관
수족구(상어, 광선, 스케이트)는 수족구 턱을 가지고 있다. 턱 표면은 (척추와 아가미 아치에 비해) 신체적 스트레스에 많이 노출되어 있어 추가적인 힘이 필요하다. 그것은 "테세래"라고 불리는 작은 육각형의 판을 층층이 있는데, 이것은 모자이크로 배열된 칼슘 염의 결정 블록이다.[25] 이것은 다른 동물에서 발견되는 뼈 조직에서 발견되는 것과 거의 같은 힘을 이 지역들에게 준다.
일반적으로 상어는 테세레 한 층만 가지고 있지만, 황소상어, 호랑이상어, 백상어 등 큰 표본의 턱은 몸 크기에 따라 2~3층 이상이다. 큰 백상어의 턱에는 최대 다섯 개의 층이 있을 수 있다.[26] 로스트럼(snout)에서 연골은 스펀지가 많고 유연해 충격의 힘을 흡수할 수 있다.
상어와 다른 현존하는 엘라스모브란치에서는 위턱이 두개골에 융접되지 않고 아래턱은 위턱과 관절한다. 연조직과 이러한 요소들을 연결하는 추가 관절의 배열을 전체적으로 턱 서스펜션이라고 한다. 원형 턱걸이가 몇 가지 있다: 원형 턱걸이가 있고, 원형 턱걸이가 있고, 궤도가형 턱걸이가 있다. 원형극에서, 팔라토쿼드레이트에는 주로 인대가 그것을 전방으로 중단시키는 콘드로크라늄과 함께 포스트보리탈 관절이 있다. 하이드로이드는 하악골과 후방으로 관절하지만, 위턱과 아래턱에 거의 지탱하지 못하는 것으로 보인다. 궤도 중심에서 궤도 공정이 궤도벽과 경첩되며, 효이드는 대부분의 현수 지지대를 제공한다. 이와는 대조적으로, 위턱과 두개골 사이에 에트모이드 관절이 있는 반면, 앞쪽 인대에 비해 턱 지지대가 훨씬 더 높을 가능성이 높다. 마지막으로, 진정한 요오스티야로도 알려진, 의협에서, 하악골 카르딜로는 두개골에 대한 인대 연결이 부족하다. 대신, 요만두 카르딜라지는 턱 지지의 유일한 수단을 제공하는 반면, 세라토히알과 기저부 원소는 아래턱과 관절을 이루지만 나머지 효이드와는 분리된다.[27][28][29]
이빨

.jpeg)
죠스는 대부분의 물고기에 간단한 뾰족한 이빨을 위한 플랫폼을 제공한다. 폐피쉬와 치마는 부숴지거나 갈기 위해 들쭉날쭉한 능선을 가진 넓은 에나멜 판으로 치아를 변형시켰다. 잉어와 미꾸라지는 오직 인두 치아만을 가지고 있다. 해마, 관상어, 철갑상어는 어떤 종류의 이빨도 가지고 있지 않다. 물고기에서, Hox 유전자 발현은 치아 개시를 위한 메커니즘을 조절한다.[30][31]
하지만 상어는 일생 동안 완전히 다른 메커니즘을 통해 계속해서 새로운 치아를 생산한다.[32][33][34] 상어 이빨은 혀 근처의 변형된 비늘에서 형성되어 결국 빠져 나올 때까지 줄지어 턱을 바깥쪽으로 움직인다.[35] 그들의 비늘은 피부 틀니클이라고 불리며, 치아는 동음이의 기관이다.[36]
상어 이빨은 턱에 직접 부착하기보다는 잇몸에 박혀 평생 끊임없이 교체된다. 교체용 치아가 여러 줄로 늘어선 형태는 턱 안쪽의 홈에서 자라고 컨베이어 벨트 위처럼 꾸준히 앞으로 나아간다. 어떤 상어들은 일생동안 3만개 혹은 그 이상의 치아를 잃는다. 치아교체율은 8~10일마다 1회씩에서 수개월까지 다양하다. 대부분의 종에서, 치아는 쿠키커터 상어에서 관찰되는 전체 행의 동시 교체와는 반대로 한 번에 하나씩 교체된다.[37]
이빨 모양은 상어의 식단에 따라 달라지는데, 연체동물과 갑각류를 먹는 이들은 으스러지는 데 사용되는 촘촘하고 납작한 이빨을 가지고 있고, 물고기를 먹는 이들은 바늘처럼 생긴 이빨을 가지고 있으며, 포유류와 같은 큰 먹이를 먹는 사람들은 톱니 모양의 모서리를 톱니 모양으로 하고, 삼각형의 위쪽 이빨을 가리켜 자르는 상어의 식단에 따라 치아 모양이 달라진다. 돌묵상어와 같은 플랑크톤 피더들의 이빨은 작고 기능적이지 않다.[38]
- 턱과 이빨

멸종된 카르샤로돈 메가로돈의 턱 재건, 1909년

손등 광선은 게, 새우, 작은 물고기를 먹기에 적합한 치아를 가지고 있다.

