규조류
Diatom| 규조류 시간 범위: | |
|---|---|
 | |
| 남극의 연간 해빙 결정 사이에 서식하는 해양 규조 표본의 광현미경 검사로 크기와 모양이 다양함을 알 수 있습니다. | |
| 과학적 분류 | |
| Clade: | SAR |
| 인크루: | 헤테로콘타 |
| 문: | 오크로피타 |
| 하위문: | 카키스타 |
| 클래스: | 바실라리오피스과 단게어드, 1933년[1] |
| 동의어 | |
규조류(Neo-Latin diatoma)[a]는 세계의 바다, 수로, 토양에서 발견되는 여러 속, 특히 미세 조류로 구성된 큰 그룹의 모든 구성원을 말합니다.살아있는 규조류는 지구 바이오매스의 상당 부분을 차지한다:[10][11] 그들은 매년 지구에서 생산되는 산소의 약 20-50%를 생산하고,[12] 그들이 살고 있는 물에서 매년 67억 미터 톤 이상의 실리콘을 흡수하며, 바다에서 발견되는 유기 물질의 거의 절반을 차지한다.죽은 규조개 껍데기는 해저의 0.5마일(800m) 깊이에 이를 수 있으며 아마존 유역 전체가 아프리카 사하라에서 불어오는 대서양 횡단 바람에 의해 운반되는 2,700만 톤의 규조개 분진에 의해 매년 수정되는데, 이 중 대부분은 한때 민물 [13][14]호수로 이루어진 보델레 제방으로부터 온다.
규조류는 단세포 유기체이다: 그들은 리본, 부채꼴, 지그재그 또는 별 모양을 취할 수 있는 단세포 또는 군집 형태로 발생한다.개별 셀의 크기는 2마이크로미터에서 [15]200마이크로미터까지 다양합니다.충분한 영양소와 햇빛이 있을 때, 살아있는 규조들의 집합은 무성 다분열에 의해 약 24시간마다 두 배씩 증가한다. 개별 세포의 최대 수명은 약 6일이다.[16]규조류는 두 가지 뚜렷한 형태를 가지고 있는데, 몇 개의 규조(중심 규조)는 반지름 대칭인 반면, 대부분의 규조(페네이트 규조)는 대체로 좌우 대칭이다.규조 해부학의 독특한 특징은 규조체가 실리카로 만들어진 세포벽으로 둘러싸여 있다는 것이다.[17]이러한 좌절은 광나노 구조로 인해 구조적인 색을 띠며, 이를 "바다의 보석"과 "살아있는 오팔"로 묘사하도록 유도한다.규조류에서의 움직임은 주로 해류와 바람에 의해 유발되는 물의 난류의 결과로 수동적으로 일어난다. 그러나 중심 규조류의 수컷 배우자는 편모를 가지고 있어 암컷 배우자를 찾기 위한 활발한 움직임을 가능하게 한다.식물과 비슷하게, 규조류는 광합성에 의해 빛 에너지를 화학 에너지로 전환하지만, 그들의 엽록체는 다른 방식으로 [18]획득되었다.
자가영양 유기체의 경우 특이하게, 규조류는 동물과 공유하는 요소 순환을 가지고 있지만, 이 순환은 규조체의 다른 대사 끝에 사용됩니다.로팔로디아과는 또한 구상체라고 불리는 시아노박테리아 내분비온을 가지고 있다.이 내심비온은 광합성 성질을 잃었지만 질소를 고정하는 능력은 유지되어 규조류가 대기 중 [19]질소를 고정할 수 있게 되었다.질소 고정성 시아노박테리아와 공생하는 다른 규조류는 헤미아울루스속, 리조솔레니아속 및 [20]채토케로스속이다.
규조 연구는 식물학의 한 분야이다.규조류는 원핵생물 및 박테리아로부터 분리되는 핵 외피 결합 세포핵을 가진 유기체인 진핵생물들로 분류된다.디아톰은 플랑크톤이라고 불리는 플랑크톤의 한 종류로 플랑크톤 중에서 가장 흔하다.규조류는 또한 해저 기질, 부유 잔해, 대식물에 부착되어 자란다.그들은 페리피톤 [21]공동체의 필수적인 구성 요소를 구성한다.또 다른 분류는 플랑크톤을 크기에 따라 8가지로 나눈다: 이 체계에서, 규조류는 미세 조류로 분류된다.개별 규조종을 분류하기 위한 몇 가지 시스템이 존재한다.
화석 증거는 규조류가 약 1억 5천만 년에서 2억 년 전이었던 쥐라기 초기 또는 그 이전에 시작되었다는 것을 암시한다.규조류에 대한 가장 오래된 화석 증거는 쥐라기 후기의 현존하는 호박속 표본이다.[22]
규조류는 과거와 현재의 환경 조건을 감시하는 데 사용되며, 수질 연구에 일반적으로 사용됩니다.규조토(규조토)는 지각에서 발견되는 규조 껍데기의 집합체이다.그것들은 부드럽고 실리카가 함유된 퇴적암으로 쉽게 분쇄되어 고운 가루로 만들어지며 일반적으로 10에서 200 μm의 입자 크기를 가지고 있다.규조토는 물여과, 연마재, 고양이 배설물, 다이너마이트 안정제 등 다양한 용도로 사용된다.
형태학
규조류는 일반적으로 크기가 [15]2에서 200마이크로미터이며, 몇 가지 더 큰 종도 있다.그들의 황갈색 엽록체는 광합성의 장소이며, 4개의 세포막을 가지고 있고 카로티노이드 푸코산틴과 같은 색소를 포함하고 있는 전형적인 이색체입니다.개인은 보통 편모가 없지만, 그들은 중심 규조류의 수컷 생식체에 존재하며 다른 그룹의 털(마스티고네임) 특성을 포함한 일반적인 이형체 구조를 가지고 있다.
규조류는 광학적 [26]특성 때문에 종종 "바다의 보석" 또는 "살아 있는 오팔"로 불립니다.이러한 구조적 색채의 생물학적 기능은 명확하지 않지만 통신, 위장, 열 교환 및/또는 자외선 [27]차단과 관련이 있을 수 있습니다.
규조류는 주로 [28]: 25–30 실리카로 구성된 좌절이라고 불리는 복잡하고 단단하지만 다공질 세포벽을 만듭니다.이 규소성[29] 벽은 다양한 모공, 갈비뼈, 미세한 가시, 가장자리 능선 및 고도를 통해 높은 패턴을 만들 수 있습니다. 이 모든 것들은 속과 종을 묘사하는 데 사용될 수 있습니다.
셀 자체는 기본적으로 평평한 플레이트 또는 밸브와 한계 연결 또는 거들 밴드를 포함하는 두 개의 반으로 구성됩니다.반쪽인 빗변은 다른 반쪽인 상피보다 약간 작습니다.규조류의 형태는 다양하다.셀의 모양은 일반적으로 원형이지만 일부 셀은 삼각형, 정사각형 또는 타원형일 수 있습니다.그들의 구별되는 특징은 오팔로 구성된 단단한 미네랄 쉘 또는 좌절입니다.



규조류는 좌골의 모양에 따라 구분되는 두 개의 그룹으로 나뉩니다: 중심 규조류와 페네이트 규조류입니다.
페네이트 규조는 좌우 대칭이다.각각의 밸브에는 랩을 따라 벌어진 틈이 있으며, 일반적으로 랩과 평행하게 껍질이 늘어납니다.그것들은 랩을 따라 흐르는 세포질을 통해 세포의 움직임을 만들어 내고 항상 단단한 표면을 따라 움직입니다.
중심 규조는 반지름 대칭이다.상·하부 밸브(상피 밸브와 하피 밸브)로 구성되어 있으며, 각각 밸브와 거들 밴드로 구성되어 있으며, 밸브는 서로 아래로 쉽게 미끄러지고 확장되어 규조 진행에 따라 세포 함량이 증가합니다.중심 규조체의 세포질은 셸의 내부 표면을 따라 위치하며 세포 중심에 위치한 큰 액포 주위에 중공 라이닝을 제공합니다.이 큰 중앙 액포는 바닷물과 비슷하지만 특정 이온 함량에 따라 변화하는 "세포 수액"으로 알려진 액체에 의해 채워집니다.세포질층은 엽록체와 미토콘드리아와 같은 몇몇 세포소기관들의 집이다.중심 규조류는 팽창하기 전에 그 핵이 밸브의 중심에 있고 분열이 완료되기 전에 세포질층의 중심을 향해 이동하기 시작합니다.중심 규조류는 껍질이 어느 축에서 뻗어 나오느냐와 가시가 존재하느냐에 따라 다양한 모양과 크기를 가지고 있습니다.