호랑이 상어 이빨은 살을 통해 톱질하도록 비스듬히 하고 톱질한다.





예
연어


수컷 연어는 산란 과정에서 턱을 개조하는 경우가 많아 굴곡이 뚜렷하다. 이 갈고리 모양의 턱은 kypes라고 불린다. kype의 목적이 완전히 명확하지는 않지만, 상대의 꼬리 밑부분(카우데자네이루) 주위에 고정시켜 지배력을 확립하는 데 사용할 수 있다.[39][40]
치클리드

일반적으로 물고기 턱은 척추동물과 마찬가지로 양대칭성을 보인다. 기생 스케일 먹는 시클리드 페리스소두스 마이크로레피스에서는 예외가 발생한다. 이 물고기의 턱은 두 가지 뚜렷한 형태학적 형태로 나타난다. 하나의 형태는 턱이 왼쪽으로 비틀어져 있어 희생자의 오른쪽 옆구리에 있는 비늘을 더 쉽게 먹을 수 있다. 다른 형태는 턱이 오른쪽으로 꼬여 있어 희생자의 왼쪽 옆구리에 있는 비늘을 쉽게 먹을 수 있다. 모집단에서 두 형태에 대한 상대적 풍부함은 주파수 의존적 선택에 의해 조절된다.[41][42][43]
치클리드에서는 일반적으로 구강 치아와 인두 치아가 다른 종류의 먹이를 처리할 수 있는 방법에서 다른 종에 따라 다르다. 1차 구강턱은 음식을 담는 데 사용되는 치아를 포함하고 있으며, 2차 구강턱은 씹는 도구 역할을 하는 치아를 가지고 있다.


이것은 다른 영양 전략을 가능하게 하고, 이것 때문에, 시클리드들은 다른 서식지를 식민지화할 수 있다. 하부 인두턱의 구조적 다양성은 많은 시클리드 종이 발생하는 이유 중 하나가 될 수 있다. 융합적 진화는 시클리드 방사선의 과정에서 서로 다른 영양 틈새와 동시에 일어났다.[44] 인두턱 기구는 상판 2개와 하판 1개로 구성되며, 모두 크기와 유형이 다른 틀니를 가지고 있다.[45] 아랫 인두의 구조는 종종 그 종의 음식종과 연관된다.[46]
조개를 깨기 위해서는 상당한 힘이 발생해야 하는데, 이것이 연체동물(예: 시클리드 베이스, Crenicichla minuano)을 먹고 사는 시클리드(cichlid bass, Crenicichla minuano)는 어금니형 치아와 턱뼈가 강화되는 이유다. 포탄으로 무장하지 않은 먹이를 잡고 물기 위해서는 포식자들이 원뿔형의 구부러진 등 이빨을 필요로 한다.[47] 초식성 치클리드 또한 치아의 구조적 차이를 가지고 있다. 조류에 특화된 시클리드들은 작은 원뿔형 치아를 갖는 경향이 있다. 꼬투리나 씨앗을 먹고 사는 종은 음식을 씹기 위해 큰 원뿔 치아를 필요로 한다.[48]
기타



스톱라이트 루즈조우는 전세계적으로 심해에서 발견되는 작은 물고기들이다. 크기에 비해 그들은 가장 넓은 물고기들 중 하나를 가지고 있다. 아래턱에는 에트모이드 막(바닥)이 없고 경첩과 변형된 혀뼈만으로 부착되어 있다. 턱 앞쪽에는 송곳니 같은 큰 이빨이 몇 개 있고 그 뒤에는 작은 철조망이 여러 개 있다. 식도로 음식을 유도하는 역할을 하는 여러 그룹의 인두 치아가 있다.[49][50]
또 다른 심해어인 펠리컨 뱀장어는 몸보다 턱이 더 크다. 턱에는 작은 이빨이 늘어서 있고 느슨하게 경첩이 달려 있다. 그들은 뱀장어 그 자체보다 큰 물고기를 삼킬 정도로 활짝 연다.
디스티코돈티과는 육식동물인 긴 턱을 가진 일반과 초식동물이나 매우 작은 생물의 포식자인 비순식적인 위턱을 가진 일반으로 나눌 수 있는 민물고기과의 일종이다.[51]
진화

초기 척추동물 턱의 외형은 "중요한 혁신"[53]과 "아마도 척추동물 역사상 가장 심오하고 급진적인 진화 단계"로 묘사되어 왔다.[4][5] 턱이 없는 물고기는 턱이 있는 물고기보다 생존에 어려움이 많았고, 대부분의 턱이 없는 물고기는 트라이아스기 시대에 멸종되었다. 그러나 턱이 없는 해파리와 등잔류인 사이클로스테놈에 대한 연구는 초기 턱이 진화하면서 일어났어야 할 척추동물 두개골의 깊은 리모델링에 대한 통찰력을 거의 만들어내지 못했다.[54][55]
관습적인 견해는 턱이 아가미 아치와 동음이의어라는 것이다.[56] 턱이 없는 물고기에서는 아가미가 입 뒤에서 열렸고, 아가미는 수척한 원소에 의해 지탱되었다. 이들 원소의 첫 번째 세트는 입을 둘러 턱을 형성했다. 아가미를 받치고 있는 두 번째 배아 아치의 윗부분은 턱이 있는 물고기의 효모판형 뼈가 되었고, 이것은 두개골을 지탱하고 따라서 턱을 두개골과 연결시킨다.[57] 효만디불라는 대부분의 물고기의 효이드 지역에서 발견되는 뼈의 집합체다. 그것은 보통 텔레오스트의 경우 턱이나 수술실을 정지시키는 역할을 한다.[58]



턱이 있는 척추동물의 전구체는 오랫동안 멸종된 뼈(기갑)가 없는 물고기, 이른바 타조류라는 것이 이제 인정되고 있다.[59][60] 턱을 가진 가장 일찍 알려진 물고기는 현재 멸종된 플라코다름과[61] 가시가 있는 상어들이다.[62]
플라코다룸은 약 4억 3천만년 전 실루리아인 시대에 처음으로 화석 기록에 나타난, 몸 앞쪽에 무겁게 갑옷을 입은 물고기의 한 종류였다. 처음에 그것들은 데본기 동안 현저하게 다양화되면서 매우 성공적이었다. 그들은 약 3억 6천만년 전, 그 기간이 끝날 무렵에 멸종되었다.[63] 이들의 가장 큰 종인 덩클레오스테우스 테렐리는 최대 10m(33ft),[64][65] 무게는 3.6t(4.0t)이었다.[66] 그것은 두개골, 흉부 실드, 아래턱 및 턱 근육 사이의 연결부를 이동 가능한 관절에 의해 결합하는 턱 개구부를 위한 4개의 막대 연결 메커니즘을 가지고 있었다.[67][68] 이 메커니즘은 던클레오스테우스 테렐리가 빠른 속도로 턱을 벌려 20밀리초 안에 턱을 벌리고 모든 과정을 50~60밀리초 안에 마칠 수 있게 해 주었는데, 이는 흡인력을 사용하여 먹이 포획을 돕는 현대 어류들과 비교된다.[67] 그들은 또한 턱을 닫을 때 높은 물림력을 발생시킬 수 있는데, 가장 큰 개인에서 가장자리에는 6,000N (1,350f lb), 가장자리에는 7,400 N (1,660 lbf)로 추정된다.[68] 이 지역에서 발생한 압력은 덩클로스 테렐리가 자유 수영, 절지동물, 암모니아이트, 그리고 다른 플라코더와 같은 무기류 먹이에 완벽하게 적응했다는 것을 암시하는 큐티클이나 피부 무장을[67] 관통하거나 구멍을 뚫을 수 있을 정도로 충분히 높았다.[68]
가시가 있는 상어는 실루리아기 동안 플라코마들과 거의 동시에 화석 기록에도 나타난 또 다른 종류의 물고기였다. 그것들은 보통 20 센티미터 미만인 대부분의 플라스마보다 작았다. 가시가 있는 상어는 플라코마만큼 다양화되지는 않았지만, 약 2억 9천만년 전에 얼리 퍼미언으로 훨씬 더 오래 살아남았다.[69]
턱에 의해 제공되는 원래의 선택적 이점은 먹이를 주는 것과 관련이 있는 것이 아니라 호흡 효율의 증가와 관련이 있을 수 있다.[70] 턱은 현대 어류와 양서류에서 여전히 관찰 가능한 부칼 펌프에서 사용되었는데, 양서류의 경우 "볼로 숨을 쉬는 것"을 사용하여 물고기의 아가미를 가로질러 물을 펌핑하거나 공기를 폐로 주입한다. 진화 시간이 지나면서, 먹이를 주는 데 있어서 턱을 더 친숙하게 사용하는 것이 척추동물들에게 선택되었고 매우 중요한 기능이 되었다. 많은 텔레오스트 물고기는 흡인용 먹이를 주는 턱과 턱 돌출부를 실질적으로 변형시켜, 수십 개의 뼈가 포함된 매우 복잡한 턱을 만들어 낸다.[71]
죠스는 물고기의 아가미를 지탱하는 인두 아치에서 유래된 것으로 생각된다. 이 아치들 중 가장 앞쪽에 있는 두 개는 턱 자체(효만디불라 참조)와 턱을 뇌케이스에 고정시키고 기계적 효율을 높이는 효이드 아치가 된 것으로 생각된다. 이 이론을 직접적으로 뒷받침할 화석 증거는 없지만, 7개의 아치를 가진 현존 턱(Gnathostome)과 9개의 아치를 가진 원시 턱 없는 척추동물(Agnatha)에서 보이는 인두 아치의 수에 비추어 보면 타당하다.