행동
대부분의 중심성 및 거미질 페네이트 규조류는 비운동성이며, 상대적으로 밀도가 높은 세포벽으로 인해 쉽게 가라앉습니다.탁 트인 물에서 플랑크톤 형성은 보통 햇빛이 비치는 지표수에서 부유하게 유지하기 위해 바람에 의해 해양수 상층의 난류 혼합에 의존합니다.많은 플랑크톤 규조류들은 또한 가시나 식민지 [33]사슬에서 자라는 능력처럼 가라앉는 속도를 늦추는 특징들을 발전시켜 왔다.이러한 어댑테이션은 표면적 대 체적 비율 및 항력을 증가시켜 물기둥에 더 오래 매달려 있도록 합니다.개별 셀은 이온 [34]펌프를 통해 부력을 조절할 수 있습니다.
일부 페네이트 규조류는 "활공"이라고 불리는 형태의 이동을 할 수 있으며, 이를 통해 라피를 통해 분비되는 접착성 점액질(밸브 [35][36]표면의 가늘고 긴 틈)을 통해 표면을 가로질러 이동할 수 있습니다.규조세포가 활공하기 위해서는 점액질 접착용 고체 기질이 있어야 한다.
세포는 단독 또는 다양한 종류의 집락으로 뭉쳐져 있으며, 이는 규소 구조, 점액 패드, 줄기 또는 튜브, 비정질 점액 덩어리 또는 세포의 스트럿 과정을 통해 분비되는 키틴(다당류)의 실로 연결될 수 있다.
_(20671468900)-47.jpg)
1897년 그란
_(20671468900)-48%2B49.jpg)
_(20671468900)-cropped.jpg)

라이프 사이클


재생산 및 세포 크기
이 유기체들 사이의 번식은 이원분열에 의한 무성분열로 이루어지며, 이 기간 동안 규조류는 두 부분으로 나뉘어 동일한 유전자를 가진 두 개의 "새로운" 규조체를 생성한다.각각의 새로운 유기체는 부모에 의해 소유되는 두 개의 좌절을 받는다 – 하나는 더 크고 다른 하나는 더 작다 – 그것은 현재 에피테카라고 불린다; 그리고 두 번째, 더 작은 좌절을 형성하는데 사용된다.더 큰 좌절을 받은 규조류는 그 부모와 같은 크기가 되지만, 더 작은 좌절을 받은 규조류는 그 부모보다 작은 상태로 유지됩니다.이로 인해 이 규조군의 평균 셀 크기가 [15]감소합니다.그러나 특정 분류군은 세포 [37]크기를 줄이지 않고 분열할 수 있는 능력을 가지고 있는 것으로 관찰되었다.그럼에도 불구하고, 규조 개체군의 세포 크기를 회복하기 위해서는, 성생식과 보조 포자 형성이 [15]일어나야 한다.
세포분열
규조류의 생식세포는 이배체(2N)이며, 따라서 감수분열이 일어날 수 있으며, 수컷과 암컷의 생식세포를 생성하고, 그 생식세포는 접합자를 형성하기 위해 융합한다.접합자는 실리카 테카(silica theca)를 벗겨내고 유기막인 보조 포자로 덮인 큰 구체로 자란다.최대 크기의 새로운 규조 세포, 즉 초기 세포가 보조 포자 내에서 형성되어 새로운 세대를 시작합니다.휴면 포자는 조건이 [28]개선될 때 발아 발생과 함께 불리한 환경 조건에 대한 반응으로 형성될 수도 있다.
정자 운동성
규조류는 대부분 비운동성이지만, 일부 종에서 발견되는 정자는 편모될 수 있지만, 운동성은 보통 활공 [28]운동으로 제한된다.중심 규조류에서, 작은 수컷 배우자는 하나의 편모를 가지고 있는 반면, 암컷 배우자는 크고 운동성이 없다.반대로, 페네이트 디아툼에서는 두 배우자 모두 편모가 [15]없다.라페가 없는 페네이트 규조인 아라피드 종은 비등방성으로 기록되었고, 따라서 중심과 라피드 페네이트 규조, 라페가 [37]있는 규조 사이의 과도기를 나타내는 것으로 여겨집니다.
미생물에 의한 분해
바다와 호수에 있는 특정 종의 박테리아는 유기 조류 [38][39]물질을 분해하기 위해 가수 분해 효소를 사용함으로써 죽은 것이나 살아있는 규조류에서 실리카의 용해 속도를 가속화할 수 있다.
생태학




분배
규조류는 널리 분포하는 집단으로 바다, 민물, 토양, 습한 표면에서 발견될 수 있다.다른 식물성 플랑크톤 [43]그룹보다 더 빨리 분열할 수 있기 때문에 영양분이 풍부한 연안 해역이나 대양 봄 개화기에 식물성 플랑크톤의 주요 성분 중 하나입니다.대부분은 탁 트인 물에서 살지만 일부는 물-침전계면(바닥)에서 표면막으로 살거나 습한 대기 조건에서도 살 수 있습니다.그것들은 유기 물질의 [44]전체 해양 1차 생산의 45%를 차지하는 바다에서 특히 중요하다.해양 식물성 플랑크톤 종의 공간적 분포는 수평과 [45][28]수직 모두 제한된다.
성장
담수 및 해양 환경의 플랑크톤 규조는 전형적으로 "붐 앤 버스트" (또는 "블룸 앤 버스트") 라이프 스타일을 보인다.상부 혼합층(영양분과 빛)의 조건이 양호할 때(봄과 같이) 경쟁력과 빠른 성장률은[43] 식물성 플랑크톤 군집("붐")을 지배할 수 있게 한다.따라서 그들은 종종 기회주의적 r-전략가(즉, 생태가 높은 성장률에 의해 정의된 유기체)로 분류된다.
현대 해양 실리콘 사이클에 대한 기여
규조류는 현대의 해양 실리콘 순환에 중요한 방식으로 기여한다: 규조류는 생물학적 생산의 대부분을 차지하는 원천이다.
영향
흔히 디디모라고 알려진 담수 규조류 디디모스페니아 제미나타는 "갈색 콧물" 또는 "바위 콧물"이라고 불리는 다량의 갈색 젤리 같은 물질을 생산하면서, 그것이 피는 곳의 수로에서 심각한 환경 악화를 일으킨다.이 규조류는 유럽 원산으로 대척지 및 북미 [46][47]일부 지역에 서식하는 침습종이다.이 문제는 호주와 [48]뉴질랜드에서 가장 자주 기록된다.
일반적으로 영양소가 고갈되면 규조세포는 침하 속도가 증가하여 상부 혼합층("버스트")을 빠져나간다.이 침하 현상은 부력 제어의 상실, 규조세포를 서로 붙이는 점액질의 합성 또는 무거운 휴지 포자의 생성에 의해 유발됩니다.상부 혼합층에서 침하하면 방목 인구와 높은 온도(그렇지 않으면 세포 대사를 증가시킬 수 있음)를 포함하여 성장에 불리한 조건에서 규조류가 제거된다.더 깊은 물이나 얕은 해저에 도달한 세포들은 다시 조건이 좋아질 때까지 쉴 수 있다.외양에서, 많은 가라앉는 세포들이 깊은 곳으로 사라지지만, 피난민들은 열전선 근처에서 지속될 수 있다.
궁극적으로, 이러한 휴식 집단의 규조 세포는 수직 혼합이 규조 세포에 들어갈 때 상부 혼합 층으로 재진입한다.대부분의 경우, 이 혼합물은 또한 상부 혼합층의 영양분을 보충하여 다음 단계의 규조 개화를 위한 장면을 준비합니다.외양(계속[49] 융기 지역으로부터 멀리 떨어진 곳)에서, 이러한 개화, 흉작, 그리고 개화 전 상태로 돌아가는 순환은 일반적으로 1년 주기로 일어나며, 규조류는 봄과 초여름에만 널리 퍼져 있다.그러나 일부 지역에서는 여름 성층 파괴와 영양소 혼입으로 인해 가을 개화가 발생할 수 있으며, 빛 수준은 아직 성장에 충분합니다.수직 혼합이 증가하고 겨울이 다가올수록 조도가 낮아지기 때문에 이러한 꽃은 봄의 꽃보다 작고 수명이 짧습니다.
외양에서 규조(봄)의 꽃은 일반적으로 실리콘의 부족으로 끝납니다.다른 미네랄과 달리 실리콘의 요구량은 규조류 특유의 것으로, 예를 들어 질소나 인의 영양소만큼 효율적으로 플랑크톤 생태계에서 재생되지 않습니다.이것은 표면 영양소 농도 지도에서 볼 수 있습니다 – 영양소가 구배를 따라 감소함에 따라 실리콘이 가장 먼저 고갈됩니다(일반적으로 질소와 인이 그 뒤를 따릅니다).
이러한 개화와 불산 주기 때문에, 규조는 해양 지표수로부터[49][50] 탄소를 수출하는 데 불균형적으로 중요한 역할을 하는 것으로 여겨진다(생물 펌프 참조).의미심장하게, 그것들은 또한 현대 [44][40]해양에서 실리콘의 생물 지구 화학적 순환을 조절하는 데 중요한 역할을 한다.
성공 이유
규조는 생태학적으로 성공적이며, 해양, 바다, 호수, 개울뿐만 아니라 토양과 [citation needed]습지 등 물을 포함한 거의 모든 환경에서 발생합니다.규소에 의한 규소의 사용은 많은 연구자들에 의해 이러한 생태학적 성공의 열쇠로 여겨진다.Raven([51]1983)은 유기 세포벽에 비해 실리카 좌절을 합성하는 데 더 적은 에너지(비교적 유기벽의 약 8%)를 필요로 하므로 전체 세포 에너지 예산을 상당히 절약할 수 있다고 지적했다.지금은 고전적인 연구에서, Egge와 Aksnes([42]1992)는 메소코스름 군집의 규조 우위가 규산의 가용성과 직접 관련이 있다는 것을 발견했다. 농도가 2μmol−3 m 이상일 때 규조류가 전형적으로 식물 플랑크톤 군집의 70% 이상을 차지한다는 것을 발견했다.다른 연구자들은[52] 규조 세포벽에 있는 생물성 실리카가 효과적인 pH 완충제로 작용하여 중탄산염을 용해된2 CO로 전환하는 것을 용이하게 한다고 제안했다.일반적으로 규소의 사용으로 얻을 수 있는 이러한 이점에도 불구하고 규조류는 일반적으로 동일한 크기의 [43]다른 조류보다 높은 성장률을 보입니다.
수집 소스
규조는 여러 출처에서 [53]얻을 수 있습니다.해양 규조류는 직접 물채취로 채취할 수 있으며, 해저 형태는 따개비, 굴, 기타 조개껍데기를 긁어내는 방법으로 확보할 수 있다.규조류는 물에 잠긴 돌과 막대기에 갈색으로 미끄러운 코팅으로 자주 나타나며, 강물과 함께 "흐름"으로 보일 수 있습니다.연못, 도랑, 석호의 수면 진흙은 거의 항상 약간의 규조들을 만들어낸다.살아있는 규조류는 종종 필라멘트 조류에 대량으로 달라붙거나 물에 잠긴 다양한 식물에 젤라틴 덩어리를 형성하는 것으로 발견됩니다.Cladophora는 종종 타원형 규조인 Coconeis로 덮여있고, Vaucheria는 종종 작은 형태로 덮여있다.규조류는 연체동물, 참치류, 어류의 먹이의 중요한 부분을 형성하기 때문에, 이러한 동물의 소화관은 종종 다른 방법으로는 쉽게 확보되지 않는 형태를 만들어낸다.항아리에 물과 진흙을 채우고 검은 종이로 싸서 직사광선을 수면 위로 내리쬐면 규조류가 생겨날 수 있다.하루 안에, 그 규조들은 쓰레기 더미를 타고 꼭대기로 올라올 것이고,[53] 고립될 수 있다.
생화학

| 시리즈의 일부 |
| 플랑크톤 |
|---|
 |
에너지원
규조류는 주로 광합성을 하지만 일부는 필수 이종영양동물이고 적절한 유기탄소원이 [54][55]있다면 빛이 없어도 살 수 있다.
실리카 대사
규조 세포는 전형적으로 서로 [56]겹치는 두 개의 밸브로 이루어진 좌절로 알려진 독특한 실리카 세포벽 안에 포함되어 있습니다.세포벽을 구성하는 생물학적 실리카는 규산 단량체의 중합에 의해 세포 내에서 합성된다.그런 다음 이 재료를 셀 외부로 압출하여 벽에 추가합니다.대부분의 종에서, 규조류가 두 개의 딸 세포를 만들기 위해 분열할 때, 각각의 세포는 두 개의 하프 중 하나를 유지하고 그 안에서 더 작은 반으로 자란다.그 결과, 각 분할 주기 후에, 모집단의 규조세포의 평균 크기는 작아집니다.일단 그러한 세포들이 단순히 분열하는 것이 아니라 특정한 최소 크기에 도달하면, 보조 포자를 형성함으로써 이러한 감소세를 반전시킨다.이 경우 크기가 확장되어 훨씬 더 큰 셀이 생성되며, 셀은 크기를 줄이는 [citation needed]분할로 돌아갑니다.보조 포자의 생산은 거의 항상 감수분열과 성적 번식과 관련이 있다.
규조류에 흡수된 실리카를 세포벽에 전달하는 정확한 메커니즘은 알려지지 않았다.규조 유전자의 배열의 대부분은 실리카 흡수와 나노 스케일 패턴의 퇴적 메커니즘에 대한 탐색에서 비롯됩니다.이 분야에서 가장 큰 성공은 전체 게놈의 염기서열과 유전자 제어 방법이 확립되면서 모델종이 된 탈라시오시라 슈도나나와 중요한 실리카 퇴적 단백질 실라핀이 [57]처음 발견된 원통형 방추형질 두 종에서 비롯됐다.폴리카티온성 펩타이드 세트인 실라핀은 방추상 세포벽에서 발견되었으며 복잡한 실리카 구조를 생성할 수 있다.이러한 구조는 규조 패턴에 특유한 크기의 모공을 보여주었습니다.T. pseudonana는 게놈 분석을 받았을 때 대부분의 게놈보다 더 많은 수의 폴리아민뿐만 아니라 세 개의 서로 다른 실리카 [58]전달 유전자를 포함한 요소 회로를 암호화한 것으로 밝혀졌다.규조류 8개 그룹의 실리카 운반 유전자에 대한 계통발생학적 연구에서 실리카 운반은 일반적으로 [57]종과 그룹화하는 것으로 밝혀졌다.이 연구는 또한 페네이트(양쪽 대칭)와 중심(반경 대칭) 규소의 실리카 운반체 사이의 구조적 차이를 발견했다.본 연구에서 비교한 시퀀스는 실리카 증착 과정에서 기능을 구별하는 잔류물을 식별하기 위해 다양한 배경을 만드는 데 사용되었다.또한, 같은 연구에서 많은 지역들이 실리카 수송의 기초 구조인 종 내에서 보존된 것으로 밝혀졌다.
이 실리카 수송 단백질은 규조류에서만 볼 수 있으며, 스폰지나 쌀과 같은 다른 종에서는 상동체가 발견되지 않습니다.이러한 실리카 수송 유전자의 분산은 또한 유전자 복제 또는 이합체화를 [57]나타내는 5개의 막 결합 세그먼트로 구성된 두 개의 반복 단위에서 진화하는 단백질의 구조를 나타낸다.규조체의 막 결합 소포에서 발생하는 실리카 퇴적은 실라핀과 긴 사슬 폴리아민의 활성의 결과로 가설화되었습니다.이 실리카 증착 베시클([59]SDV)은 골지 유래 베시클과 융합된 산성 구획으로 특징지어졌습니다.이 두 가지 단백질 구조는 규조체 좌절을 기준으로 불규칙한 모공을 가진 패턴화된 실리카 인비보 시트를 생성하는 것으로 나타났다.이러한 단백질이 복잡한 구조를 만들기 위해 어떻게 작용하는지에 대한 한 가지 가설은 SDV 내에 잔류물이 보존된다는 것인데, 이용 가능한 다양한 배열의 수가 제한되어 있기 때문에 불행히도 식별하거나 관찰하기가 어렵다.실리카의 매우 균일한 증착의 정확한 메커니즘은 아직 알려지지 않았지만, 실라핀과 연결된 탈라시오시라 유사나나 유전자는 나노 크기의 실리카 증착의 유전자 제어 대상으로 여겨지고 있다.
요소 순환
규조류의 특징은 요소 순환인데, 이것은 그것들을 동물과 진화적으로 연결시켜준다.이것은 Andrew Allen, Chris Bowler, 그리고 동료들에 의해 수행된 연구에서 발견되었습니다.2011년에 발표된 그들의 연구결과는 규조들이 기능하는 요소 사이클을 가지고 있다는 것이 매우 중요했다. 왜냐하면, 그 이전에는 요소 사이클이 규조보다 수억 년 전에 출현한 메타조안으로부터 유래한 것으로 생각되었기 때문이다.그들의 연구는 규조류와 동물이 요소 순환을 다른 목적에 사용하는 반면, 그들은 동물과 식물들이 [60]그렇지 않은 방식으로 진화적으로 연결되어 있는 것으로 보여진다는 것을 보여주었다.
색소
디아톡스의 주요 색소는 클로로필 a 및 c, 베타카로틴, 푸코산틴,[15] 디아톡산틴 및 디아디노산틴이다.
스토리지 제품
분류법

.jpg)