메켈의 연골은 척추동물의 하악골(하악턱)이 진화한 연골 조각이다. 원래 그것은 초기 물고기의 첫 번째 길 아치(앞쪽이 가장 가까운 곳)를 지탱하는 두 개의 포장마차 중 아래쪽이었다. 그리고 나서 그것은 더 길고 튼튼해졌고, 발달한 턱을 닫을 수 있는 근육을 얻었다.[72] 초기 어류나 콘드리히테얀(상어와 같은 갑상어류)에서는 메켈의 연골이 계속해서 아래턱의 주성분이 되었다. 그러나 성인 형태의 골수족과 그 후손들(암피비아인, 파충류, 새, 포유류)에서 연골은 뼈로 덮여 있었다. 비록 그들의 배아에서 턱은 처음에는 메켈의 연골로 발달하지만 말이다. 테트라포드에서 연골은 턱 뒷부분에서 부분적으로 오물화되어(뼈로 변화) 관절뼈가 되며, 포유류를 제외한 모든 테트라포드에서 턱관절의 일부를 형성한다.[72]
참고 항목
메모들
- ^ Jump up to: a b Fraser, G. J.; Hulsey, C. D.; Bloomquist, R. F.; Uyesugi, K.; Manley, N. R.; Streelman, J. T. (2009). "An ancient gene network is co-opted for teeth on old and new jaws". PLOS Biology. 7 (2): e1000031. doi:10.1371/journal.pbio.1000031. PMC 2637924. PMID 19215146.
- ^ Mabuchi, K.; Miya, M.; Azuma, Y.; Nishida, M. (2007). "Independent evolution of the specialized pharyngeal jaw apparatus in cichlid and labrid fishes". BMC Evolutionary Biology. 7 (1): 10. doi:10.1186/1471-2148-7-10. PMC 1797158. PMID 17263894.
- ^ Alfaro, M. E.; Brock, C. D.; Banbury, B. L.; Wainwright, P. C. (2009). "Does evolutionary innovation in pharyngeal jaws lead to rapid lineage diversification in labrid fishes?". BMC Evolutionary Biology. 9 (1): 255. doi:10.1186/1471-2148-9-255. PMC 2779191. PMID 19849854.
- ^ Jump up to: a b Gai, Z.; Zhu, M. (2012). "The origin of the vertebrate jaw: Intersection between developmental biology-based model and fossil evidence". Chinese Science Bulletin. 57 (30): 3819–3828. Bibcode:2012ChSBu..57.3819G. doi:10.1007/s11434-012-5372-z.
- ^ Jump up to: a b Maisey, J. G. (2000). Discovering Fossil Fishes. Westview Press. pp. 1–223. ISBN 978-0-8133-3807-1.
- ^ Jump up to: a b Westneat, Mark W. (September 1990). "Feeding mechanics of teleost fishes (Labridae; Perciformes): A test of four-bar linkage models". Journal of Morphology. 205 (3): 269–295. doi:10.1002/jmor.1052050304. PMID 29865760. S2CID 46933606.
- ^ Olsen, Aaron M.; Camp, Ariel L.; Brainerd, Elizabeth L. (15 December 2017). "The opercular mouth-opening mechanism of largemouth bass functions as a 3D four-bar linkage with three degrees of freedom". Journal of Experimental Biology. 220 (24): 4612–4623. doi:10.1242/jeb.159079. PMID 29237766.
- ^ Muller, M (29 May 1996). "A novel classification of planar four-bar linkages and its application to the mechanical analysis of animal systems". Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences. 351 (1340): 689–720. Bibcode:1996RSPTB.351..689M. doi:10.1098/rstb.1996.0065. PMID 8927640.
- ^ Jump up to: a b c d 로머 & 파슨스 1977, 페이지 173–177
- ^ 하악골은 적어도 1858년 초판 그레이 아나토미(Gray's Anatomy)로 거슬러 올라가는 구식 용어지만, 하악골이라고도 불리는 일부 출처에도 있다.
- ^ Jump up to: a b c 로머 & 파슨스 1977, 페이지 244–247
- ^ 1989년 OED 2판
- ^ "maxilla". Merriam-Webster Online Dictionary.
- ^ Jump up to: a b 로머 & 파슨스 1977, 페이지 217–243
- ^ Wueringer, B. E.; Squire, L., Jr; Kajiura, S. M.; Hart, N. S.; Collin, S. P. (2012). "The function of the sawfish's saw". Current Biology. 22 (5): R150–R151. doi:10.1016/j.cub.2012.01.055. PMID 22401891.
- ^ Westneat, M. W. (1 November 2004). "Evolution of Levers and Linkages in the Feeding Mechanisms of Fishes". Integrative and Comparative Biology. 44 (5): 378–389. doi:10.1093/icb/44.5.378. PMID 21676723.
- ^ Benton, Michael (2005). "The Evolution of Fishes After the Devonian". Vertebrate Palaeontology (3rd ed.). John Wiley & Sons. pp. 175–84. ISBN 978-1-4051-4449-0.
- ^ Jump up to: a b c d e f Motta, Philip Jay (23 February 1984). "Mechanics and Functions of Jaw Protrusion in Teleost Fishes: A Review". Copeia. 1984 (1): 1–18. doi:10.2307/1445030. JSTOR 1445030.
- ^ Jump up to: a b Liem, Karel F (February 1980). "Adaptive Significance of Intra- and Interspecific Differences in the Feeding Repertoires of Cichlid Fishes". American Zoologist. 20 (1): 295–314. doi:10.1093/icb/20.1.295.
- ^ Jump up to: a b Wainwright, Peter C.; Alfaro, Michael E.; Bolnick, Daniel I.; Hulsey, C. Darrin (2005). "Many-to-One Mapping of Form to Function: A General Principle in Organismal Design?". Integrative and Comparative Biology. 45 (2): 256–262. doi:10.1093/icb/45.2.256. PMID 21676769.
- ^ Chisholm, Hugh, ed. (1911). . Encyclopædia Britannica. 28 (11th ed.). Cambridge University Press. p. 839.
- ^ Westneat, Mark W.; Wainwright, Peter C. (November 1989). "Feeding mechanism ofEpibulus insidiator (Labridae; Teleostei): Evolution of a novel functional system". Journal of Morphology. 202 (2): 129–150. doi:10.1002/jmor.1052020202. PMID 29865677. S2CID 46933765.
- ^ Mehta, Rita S.; Wainwright, Peter C. (6 September 2007). "Raptorial jaws in the throat help moray eels swallow large prey". Nature. 449 (7158): 79–82. Bibcode:2007Natur.449...79M. doi:10.1038/nature06062. PMID 17805293. S2CID 4384411.
- ^ Graham, Anthony; Richardson, Jo (2012). "Developmental and evolutionary origins of the pharyngeal apparatus". EvoDevo. 3 (1): 24. doi:10.1186/2041-9139-3-24. PMC 3564725. PMID 23020903.
- ^ Hamlett, W. C. (1999f). Sharks, Skates and Rays: The Biology of Elasmobranch Fishes. Johns Hopkins University Press. ISBN 978-0-8018-6048-5. OCLC 39217534.
- ^ Martin, R. Aidan. "Skeleton in the Corset". ReefQuest Centre for Shark Research. Retrieved 21 August 2009.
- ^ Wilga, C. D. (2005). "Morphology and evolution of the jaw suspension in lamniform sharks". Journal of Morphology. 265 (1): 102–119. doi:10.1002/jmor.10342. PMID 15880740. S2CID 45227734.
- ^ Wilga, C. D.; Motta, P. J.; Sanford, C. P. (2007). "Evolution and ecology of feeding in elasmobranchs". Integrative and Comparative Biology. 47 (1): 55–69. doi:10.1093/icb/icm029. PMID 21672820.
- ^ Motta, Philip J.; Huber, Daniel R. (2012). "Prey Capture Behavior and Feeding Mechanisms of Elasmobranchs". In Carrier, J. C.; Musick, J. A.; Heithaus, M. R. (eds.). Biology of Sharks and Their Relatives (Second ed.). CRC Press. pp. 153–210. ISBN 978-1-4398-3924-9.
- ^ Fraser GJ, Hulsey CD, Bloomquist RF, Uyesugi K, Manley NR, Streelman JT (February 2009). Jernvall J (ed.). "An Ancient Gene Network Is Co-opted for Teeth on Old and New Jaws". PLOS Biology. 7 (2): e31. doi:10.1371/journal.pbio.1000031. PMC 2637924. PMID 19215146.
- ^ Fraser GJ, Bloomquist RF, Streelman JT (2008). "A periodic pattern generator for dental diversity". BMC Biology. 6: 32. doi:10.1186/1741-7007-6-32. PMC 2496899. PMID 18625062.
- ^ 데이브 애벗, 샤크스 여기서 발견
- ^ Boyne, Philip J. (March 1970). "Study of the Chronologic Development and Eruption of Teeth in Elasmobranchs". Journal of Dental Research. 49 (3): 556–560. doi:10.1177/00220345700490031501. PMID 5269110. S2CID 34954175.
- ^ Sasagawa I (June 1989). "The fine structure of initial mineralisation during tooth development in the gummy shark, Mustelus manazo, Elasmobranchia". Journal of Anatomy. 164: 175–87. PMC 1256608. PMID 2606790.
- ^ 마이클 E. 윌리엄스, 죠스: 초창기, 여기에서 [1].
- ^ Luan, X.; Ito, Y.; Diekwisch, T.G.H. (2005). "Evolution and development of Hertwig's epithelial root sheath". Developmental Dynamics. 235 (5): 1167–1180. doi:10.1002/dvdy.20674. PMC 2734338. PMID 16450392.