스테파노디스커스한츠치이
.jpg)
신경성 지협신경성 지협

오돈텔라오리타
규조류는 클로로필 a와 c가 풍부한 플라스티드를 많이 포함하고 있는 원생생물들의 큰 그룹에 속합니다.이 집단은 이종혼합체, 번데기, 크로미스트 또는 스트라메노파일로 다양하게 언급되어 왔다.금조류나 다시마와 같은 자가영양동물과 물 곰팡이, 오팔리니드, 방선동물과 같은 이종영양동물이다.이 지역의 시위자 분류는 아직 미정입니다.계급에 있어서, 그들은 분류, 문, 왕국, 또는 그것들의 중간적인 것으로 취급되어 왔다.결과적으로, 규조류는 보통 규조류 또는 바실라리오피케아라고 불리는 분류에서 보통 바실라리오피타라고 불리는 분류(=문)까지 분류되며, 이에 상응하는 하위 그룹의 서열 변화가 있다.
속과 종
2012년 Guiry에[61] 따르면 현존하는 규조류는 약 20,000종으로 추정되며, 그 중 약 12,000종이 현재까지 명명되었다(다른 출처에서는 더 광범위한 추정치를[15][62][63][64] 제공한다).약 1,000-1,300개의 규조류가 현존 및 [65][66]화석으로 기술되어 있으며, 그 중 250-300개는 [67]화석으로만 존재한다.
클래스 및 순서
여러 해 동안 규조류는 분류(Bacillariophyae) 또는 문(Bacillariophyta)으로 취급되어 중심 규조류와 페나트 규조류(Centrales and Pennales)에 해당하는 단 2개의 목으로 구분되었다.이 분류는 1990년 Round, Crawford 및 Mann에 의해 광범위하게 정비되었으며, 그는 규조류를 더 높은 등급(동물 분류에서 문과에 해당)으로 취급하고, 주요 분류 단위를 분류로 승격시켜 중심 규조류를 단일 등급으로 유지하되, 이전의 페네이트를 쪼개었다.규조류는 두 개의 분리된 분류로 나뉘는데, Fragilariophyceae와 Bacillariophyceae(후자의 이름은 유지되지만 수정된 정의를 가지고 있음)로 나뉘며, 이들 사이에는 45개의 목(대부분 새로운 목)이 포함되어 있다.
현재(2020년 중반 집필) Round 등의 1990년 시스템은 새로운 분자 연구의 출현과 함께 수정이 필요하지만, 이를 대체할 최선의 시스템은 불분명하며, AlgeBase, World Register of Marine Base 및 그 기여 데이터베이스 Diatom Base와 같은 널리 사용되는 현재 시스템은 불명확하다.2015년 루지에로 외 연구진(Ruggiero et al., 2015)에 대표되는 "모든 생명"은 계통발생학적 그룹 및 인접한 그룹의 처리와 더 잘 일치하기 위해 분류/문이 아닌 전체 분류로 취급되고 라운드 외 연구진의 분류가 하위 분류로 분류되었지만, 모두 라운드 외 치료법을 기초로 유지한다(참조).아래의 각 섹션을 참조해 주세요.
2004년에 시작된 린다 메들린과 동료들의 제안 중 하나는 페네이트와 더 밀접하게 관련된 것으로 간주되는 중심 규조목의 일부를 새로운 등급인 메디오파이스에로 분할하는 것입니다.메디오파이스에는 페네이트 규조 자체가 나머지 중심보다 페네이트 규조류와 더 밀접하게 일치합니다.나중에 Coscinodiscophyceae-Mediophyceae-Bacillariophyceae 또는 CMB(Mediophyceae+Bacillariophyceae) 가설로 명명된 이 가설은 D.G Mann에 의해 다른 가설로 받아들여졌다.Archibald 등이 편집한 2017년 원생 핸드북의 Bacillariophyta 장에 있는 일련의 합성(2005년, 2012년, 2019년)과 Medlin 등 원본 "Coscinodiscophyceae"의 명백한 비일관성을 반영한 수정이 있다.한편, E.C.가 이끄는 그룹.Theriot은 구조 계조 가설(SGH)로 불리며 Mediophyceae를 단통군으로 인정하지 않는 다른 계통 발생 가설을 선호한다. Parks 등, 2018년의 분석에서는 반지름 중심 규조류가 단통군이 아닌 것으로 밝혀졌다(Medlin 등의 Coscinodophyae).변칙속인 메디오피케아에서 아테야까지의 단생동물.이러한 상충되는 계획의 상대적 장점에 대한 논의는 관련 [68][69][70][71]당사자들에 의해 계속된다.
Adl et al., 2019 치료
2019년에 Adl [72]등은 다음과 같은 규조류 분류를 제시하면서 "이 개정안은 지난 10년간 규조류의 계통 발생의 수많은 발전을 반영한다.메디오파이스와 페나테 규조류 이외의 분류군의 표본 추출이 불충분하고 모든 규조류가 알려져 있고 예상되는 다양성 때문에 많은 분류군이 높은 분류 수준으로 나타납니다(그리고 높은 분류는 오히려 평평합니다).이 분류는 규조류를 문(Diatomeae/Bacillariophyta)으로 취급하고, 메들린과 동료의 분류를 받아들이고, 다른 방법으로 분리된 여러 속들에 대한 새로운 아종과 분류를 도입하며, 이전에 확립된 분류군을 아종으로 분류하지만, 목이나 과는 나열하지 않는다.추정한 순위는 명확성을 위해 추가되었다(Adl. 등은 순위를 사용하지 않지만, 분류의 이 부분에서 의도한 순위는 사용된 식물 명명법 체계 내에서 사용된 어미의 선택에서 명백하다).
- Clade Diatomista Derelle et al. 2016, 수정.Cavalier-Smith 2017(기타 황화식물군의 하위 집합과 디아톰)
- Diatomeae Dumortier 1821 [= Bacillariophta Haeckel 1878] (diatoms)
- Adl et al. 2019 렙토실린드로피티나D.G.만아문
- Adl 등 2019년 렙토실린드로피체아 D.G. 만 분류(렙토실린드루스, 테누이실린드루스)
- Adl et al. 2019(Corethrophyceae D.G. Mann) 분류
- 아들 외 연구진(Ellerbeckiophytina D.G. Mann) 아문(Ellerbeckia)
- Adl et al. 2019 Probosciophytina D.G. Mann 아문(Proboscia)
- Adl et al. 2019 Melosirophytina D.G. Mann 아문(Aulacosira, Melosira, Hyalodiscus, Stephanopyxis, Paralia, Endictya)
- 아문 Coscinodiscophylam Medlin & Kaczmarska 2004, emend. (Actinoptychus, Coscinodiscus, Actinocyclus, Aulacodiscus, Stellerima)
- 아드르 외 연구진(Guinardia, Rhizosolenia, Pseudosolenia)의 Rhizosoleniophytina D.G. Mann 아문
- 아문(Arachnoidisctina D.G. Mann) 아문(Arachnoidiscus)
- 바실라리오피티나 메들린과 카츠마르스카 2004 아문, 수정.
- Class Mediophyceae Jouse & Proshkina-Lavrenko in Medlin & Kaczmarska 2004
- Round et al. 1990년 Chaetocerotophycidae Round & R.M. Crawford 하위 분류, 수정.
- Round et al. 1990년 아강 Lithodesmiophycidae Round & R.M. Crawford, 수정.
- Round et al. 1990년 Thalassiosirophycidae Round & R.M. 크로포드 아강
- Round 등 1990년 Round & Round & R.M. Crawford
- 아드르 외 연구진의 아강 오돈텔로피쿠스과 D.G. 만
- 아강 국화과 D.G. 만(Adl et al. 2019)
- Adl et al. 2019년 비둘피오피오파이스아과 D.G. 만
- Biddulphiophycidae Round와 R.M. Crawford의 하위 분류, 1990년 Round 등.
- 비둘피오파시카에인서테세디스(Atheya)
- 1878년 학번 바실라리오피셰이 해켈 1878년, 수정.
- 박실라리오피케아 인테르테아 세디스(Striatellaceae)
- 메들린아강(Urneidophycidae
- 1990년 라운드, 크로포드, 맨의 아강 Fragilariophycidae 라운드, 수정.
- Round, Crawford, Mann 1990의 Bacillariophycidae D.G. Mann 아강, 수정.
자세한 내용은 규조 분류법을 참조하십시오.
갤러리
- 스캔 전자 현미경 이미지

스리렐라 나선형 규조개

막 필터에 디아톰스 탈라시오시라 스패, 모공 크기 0.4μm.

디아톰 패럴리아 설카타.

아칸테스 트리노디스





바실라리아 팍실리퍼의 독립 세포
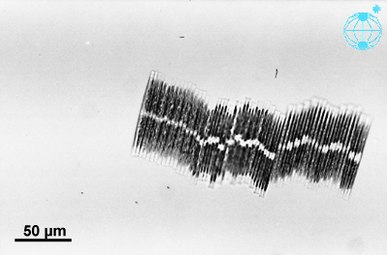
바실라리아 팍실리퍼 군락군


| 외부 비디오 | |
|---|---|
남극 대륙의 거대한 규조류와 독점적인 식민지 규조인 바실라리아 패러독사 등 3종의 규조류가 국제우주정거장으로 보내졌다.바실라리아의 세포들은 미세유체학법에 [73]의해 부분적이지만 반대되는 동기화로 서로 옆으로 이동했다.
진화 및 화석 기록
기원.
헤테로콘트 엽록체는 식물에서 발생한 원핵생물에서 직접 유래한 것이 아니라 홍조류 엽록체에서 유래한 것으로 보인다.이것은 그들이 다른 많은 해조류보다 더 최근의 기원을 가졌다는 것을 암시한다.그러나, 화석의 증거는 거의 없고, 규조 자체의 진화에 의해서만 이종 혼혈동물이 화석 기록에 심각한 인상을 남긴다.
초기 화석
최초의 화석[74] 규조류는[75] 분자 시계와 퇴적물 증거로 더 이른 기원을 시사하지만, 초기 쥐라기(약 185 Ma 전)[74]부터 시작되었다.이들의 기원은 많은 해양 틈새들이 [76]열리게 된 페르미아의 대멸종(약 250Ma)과 관련이 있을 것으로 추측되고 있다.이 사건과 화석 규조류가 처음 출현한 시기 사이의 차이는 규조류가 비규산화되고 그 진화가 [77]불가사의했던 시기를 나타낼 수 있다.규화의 도래 이후 규조류는 백악기 초기까지 발견된 주요 화석 퇴적물과 규조토와 같은 일부 암석들이 거의 전부 규조류로 이루어져 화석 기록에 큰 인상을 남겼다.
실리콘 사이클과의 관계
규조류가 트라이아스기 이후 존재했을지 모르지만, 규조류가 득세한 시기와 실리콘 사이클의 "인수"는 더 최근에 일어났다.Phanerozeoic 이전(544 Ma 이전)에는 미생물이나 무기 과정이 바다의 실리콘 [78][79][80]주기를 약하게 조절했다고 믿어졌다.그 후, 주기는 방사선과와 규소성 스펀지에 의해 지배되고(그리고 더 강력하게 규제된다), 전자는 동물성 플랑크톤으로, 후자는 주로 [81]대륙붕에서 좌식 필터 피더로 나타난다.최근 100My 이내에 실리콘 사이클은 더욱 엄격한 제어 하에 있으며, 이는 규조류의 생태학적 우위에 기인한다고 생각된다.
그러나 정확한 인수 시기는 불분명하고 화석 기록에 대한 저자들의 해석도 엇갈리고 있다.석회질 스폰지의 [82]선반으로부터의 이동과 같은 몇몇 증거들은 이러한 점령이 백악기 (146 Ma에서 66 Ma)에 시작되었음을 시사하는 반면, 방사선과 의사들의 증거는 "인수"가 신생대 (현재 [83]66 Ma)까지 시작되지 않았음을 시사한다.
초원과의 관계
마이오세 동안 초원 생물군의 팽창과 풀의 진화적 방사선은 바다에 대한 용해성 실리콘의 플럭스를 증가시켰다고 여겨지며, 이것이 신생대에 [84][85]규조류를 촉진시켰다는 주장이 제기되어 왔다.최근의 연구는 규조류와 초원의 다양성 모두 마이오세 [86]중기에 비해 크게 증가했지만 규조류의 성공이 풀의 진화로부터 분리되었음을 시사한다.
기후와의 관계
신생대 위의 규조 다양성은 지구 온도, 특히 적도-극 온도 구배에 매우 민감합니다.과거 따뜻한 바다, 특히 따뜻한 극지방은 규조 다양성이 상당히 낮았던 것으로 나타났다.지구 온난화 [87]시나리오에서 예측된 바와 같이, 극지 온난화가 강화된 미래의 따뜻한 바다는 이론적으로 규조 다양성의 상당한 손실을 초래할 수 있지만, 현재의 지식으로는 이것이 빠르게 일어날 것인지 수만 [86]년 이상에 걸쳐서만 일어날 것인지 말할 수 없다.
조사방법
규조류의 화석 기록은 주로 해양 퇴적물과 비해양 퇴적물에서의 규조성 좌절을 회복함으로써 확립되었다.규조류는 해양과 해양이 아닌 지층학 기록을 가지고 있지만, 시간에 제약된 진화의 기원과 독특한 분류군의 멸종에 바탕을 둔 규조 생물층학은 해양 시스템에만 잘 발달되어 널리 적용되고 있다.규조류의 지속 시간은 [88]육지에 노출된 해양핵과 암석 염기서열 연구를 통해 입증되었다.규조 바이오존이 지자기 극성 시간 척도에 따라 잘 확립되고 보정되는 경우(예: 남양, 북태평양, 동부 적도 태평양), 규조 기반 연령 추정치는 신생대 규조 조립의 일반적인 연령 분해능은 수십만 년이지만 10만 년 이내로 분해될 수 있다.
호수 퇴적물에 보존된 규조류는 4차 기후의 고환경적 재건, 특히 수심과 염도의 변동을 경험하는 폐쇄 유역 호수에 널리 사용된다.
동위원소 기록