- ^ Martin, R. Aidan. "Skin of the Teeth". Retrieved 28 August 2007.
- ^ Gilbertson, Lance (1999). Zoology Laboratory Manual. New York: McGraw-Hill Companies, Inc. ISBN 978-0-07-237716-3.
- ^ Witten, P. E.; Hall, B. K. (2003). "Seasonal changes in the lower jaw skeleton in male Atlantic salmon (Salmo salar L.): remodelling and regression of the kype after spawning". Journal of Anatomy. 203 (5): 435–450. doi:10.1046/j.1469-7580.2003.00239.x. PMC 1571185. PMID 14635799.
- ^ Groot, C.; Margolis, L. (1991). Pacific salmon life histories. UBC Press. p. 143. ISBN 978-0-7748-0359-5.
- ^ Jump up to: a b Lee, H. J.; Kusche, H.; Meyer, A. (2012). "Handed Foraging Behavior in Scale-Eating Cichlid Fish: Its Potential Role in Shaping Morphological Asymmetry". PLOS ONE. 7 (9): e44670. Bibcode:2012PLoSO...744670L. doi:10.1371/journal.pone.0044670. PMC 3435272. PMID 22970282.
- ^ Hori, M. (1993). "Frequency-dependent natural selection in the handedness of scale-eating cichlid fish". Science. 260 (5105): 216–219. Bibcode:1993Sci...260..216H. doi:10.1126/science.260.5105.216. PMID 17807183. S2CID 33113282.
- ^ Stewart, T. A.; Albertson, R. C. (2010). "Evolution of a unique predatory feeding apparatus: functional anatomy, development and a genetic locus for jaw laterality in Lake Tanganyika scale-eating cichlids". BMC Biology. 8 (1): 8. doi:10.1186/1741-7007-8-8. PMC 2828976. PMID 20102595.
- ^ Muschick, Moritz; Indermaur, Adrian; Salzburger, Walter (December 2012). "Convergent Evolution within an Adaptive Radiation of Cichlid Fishes". Current Biology. 22 (24): 2362–2368. doi:10.1016/j.cub.2012.10.048. PMID 23159601. S2CID 18363916.
- ^ Casciotta, Jorge R.; Arratia, Gloria (July 1993). "Jaws and teeth of american cichlids (Pisces: Labroidei)". Journal of Morphology. 217 (1): 1–36. doi:10.1002/jmor.1052170102. PMID 29865430. S2CID 46927413.
- ^ Burress, Edward D. (April 2015). "Cichlid fishes as models of ecological diversification: patterns, mechanisms, and consequences". Hydrobiologia. 748 (1): 7–27. doi:10.1007/s10750-014-1960-z. S2CID 15963069.
- ^ Burress, Edward D.; Duarte, Alejandro; Gangloff, Michael M.; Siefferman, Lynn (January 2013). "Isotopic trophic guild structure of a diverse subtropical South American fish community". Ecology of Freshwater Fish. 22 (1): 66–72. doi:10.1111/eff.12002.
- ^ Genner, Martin J.; Turner, George F.; Hawkins, Stephen J. (1999). "Foraging of Rocky Habitat Cichlid Fishes in Lake Malawi: Coexistence through Niche Partitioning?". Oecologia. 121 (2): 283–292. Bibcode:1999Oecol.121..283G. doi:10.1007/s004420050930. JSTOR 4222466. PMID 28308568. S2CID 13285836.
- ^ Kenaley, C. P. (2007). "Revision of the Stoplight Loosejaw Genus Malacosteus (Teleostei: Stomiidae: Malacosteinae), with Description of a New Species from the Temperate Southern Hemisphere and Indian Ocean". Copeia. 2007 (4): 886–900. doi:10.1643/0045-8511(2007)7[886:ROTSLG]2.0.CO;2.
- ^ Sutton, Tracey T. (November 2005). "Trophic ecology of the deep-sea fish Malacosteus niger (Pisces: Stomiidae): An enigmatic feeding ecology to facilitate a unique visual system?". Deep Sea Research Part I: Oceanographic Research Papers. 52 (11): 2065–2076. Bibcode:2005DSRI...52.2065S. doi:10.1016/j.dsr.2005.06.011.
- ^ Nelson, Joseph, S. (2006). Fishes of the World. John Wiley & Sons, Inc. ISBN 978-0-471-25031-9.
- ^ Benton 2005. CATEREFBenton
- ^ Kimmel, C. B.; Miller, C. T.; Keynes, R. J. (2001). "Neural crest patterning and the evolution of the jaw". Journal of Anatomy. 199 (1&2): 105–119. doi:10.1017/S0021878201008068. PMC 1594948. PMID 11523812.
- ^ Janvier, P. (2007). "Homologies and Evolutionary Transitions in Early Vertebrate History". In Anderson, J. S.; Sues, H.-D. (eds.). Major Transitions in Vertebrate Evolution. Indiana University Press. pp. 57–121. ISBN 978-0-253-34926-2.
- ^ Khonsari, R. H.; Li, B.; Vernier, P.; Northcutt, R. G.; Janvier, P. (2009). "Agnathan brain anatomy and craniate phylogeny". Acta Zoologica. 90 (s1): 52–68. doi:10.1111/j.1463-6395.2008.00388.x. S2CID 56425436.
- ^ 예를 들어 (1) 두 개의 뼈 세트는 모두 신경 파고 세포(대부분의 다른 뼈와 같이 중피 조직이 아닌)로 만들어졌으며, (2) 두 구조물은 모두 앞으로 구부러지고 가운데에 경첩되는 상하의 막대를 형성하며, (3) 턱의 근골은 턱 없는 물고기의 아가리와 동질적으로 보인다.(길버트 2000)
- ^ Gilbert (2000). Evolutionary Embryology.
- ^ Clack, J. A. (1994). "Earliest known tetrapod braincase and the evolution of the stapes and fenestra ovalis". Nature. 369 (6479): 392–394. Bibcode:1994Natur.369..392C. doi:10.1038/369392a0. S2CID 33913758.
- ^ Donoghue, P. C.; Purnell, M. A. (2005). "Genome duplication, extinction and vertebrate evolution". Trends in Ecology & Evolution. 20 (6): 312–319. doi:10.1016/j.tree.2005.04.008. PMID 16701387.
- ^ Forey, P. L.; Janvier, P. (1993). "Agnathans and the origin of jawed vertebrates". Nature. 361 (6408): 129–134. Bibcode:1993Natur.361..129F. doi:10.1038/361129a0. S2CID 43389789.
- ^ "Placodermi: Overview". Palaeos. Retrieved 10 December 2014.
- ^ "Acanthodii". Palaeos. Retrieved 10 December 2014.
- ^ "More About Placoderms". Devonian Times. 9 July 2005.
- ^ "Ancient Fish With Killer Bite". Science News. 19 May 2009.
- ^ Palmer, D., ed. (1999). The Marshall Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals. London: Marshall Editions. p. 33. ISBN 978-1-84028-152-1.
- ^ "Monster fish crushed opposition with strongest bite ever". The Sydney Morning Herald. 30 November 2006.
- ^ Jump up to: a b c Anderson, P.S.L.; Westneat, M. (2007). "Feeding mechanics and bite force modelling of the skull of Dunkleosteus terrelli, an ancient apex predator". Biology Letters. 3 (1): 76–79. doi:10.1098/rsbl.2006.0569. PMC 2373817. PMID 17443970.
- ^ Jump up to: a b c Anderson, P. S. L.; Westneat, M. (2009). "A biomechanical model of feeding kinematics for Dunkleosteus terrelli (Arthrodira, Placodermi)". Paleobiology. 35 (2): 251–269. doi:10.1666/08011.1. S2CID 86203770.
- ^ "More About Acanthodians (spiny fins)". Devonian Times. 9 July 2005.
- ^ Smith, M.M.; Coates, M.I. (2000). "10. Evolutionary origins of teeth and jaws: developmental models and phylogenetic patterns". In Teaford, Mark F.; Smith, Moya Meredith; Ferguson, Mark W.J. (eds.). Development, function and evolution of teeth. Cambridge: Cambridge University Press. p. 145. ISBN 978-0-521-57011-4.
- ^ Britt, Robert Roy (28 November 2006). "Prehistoric Fish Had Most Powerful Jaws". Live Science.
- ^ Jump up to: a b "The Gill Arches: Meckel's Cartilage". palaeos. Retrieved 4 December 2014.
기타리딩
- Benton, Michael J (2009). Vertebrate Palaeontology (3rd ed.). John Wiley & Sons. ISBN 978-1-4051-4449-0.
- Botella, H.; Blom, H.; Dorka, M.; Ahlberg, P. E.; Janvier, P. (2007). "Jaws and teeth of the earliest bony fishes". Nature. 448 (7153): 583–586. Bibcode:2007Natur.448..583B. doi:10.1038/nature05989. PMID 17671501. S2CID 4337868.
- Compagnucci, C; Debiais-Thibaud, M; Coolen, M; Fish, J; Griffin, J N; Bertocchini, F; Minoux, M; Rijli, F M; Borday-Birraux, V; Casane, D; Mazanc, S; Depew, M J (2013). "Pattern and polarity in the development and evolution of the gnathostome jaw: Both conservation and heterotopy in the branchial arches of the shark, Scyliorhinus canicula". Developmental Biology. 377 (2): 428–448. doi:10.1016/j.ydbio.2013.02.022. PMID 23473983.
- Depew, M J; Lufkin, T; Rubenstein, J L (2002). "Specification of jaw subdivisions by Dlx genes". Science. 298 (5592): 381–385. doi:10.1126/science.1075703. PMID 12193642. S2CID 10274300.