규조류가 죽으면 그들의 껍데기는 해저에 가라앉아 미세 화석이 될 수 있다.시간이 지남에 따라, 이 미세 화석은 해양 퇴적물에 오팔 퇴적물로 묻히게 된다.고기후학은 과거의 기후에 대한 연구이다.프록시 데이터는 현대의 퇴적물 샘플에서 수집된 요소를 과거의 기후 및 해양 조건과 관련짓기 위해 사용된다.고엽산염 프록시란 직접적인 기상 또는 해양 [89]측정의 대체 역할을 하는 보존되거나 화석화된 물리적 표지를 말한다.프록시의 예로는 규조 동위원소 기록의 13C, 18O, 30Si(13Cdiatom, 18Odiatom, 30Sidiatom 이상)가 있다.2015년 스완과 스넬링은 이러한 동위원소 기록을 사용하여 영양소 공급과 연조직 생물 펌프의 효율성을 포함한 북서태평양의 광구 조건의 과거 변화를 현대에서 마지막 간빙기와 일치하는 해양 동위원소 단계 5e까지 문서화했다.해양 동위원소 단계에서 오팔 생산성의 피크는 지역 할로카인 층화의 파괴와 광대에 [90]대한 영양소 공급 증가와 관련이 있다.


할로크라인과 성층수 기둥의 초기 발달은 2.73Ma에서 주요 북반구 빙하가 시작되면서 몬순 강우 및/또는 빙하 용융수 증가, 해수면 [91][92][93][94]온도 상승으로 인해 이 지역에 대한 담수의 유속이 증가했기 때문이다.이 염분 약층이 늦어서 플리 오세 그리고 일찍 사차glacial–interglacial cycles,[95] 다른 studie을 통해 승리를 거둔 것으로 보인다 심해 수 upwelling 이것과 관련된 수의 감소 전 세계적으로 시원한 조건의 신설과 빙하의 북반구를 가로질러 2.73Ma.[92]로 확대에 기여했다.s이다성층 경계가 4차 말기 빙하 종말과 간빙하 [96][97][98][99][100][90]초기에 무너졌을 수 있음을 보여준다.
다양화
백악기의 규조류 기록은 제한적이지만, 최근의 연구에 따르면 규조류의 종류가 점차 다양해지고 있다.바다에서 석회질 골격을 가진 유기체에 극적으로 영향을 미쳤던 백악기-팔레오진 멸종 사건은 규조 [101]진화에 비교적 적은 영향을 미친 것으로 보인다.
이직률.
신생대 동안 해양 규조류의 대멸종은 관찰되지 않았지만, 해양 규조종 집합에서 비교적 빠른 진화 전환 시기는 고생세-에오세 [102]경계 부근과 에오세-올리고세 [103]경계 부근에서 발생했다.극지방의 점진적인 냉각과 보다 풍토적인 규조 조립의 발달에 대응하여, 조립품의 추가적인 교체가 Miosene [104]중기와 Pliocene 사이에 여러 시기에 일어났다.
올리고세부터 제4기에 걸쳐 보다 섬세한 규조류(diatom respultules)[88]에 대한 세계적인 추세가 나타났다.이는 남극 대륙의 주요 빙상 팽창이 시작될 때 위도 열 구배를 증가시키고 네오겐과 사분기를 통해 양극성 빙하 세계를 향해 점차적으로 냉각함으로써 초래된 해양 표면과 심해의 순환이 점점 활발해지는 것과 일치한다.이것은 규조들이 좌절을 형성하기 위해 더 적은 실리카를 섭취하도록 만들었다.해양의 혼합 증가는 특히 해안과 해양이 융기하는 지역에서 지표수의 규조 생장에 필요한 실리카와 다른 영양소를 재생한다.
유전학
표현된 시퀀스 태그 지정
2002년, 1,000개의 발현 배열 태그(EST)[105]를 사용하여 파에오닥탐문 삼각류 유전자 레퍼토리의 특성에 대한 최초의 통찰이 설명되었다.그 후 EST의 수는 12,000개로 확대되었고 기능 [106]분석을 위해 규조 EST 데이터베이스가 구축되었다.이러한 배열은 녹색 조류 Chlamydomonas reinhardtii, 적색 조류 Cyanidioszon merolae 및 규조류 Thalassiosira pseudonana의 [107]추정 완전 단백질과 P. tricornutum 간의 비교 분석을 위해 사용되었습니다.규조 EST 데이터베이스는 현재 다양한 조건에서 자란 P. tricornutum(16개의 라이브러리) 및 T. pseudonana(7개의 라이브러리) 세포로부터의 200,000개 이상의 EST로 구성되어 있으며, 이들 중 다수는 서로 다른 비생물학적 [108]스트레스에 대응한다.
게놈 배열 분석
2004년에는 중심 규조류인 탈라시오시라 의사나나(32.4Mb)의 전체 게놈 [109]배열을 분석했으며, 2008년에는 페네이트 규조류인 파에오닥탐 트리코르누툼(27.4Mb)[110]의 배열을 분석하였다.두 게놈을 비교한 결과, P. tricornutum 게놈은 T. pseudonana보다 적은 유전자(1,776개 대비 10,402개)를 포함하고 있으며, 두 게놈 사이의 주요 합성(유전자 순서)는 검출되지 않았다.T. pseudonana 유전자는 P. tricornutum의 0.79에 반해 유전자당 평균 1.52개의 인트론을 나타내며, 이는 중심 [110][111]규조체에서 최근 널리 퍼진 인트론 증가를 시사한다.비교적 최근의 진화적 발산(9천만 년)에도 불구하고, 중심체와 페네이트 사이의 분자적 발산 정도는 다른 진핵생물 [110]그룹에 비해 바실라리오피스아 내에서의 빠른 진화 속도를 나타낸다.비교게노믹스는 또한 특정 종류의 트랜스포저블 요소인 Diatom Copia 유사 역트랜스포존(또는 CoD)이 T. pseudonana와 관련하여 P. tricornutum 게놈에서 유의하게 증폭되어 각각의 [112]게놈의 5.8과 1%를 구성한다는 것을 확인했다.
내공생체유전자전달
규조 유전체학은 내공생체 유전자 전달 과정의 범위와 역학에 대한 많은 정보를 가져왔다.다른 유기체의 T. pseudonana 단백질과 호몰로지를 비교한 결과, 수백 개가 플랜태 계통에서 가장 가까운 호몰로지를 가지고 있는 것으로 나타났습니다.규조 게놈에 대한 EGT는 T. pseudonana 게놈이 Gillardia theta(크립토모나드) 핵형 게놈에 의해 코드된 유전자와 가장 가까운 6개의 단백질을 코드한다는 사실로 설명될 수 있다.이들 유전자 중 4개는 또한 적색 조류 플라스티드 게놈에서 발견되며, 따라서 적색 조류 플라스티드에서 적색 조류 핵(핵형질)에서 이종 조류 숙주 [109]핵까지 연속적인 EGT를 나타낸다.규조 프로테옴의 보다 최근의 계통학적 분석은 식물 유래 규조 유전자의 70%가 녹색 계통의 기원이고 그러한 유전자가 다른 지층의 게놈에서도 발견된다는 사실로 뒷받침되는 크롬베올레이트의 공통 조상에서 플라시노피테와 같은 내심비온트에 대한 증거를 제공했다.따라서, 크롬알베올레이트는 먼저 녹조와의 연속적인 2차 내심증 산물이며, 다음으로 앞의 유전체 발자국을 보존하지만 녹색 플라스티드를 [113]대체한 홍조와의 연속적인 2차 내심증 산물이라는 것이 제안되었다.그러나 규조단백질과 크롬알베올레이트 진화역사의 계통유전학 분석은 홍조류 등 염기서열이 낮은 계통으로부터 상보적인 게놈 데이터를 이용할 가능성이 높다.
수평 유전자 이동
EGT와 더불어 수평 유전자 이동(HGT)은 내심균 이벤트와는 독립적으로 발생할 수 있다.P. tricornutum 게놈의 출판물은 적어도 587 P. tricornutum 유전자가 박테리아 유전자와 가장 밀접한 관련이 있는 것으로 보이며, P. tricornutum proteome의 5% 이상을 차지한다고 보고했다.이들 중 절반 가량은 T. pseudonana 게놈에서도 발견되며, 규조류 [110]혈통에 고대인이 혼재했음을 증명한다.
유전공학
지구화학적 순환에서 규조류의 큰 중요성을 뒷받침하는 생물학적 메커니즘을 이해하기 위해 과학자들은 [114]90년대부터 파에오닥탐문 트리코르누툼과 살라시오시라 종을 모델 유기체로 사용해 왔다.몇 분자 생물학 도구 현재 돌연변이 또는 이식 유전자 라인을 생성하는 데:인자 transgenes을 포함하는 세포는biolistic method[115]또는 transkingdom 세균 conjugation[116](10−6과 10−4 수익률 respectively[115][116]과), 그리고 전기 천공 법 또는 사용과 같은 다른 고전적인 형질 주입 방법을 사용하여에 삽입될 수 있다.기뇌 촬영 중repor 왔다.ted는 효율이 [116]낮은 결과를 제공합니다.
트랜스감염된 플라스미드는 (CEN6-ARSH4-HIS3 효모 중심색체 배열[116] 덕분에) 규조체의 염색체에 무작위로 통합되거나 안정적인 원형 에피솜으로 유지될 수 있다.플루오마이신/제오신 내성 유전자 Sh Ble은 선택 [114][117]마커로서 일반적으로 사용되며,[116][117] 다양한 트랜스젠이 성공적으로 도입되어 세대를 통해 안정적인 전달 또는 [117]제거 가능성과 함께 디아톰에서 발현되고 있다.
게다가 이러한 시스템은 이제 CRISPR-Cas 게놈 에디션 도구를 사용할 수 있게 되어 기능적인 녹아웃 돌연변이를[117][118] 빠르게 생산하고 규조류의 세포 과정을 보다 정확하게 이해할 수 있게 되었습니다.
인간의 용도
고생물학
규조류의 분해와 부패는 유기 및 무기(규산염의 형태) 침전물로 이어지며, 무기 성분은 점토와 실트의 퇴적물에 포함되어 영구적인 지질학적 기록을 형성하기 때문에 해양 바닥이나 만 진흙의 코어링에 의해 과거의 해양 환경을 분석하는 방법으로 이어질 수 있습니다.그러한 해양 지층(실리액 분비물 참조).
산업의