- Forey, Peter; Janvier, Philippe (2000). "Agnathans and the origin of jawed vertebrates". In Gee, Henry (ed.). Shaking the tree: readings from Nature in the history of life. USA: University of Chicago Press; Nature/Macmillan Magazines. pp. 251–266. ISBN 978-0-226-28497-2.
- Gilbert, Scott F. (2000). "The anatomical tradition: Evolutionary Embryology: Embryonic homologies". Developmental Biology. Sunderland (MA): Sinauer Associates, Inc. (NCBI). Retrieved 9 April 2018. (3번과 4번 파라사, 가장 유명한 사례 중 하나...)
- Hulsey, CD; Fraser, GJ; Streelman, JT (2005). "Evolution and development of complex biomechanical systems: 300 million years of fish jaws". Zebrafish. 2 (4): 243–257. CiteSeerX 10.1.1.210.7203. doi:10.1089/zeb.2005.2.243. PMID 18248183.
- Koentges, G; Matsuoka, T (2002). "Jaws of the fates". Science. 298 (5592): 371–373. doi:10.1126/science.1077706. PMID 12376690. S2CID 20212436.
- Lingham-Soliar, Theagarten (2014). "The First Vertebrates, Jawless Fishes, the Agnathans". The Vertebrate Integument Volume 1. pp. 11–31. doi:10.1007/978-3-642-53748-6_2. ISBN 978-3-642-53747-9.
- Lingham-Soliar, T. (2014). "The Earliest Jawed Vertebrates, the Gnathostomes". The Vertebrate Integument. 1. Springer. pp. 33–58. ISBN 978-3-642-53748-6.
- Mallatt, J. (2008). "The origin of the vertebrate jaw: Neoclassical ideas versus newer, development-based ideas". Zoological Science. 25 (10): 990–998. doi:10.2108/zsj.25.990. PMID 19267635. S2CID 3104126.
- Mehta, Rita S.; Wainwright, Peter C. (May 2008). "Functional morphology of the pharyngeal jaw apparatus in moray eels". Journal of Morphology. 269 (5): 604–619. doi:10.1002/jmor.10612. PMID 18196573. S2CID 17013964.
- Muschick, M.; Salzburger, W. (2013). "Pharyngeal jaws and their evolutionary, ecological and behavioural significance" (PDF). In Muschick, Moritz (ed.). Convergence and plasticity in the adaptive radiation of cichlid fishes (PhD thesis). University of Basel. pp. 13–37.
- Oisi, Y; Ota, K G; Kuraku, S; Fujimoto, S; Kuratani, S (2013). "Craniofacial development of hagfishes and the evolution of vertebrates". Nature. 493 (7431): 175–180. Bibcode:2013Natur.493..175O. doi:10.1038/nature11794. PMID 23254938. S2CID 4403344.
- Romer, Alfred Sherwood; Parsons, Thomas S. (1977). The Vertebrate Body. Philadelphia, PA: Holt-Saunders International. ISBN 978-0-03-910284-5.
- Soukup, V; Horácek, I; Cerny, R (2013). "Development and evolution of the vertebrate primary mouth". Journal of Anatomy. 222 (1): 79–99. doi:10.1111/j.1469-7580.2012.01540.x. PMC 3552417. PMID 22804777.
- Wainwright, P. C. (2006). "Functional Morphology of the Pharyngeal Jaw Apparatus". In Shadwick, R. E.; Lauder, G. V. (eds.). Fish Biomechanics. Fish Physiology. 23. Academic Press. pp. 77–102. ISBN 978-0-08-047776-3. 전체 보기
- Westneat, M. W. (2006). "Skull Biomechanics and Suction Feeding in Fishes". In Shadwick, R. E.; Lauder, G. V. (eds.). Fish Biomechanics. Fish Physiology. 23. Academic Press. pp. 29–76. ISBN 978-0-08-047776-3.
- Westneat, Mark W. (2004). "Evolution of levers and linkages in the feeding mechanisms of fishes". Integrative and Comparative Biology. 44 (5): 378–389. doi:10.1093/icb/44.5.378. PMID 21676723.
외부 링크
- "Moray Eels Are Uniquely Equipped to Pack Big Prey Into Their Narrow Bodies" (Press release). National Science Foundation. 5 September 2007.
- Myers, PZ (13 March 2007). "Evolution of the jaw". Pharyngula.
- Barford, Eliot (25 September 2013). "Ancient fish face shows roots of modern jaw". News. Nature.
- Zhu, Min; Yu, Xiaobo; Erik Ahlberg, Per; Choo, Brian; Lu, Jing; Qiao, Tuo; Qu, Qingming; Zhao, Wenjin; Jia, Liantao; Blom, Henning; Zhu, You’an (2013). "A Silurian placoderm with osteichthyan-like marginal jaw bones". Nature. 502 (7470): 188–193. Bibcode:2013Natur.502..188Z. doi:10.1038/nature12617. PMID 24067611. S2CID 4462506.