규조 및 규조토와 같은 껍데기(과립)는 미세 연마 및 액체 여과에 사용되는 중요한 산업 자원입니다.나노기술의 [119]재료로서 그들의 현미경 껍데기의 복잡한 구조가 제안되어 왔다.
규조석은 천연 나노 물질로 간주되며 다양한 세라믹 제품, 건설 세라믹, 내화 세라믹, 특수 산화물 세라믹스의 생산, 여과재로 사용되는 습도 조절 재료, 시멘트 생산 산업에서 사용되는 재료, 생산용 초기 재료 등 다양한 용도와 용도가 있다.prolonged-release 약 통신사의 Uction 산업 규모에 흡수 물질이 도자기, 유리 산업 촉매요 지지하는 것 플라스틱과 페인트, 산업 물, 살충제 보유자의 정화에 필러뿐만 아니라 특정 토양의 물리적, 화학적 특징을 개선시키는 데에 쓰이고, 다른 용도의 생산.[120][121][122]
규조류는 또한 바닷물을 포함한 규조류를 함유하는 물질의 출처를 알아내는 데에도 사용된다.
법의학
법의학에서 규조 분석의 주요 목표는 물에 몸을 담그는 사체 침수와 침수에 의한 죽음을 구별하는 것이다.실험실 검사를 통해 체내에 규조류가 있는지 알 수 있다.규조류의 실리카 기반 골격은 쉽게 부패하지 않기 때문에 심하게 분해된 물체에서도 검출될 수 있다.체내에서 자연적으로 발생하는 것이 아니기 때문에, 실험실 검사에서 시신이 수습된 물에서 발견된 것과 같은 종류의 규조류가 발견되면, 사망의 원인으로 익사했다는 좋은 증거가 될 수 있다.시신에서 발견된 규조류의 혼합물은 주변 물과 같거나 다를 수 있어 시신이 [123]발견된 장소와 같은 장소에서 익사했는지 여부를 알 수 있다.
나노테크놀로지
규조류에 의한 실리카의 퇴적 또한 나노기술에 [124]유용한 것으로 입증될 수 있다.규조세포는 다양한 형태와 크기의 밸브를 반복적이고 안정적으로 제조하여 규조세포가 광학계, 반도체 나노 리소그래피, 심지어 약물 전달용 차량 등 다양한 장치에서 사용될 수 있는 마이크로 또는 나노 스케일 구조를 잠재적으로 제조할 수 있도록 합니다.적절한 인위적 선택 절차를 통해 특정 형태와 크기의 밸브를 생산하는 규조류를 체모스타트 배양에서 배양하여 나노크기의 [125]성분을 대량 생산하도록 진화시킬 수 있다.규조류가 보통 [126]세포벽을 만들기 위해 사용하는 이산화규소 대신 감광성 이산화티타늄을 대체함으로써 규조류가 태양전지의 구성요소로 사용될 수 있다는 제안도 제기되었다.규조 바이오 연료 생산 태양 전지판도 [127]제안되었다.
발견 이력
규조류의 첫 삽화는 1703년 영국 왕립학회의 거래에서 타벨라리아의 [128]확실한 그림을 보여주는 기사에서 찾을 수 있다.비록 그 출판물은 이름이 알려지지 않은 영국 신사에 의해 쓰여졌지만, 그가 스태퍼드셔의 [128][129]찰스 킹이었다는 증거가 최근 있다.우리가 공식적으로 확인된 최초의 규조인 식민지 바실라리아 팍실리페라를 발견한 것은 불과 80년이 지난 1783년 덴마크 박물학자 오토 프리드리히 [128]뮐러에 의해 발견되고 기술되었다.그의 뒤를 잇는 다른 많은 사람들처럼, 그는 그것이 움직이는 능력 때문에 동물이라고 잘못 생각했다.찰스 다윈조차도 카보베르데 제도에서 규조류가 먼지 속에 남아 있는 것을 보았지만, 그는 그것이 무엇인지 확신하지 못했다.그들은 나중에야 그에게 규소성 다성체로 판명되었다.다윈이 나중에 남아메리카 남단 티에라 델 푸에고 원주민인 푸에기노스의 페이스 페인트에서 주목한 인푸소리아는 나중에 같은 방식으로 확인되었다.그의 생전에, 규소성 다성체는 규조류에 속하는 것으로 밝혀졌고 다윈은 그 아름다움을 뒷받침하는 이유를 이해하려고 애썼다.그는 저명한 암호학자 G. H. K.와 의견을 교환했다.그 주제에 대해 생각해 봅시다.'종의 기원' 제4판에서 그는 "규조과의 미세한 규소성 케이스보다 아름다운 물체는 거의 없다: 이것들은 현미경의 높은 힘 하에서 검사되고 감탄될 수 있도록 창조되었는가?"라고 언급했고, 그들의 정교한 형태학이 가진 것보다 기능적인 기초를 가지고 있어야 한다고 추론했다.g는 순전히 인간이 [130]존경하기 위해 만들어졌다.
「 」를 참조해 주세요.
메모들
레퍼런스
- ^ Dangeard, P. (1933)트라이트 달골로지Paul Lechvalier and Fils, Paris, [1] 2015년 10월 4일 Wayback Machine에서 아카이브 완료.
- ^ Dumortier, B.-C. (1822). Commentationes botanicae. Observations botaniques, dédiées à la Société d'Horticulture de Tournay (PDF). Tournay: Imprimerie de Ch. Casterman-Dieu, Rue de pont No. 10. pp. [i], [1]-116, [1, tabl., err.] Archived from the original (PDF) on 6 October 2015 – via Algaebase.
- ^ Labenhorst, L. Flora europaea algarum at dulcis et subsuminae (1864–1868).제1장 Algas diatomaceas collectens, cum pigure generium generium xylographice (1864). 페이지 1 ~ 359.Lipsiae [라이프치히] :아푸드 에두아르둠 금머럼
- ^ Haeckel, E. (1878년)Das Protistenreich는 2014년 11월 10일 웨이백 머신에 보관되었습니다.
- ^ Engler, A. & Gilg, E. (1919년)Icllabus der Pfranzenfamilien : eine ü베르시히트 über das Gesamte Pfranzensystem mit be be be geum ber ücsichtigung der Medizinal - und Nutzpflanzen, nebst einer üversicht üver die flen über die flen ü베르시히트 über die flanzflanze die flant flant flanzau.베를린, 오후 395시
- ^ 【α】【μ】Liddell, Henry George; Scott, Robert; 페르세우스 프로젝트의 그리스 영어 사전
- ^ ιιμο 【중략】Liddell, Henry George; Scott, Robert; 페르세우스 프로젝트의 그리스 영어 사전
- ^ 【α】【μ】.Liddell, Henry George; Scott, Robert; 페르세우스 프로젝트의 그리스 영어 사전
- ^ The Compact Oxford English Dictionary. Clarendon Press. 1971. ISBN 0918414083.
- ^ "The Air You're Breathing? A Diatom Made That". Live Science. 11 June 2014. Archived from the original on 30 April 2018. Retrieved 30 April 2018.
- ^ "What are Diatoms?". Diatoms of North America. Archived from the original on 25 January 2020. Retrieved 28 January 2020.
- ^ Treguer, P.; Nelson, D. M.; Van Bennekom, A. J.; Demaster, D. J.; Leynaert, A.; Queguiner, B. (1995). "The Silica Balance in the World Ocean: A Reestimate". Science. 268 (5209): 375–9. Bibcode:1995Sci...268..375T. doi:10.1126/science.268.5209.375. PMID 17746543. S2CID 5672525.
- ^ "King's College London – Lake Megachad". www.kcl.ac.uk. Archived from the original on 27 November 2018. Retrieved 5 May 2018.
- ^ Bristow, C.S.; Hudson-Edwards, K.A.; Chappell, A. (2010). "Fertilizing the Amazon and equatorial Atlantic with West African dust". Geophys. Res. Lett. 37 (14): L14807. Bibcode:2010GeoRL..3714807B. doi:10.1029/2010GL043486. S2CID 128466273.
- ^ a b c d e f g Grethe R. Hasle; Erik E. Syvertsen; Karen A. Steidinger; Karl Tangen (25 January 1996). "Marine Diatoms". In Carmelo R. Tomas (ed.). Identifying Marine Diatoms and Dinoflagellates. Academic Press. pp. 5–385. ISBN 978-0-08-053441-1. Archived from the original on 3 January 2014. Retrieved 13 November 2013.
- ^ "Gas Guzzlers". Archived from the original on 22 May 2018. Retrieved 22 May 2018.
- ^ "More on Diatoms". University of California Museum of Paleontology. Archived from the original on 4 October 2012. Retrieved 20 May 2015.
- ^ van den Hoek, C.; Mann, D.G.; Jahns, H.M. (1995). Algae: an introduction to Phycology. Cambridge: Cambridge University Press. pp. 165–218. ISBN 978-0-521-31687-3.
- ^ Nakayama, T.; Kamikawa, R.; Tanifuji, G.; Kashiyama, Y.; Ohkouchi, N.; Archibald, J. M.; Inagaki, Y. (2014). "Complete genome of a nonphotosynthetic cyanobacterium in a diatom reveals recent adaptations to an intracellular lifestyle". Proceedings of the National Academy of Sciences of the United States of America. 111 (31): 11407–11412. Bibcode:2014PNAS..11111407N. doi:10.1073/pnas.1405222111. PMC 4128115. PMID 25049384.
- ^ Pierella Karlusich, Juan José; Pelletier, Eric; Lombard, Fabien; Carsique, Madeline; Dvorak, Etienne; Colin, Sébastien; Picheral, Marc; Cornejo-Castillo, Francisco M.; Acinas, Silvia G.; Pepperkok, Rainer; Karsenti, Eric (6 July 2021). "Global distribution patterns of marine nitrogen-fixers by imaging and molecular methods". Nature Communications. 12 (1): 4160. Bibcode:2021NatCo..12.4160P. doi:10.1038/s41467-021-24299-y. ISSN 2041-1723. PMC 8260585. PMID 34230473.
- ^ Wehr, J. D.; Sheath, R. G.; Kociolek, J. P., eds. (2015). Freshwater Algae of North America: Ecology and Classification (2nd ed.). San Diego: Academic Press. ISBN 978-0-12-385876-4.
- ^ Girard, Vincent; Saint Martin, Simona; Buffetaut, Eric; Saint Martin, Jean-Paul; Néraudeau, Didier; Peyrot, Daniel; Roghi, Guido; Ragazzi, Eugenio; Suteethorn, Varavudh (2020). "Thai amber: insights into early diatom history?". BSGF - Earth Sciences Bulletin. 191: 23. doi:10.1051/bsgf/2020028. ISSN 1777-5817.
- ^ Colin, S., Coelho, L.P., Sunagawa, S., Bowler, C., Karsenti, E., Bork, P., Pepperkok, R. 및 De Vargas, C. (2017) "환경 미생물 생물 생물학과 생태학을 위한 정량적 3D 영상촬영"eLife, 6: e26066.doi: 10.7554/eLife.26066.002.
 자료는 Wayback Machine에서 2017년 10월 16일 아카이브된 Creative Commons Attribution 4.0 International License에 따라 제공되는 이 소스로부터 복사되었습니다.
자료는 Wayback Machine에서 2017년 10월 16일 아카이브된 Creative Commons Attribution 4.0 International License에 따라 제공되는 이 소스로부터 복사되었습니다. - ^ 아북극 태평양 내부 공간 2020년 10월 27일 웨이백 머신에 보관NASA 지구 탐험대, 2018년 9월 4일
 이 문서에는 퍼블릭 도메인에 있는 이 소스로부터의 텍스트가 포함되어 있습니다..
이 문서에는 퍼블릭 도메인에 있는 이 소스로부터의 텍스트가 포함되어 있습니다.. - ^ Rousseaux, Cecile S.; Gregg, Watson W. (2015). "Recent decadal trends in global phytoplankton composition". Global Biogeochemical Cycles. 29 (10): 1674–1688. Bibcode:2015GBioC..29.1674R. doi:10.1002/2015GB005139.
- ^ Parker, Andrew R.; Townley, Helen E. (2007). "Biomimetics of photonic nanostructures". Nature Nanotechnology. 2 (6): 347–53. Bibcode:2007NatNa...2..347P. doi:10.1038/nnano.2007.152. PMID 18654305.
- ^ Gordon, Richard; Losic, Dusan; Tiffany, Mary Ann; Nagy, Stephen S.; Sterrenburg, Frithjof A.S. (2009). "The Glass Menagerie: Diatoms for novel applications in nanotechnology". Trends in Biotechnology. 27 (2): 116–27. doi:10.1016/j.tibtech.2008.11.003. PMID 19167770.
- ^ a b c d e Rita A. Horner (2002). A taxonomic guide to some common marine phytoplankton. Biopress. pp. 25–30. ISBN 978-0-948737-65-7. Archived from the original on 1 August 2020. Retrieved 13 November 2013.
- ^ "Glass in Nature". The Corning Museum of Glass. Archived from the original on 7 March 2013. Retrieved 19 February 2013.
- ^ Taylor, J. C., Harding, W. R. 및 Archibald, C.(2007)남아프리카 공화국의 일반적인 규조류에 대한 일러스트레이션 안내서입니다.게지나:물 조사 위원회ISBN 9781770054844.
- ^ Mishra, M., Arukha, A.P., Bashir, T., Yadav, D. 및 Prasad, G.B.K.S. (2017) "규조류의 모든 새로운 면: 나노물질 및 그 이상의 잠재적 원천"미생물학 분야 프런티어, 8:1239.doi:10.3389/fmicb.2017.01239. 자료는 Wayback Machine에서 2017년 10월 16일 아카이브된 Creative Commons Attribution 4.0 International License에 따라 제공되는 이 소스로부터 복사되었습니다.
- ^ a b Zhang, D.; Wang, Y.; Cai, J.; Pan, J.; Jiang, X.; Jiang, Y. (2012). "Bio-manufacturing technology based on diatom micro- and nanostructure". Chinese Science Bulletin. 57 (30): 3836–3849. Bibcode:2012ChSBu..57.3836Z. doi:10.1007/s11434-012-5410-x.
- ^ Padisák, Judit; Soróczki-Pintér, Éva; Rezner, Zsuzsanna (2003), Martens, Koen (ed.), "Sinking properties of some phytoplankton shapes and the relation of form resistance to morphological diversity of plankton – an experimental study" (PDF), Aquatic Biodiversity: A Celebratory Volume in Honour of Henri J. Dumont, Developments in Hydrobiology, Springer Netherlands, pp. 243–257, doi:10.1007/978-94-007-1084-9_18, ISBN 9789400710849, archived from the original (PDF) on 23 July 2018, retrieved 4 October 2019
- ^ Anderson, Lars W. J.; Sweeney, Beatrice M. (1 May 1977). "Diel changes in sedimentation characteristics of Ditylum brightwelli: Changes in cellular lipid and effects of respiratory inhibitors and ion-transport modifiers1". Limnology and Oceanography. 22 (3): 539–552. Bibcode:1977LimOc..22..539A. doi:10.4319/lo.1977.22.3.0539. ISSN 1939-5590.
- ^ Poulsen, Nicole C.; Spector, Ilan; Spurck, Timothy P.; Schultz, Thomas F.; Wetherbee, Richard (1 September 1999). "Diatom gliding is the result of an actin-myosin motility system". Cell Motility and the Cytoskeleton. 44 (1): 23–33. doi:10.1002/(SICI)1097-0169(199909)44:1<23::AID-CM2>3.0.CO;2-D. ISSN 1097-0169. PMID 10470016.
- ^ Mann, David G. (February 2010). "raphid diatoms". The Tree of Life Web Project. Archived from the original on 27 September 2019. Retrieved 27 September 2019.
- ^ a b G. Drebes (1 January 1977). "Chapter 9: Sexuality". In Dietrich Werner (ed.). The Biology of Diatoms. Botanical Monographs. Vol. 13. University of California Press. pp. 250–283. ISBN 978-0-520-03400-6. Archived from the original on 1 August 2020. Retrieved 14 November 2013.
- ^ Azam, Farooq; Bidle, Kay D. (1999). "Accelerated dissolution of diatom silica by marine bacterial assemblages". Nature. 397 (6719): 508–12. Bibcode:1999Natur.397..508B. doi:10.1038/17351. S2CID 4397909. INIST:1755031.
- ^ Zakharova, Yulia R.; Galachyants, Yuri P.; Kurilkina, Maria I.; Likhoshvay, Alexander V.; Petrova, Darya P.; Shishlyannikov, Sergey M.; Ravin, Nikolai V.; Mardanov, Andrey V.; Beletsky, Alexey V.; Likhoshway, Yelena V. (2013). "The Structure of Microbial Community and Degradation of Diatoms in the Deep Near-Bottom Layer of Lake Baikal". PLOS ONE. 8 (4): e59977. Bibcode:2013PLoSO...859977Z. doi:10.1371/journal.pone.0059977. PMC 3613400. PMID 23560063.
- ^ a b Treguer, P.; Nelson, D. M.; Van Bennekom, A. J.; Demaster, D. J.; Leynaert, A.; Queguiner, B. (1995). "The Silica Balance in the World Ocean: A Reestimate". Science. 268 (5209): 375–9. Bibcode:1995Sci...268..375T. doi:10.1126/science.268.5209.375. PMID 17746543. S2CID 5672525.
- ^ Kilias, Estelle S.; Junges, Leandro; Šupraha, Luka; Leonard, Guy; Metfies, Katja; Richards, Thomas A. (2020). "Chytrid fungi distribution and co-occurrence with diatoms correlate with sea ice melt in the Arctic Ocean". Communications Biology. 3 (1): 183. doi:10.1038/s42003-020-0891-7. PMC 7174370. PMID 32317738. S2CID 216033140.
- ^ a b Egge, J. K.; Aksnes, D. L. (1992). "Silicate as regulating nutrient in phytoplankton competition". Mar. Ecol. Prog. Ser. 83: 281–289. Bibcode:1992MEPS...83..281E. doi:10.3354/meps083281.
- ^ a b c Furnas, Miles J. (1990). "In situ growth rates of marine phytoplankton: Approaches to measurement, community and species growth rates". Journal of Plankton Research. 12 (6): 1117–51. doi:10.1093/plankt/12.6.1117. INIST:5474600.
- ^ a b Yool, Andrew; Tyrrell, Toby (2003). "Role of diatoms in regulating the ocean's silicon cycle". Global Biogeochemical Cycles. 17 (4): n/a. Bibcode:2003GBioC..17.1103Y. CiteSeerX 10.1.1.394.3912. doi:10.1029/2002GB002018. S2CID 16849373.
- ^ Lipps, Jere H. (1970). "Plankton Evolution". Evolution. 24 (1): 1–22. doi:10.2307/2406711. JSTOR 2406711. PMID 28563010.
- ^ 디디모, 외계인 우리 사이2015년 10월 7일 캐나다 가상박물관 Wayback Machine 가상전시회 아카이브
- ^ "DEP Reports Didymo Discovered in the West Branch Farmington River. Retrieved on 2014-01-15". Archived from the original on 16 February 2015. Retrieved 27 April 2015.
- ^ "Didymo Stakeholder Update – 31 October 2008". MAF Biosecurity New Zealand www.biosecurity.govt.nz. Archived from the original on 12 February 2013. Retrieved 1 December 2013.
- ^ a b Dugdale, R. C.; Wilkerson, F. P. (1998). "Silicate regulation of new production in the equatorial Pacific upwelling". Nature. 391 (6664): 270–273. Bibcode:1998Natur.391..270D. doi:10.1038/34630. S2CID 4394149.
- ^ Smetacek, V. S. (1985). "Role of sinking in diatom life-history cycles: Ecological, evolutionary and geological significance". Mar. Biol. 84 (3): 239–251. doi:10.1007/BF00392493. S2CID 85054779.
- ^ Raven, J. A. (1983). "The transport and function of silicon in plants". Biol. Rev. 58 (2): 179–207. doi:10.1111/j.1469-185X.1983.tb00385.x. S2CID 86067386.
- ^ Milligan, A. J.; Morel, F. M. M. (2002). "A proton buffering role for silica in diatoms". Science. 297 (5588): 1848–1850. Bibcode:2002Sci...297.1848M. doi:10.1126/science.1074958. PMID 12228711. S2CID 206507070.
- ^ a b Chamberlain, C. J. (1901) 미국 시카고 대학 출판부 식물 조직학에서의 방법
- ^ Armstrong, E; Rogerson, A; Leftley, Jw (2000). "Utilisation of seaweed carbon by three surface-associated heterotrophic protists, Stereomyxa ramosa, Nitzschia alba and Labyrinthula sp". Aquatic Microbial Ecology. 21: 49–57. doi:10.3354/ame021049. ISSN 0948-3055.
- ^ LEWIN, JOYCE; LEWIN, R. A. (1967). "Culture and Nutrition of Some Apochlorotic Diatoms of the Genus Nitzschia". Microbiology. 46 (3): 361–367. doi:10.1099/00221287-46-3-361. ISSN 1350-0872.
- ^ "Diatoms". Archived from the original on 2 February 2016. Retrieved 13 February 2016.
- ^ a b c Thamatrakoln, K.; Alverson, A.J.; Hildebrand, M. (2006). "Comparative Sequence Analysis of Diatom Silicon Transporters: Toward a Mechanistic Model of Silicon Transport". Journal of Phycology. 42 (4): 822–834. doi:10.1111/j.1529-8817.2006.00233.x. S2CID 86674657.
- ^ Kröger, Nils; Deutzmann, Rainer; Manfred, Sumper (November 1999). "Polycationic Peptides from Diatom Biosilica That Direct Silica Nanosphere Formation". Science. 286 (5442): 1129–1132. doi:10.1126/science.286.5442.1129. PMID 10550045. S2CID 10925689.[영구 데드링크]
- ^ Kroger, Nils (2007). Handbook of Biomineralization: Biological Aspects and Structure Formation. Weinheim, Germany: Wiley-VCH Verlag GmbH. pp. chapter 3.
- ^ Allen, Andrew E.; Dupont, Christopher L.; Oborník, Miroslav; Horák, Aleš; Nunes-Nesi, Adriano; McCrow, John P.; Zheng, Hong; Johnson, Daniel A.; Hu, Hanhua; Fernie, Alisdair R.; Bowler, Chris (2011). "Evolution and metabolic significance of the urea cycle in photosynthetic diatoms". Nature. 473 (7346): 203–7. Bibcode:2011Natur.473..203A. doi:10.1038/nature10074. PMID 21562560. S2CID 4350884. Archived from the original on 19 February 2022. Retrieved 20 May 2021.
- "Animal-like urea cycle in ocean's tiny diatoms enables marine phytoplankton to use carbon and nitrogen from their environment". ScienceDaily (Press release). 12 May 2011.
- ^ Guiry, M.D. (2012). "How many species of algae are there?". Journal of Phycology. 48 (5): 1057–1063. doi:10.1111/j.1529-8817.2012.01222.x. PMID 27011267. S2CID 30911529.
- ^ Frank Eric Round; R. M. Crawford; D. G. Mann (1990). The Diatoms: Biology & Morphology of the Genera. Cambridge University Press. ISBN 978-0-521-36318-1. Archived from the original on 24 May 2021. Retrieved 13 November 2013.[페이지 필요]
- ^ 캔터-런드, H., 룬드, J.W.G.(1995).민물 조류: 그들의 미시적인 세계는 바이오프레스 유한회사라고 설명했습니다.ISBN 0-948737-25-5.[page needed]
- ^ Mann, David G. (2005). "The species concept in diatoms: Evidence for morphologically distinct, sympatric gamodemes in four epipelic species". Plant Systematics and Evolution. 164 (1/4): 215–37. doi:10.1007/BF00940439. JSTOR 23675282. S2CID 37684109.
- ^ Fourtanier, Elisabeth; Kociolek, J. Patrick (1999). "Catalogue of the Diatom Genera". Diatom Research. 14 (1): 1–190. doi:10.1080/0269249X.1999.9705462.
- ^ 2020년 7월 현재, 세계 해양 종 등록에는 모든 서식지에서 1,356개의 규조속 이름이 등록되어 있으며, 이 중 1,248개가 "승인"되어 있다.
- ^ 2020년 7월 세계해양종등록부에 문의하면 299개의 "화석만" 속명이 반환되며, 이 중 285개는 "승인"된다.
- ^ Theriot, Edward C.; Cannone, Jamie J.; Gutell, Robin R.; Alverson, Andrew J. (2009). "The limits of nuclear-encoded SSU rDNA for resolving the diatom phylogeny". European Journal of Phycology. 4 (3): 277–290. doi:10.5091/plecevo.2010.418. PMC 2835975. PMID 20224747.
- ^ Theriot, Edward C.; Ashworth, Matt; Ruck, Elizabeth; Nakov, Teofil; Jansen, Robert K. (2010). "A preliminary multigene phylogeny of the diatoms (Bacillariophyta): challenges for future research". Plant Ecology and Evolution. 143 (3): 277–290. doi:10.1080/09670260902749159. PMC 2835975. PMID 20224747.
- ^ Parks, Matthew B.; Wickett, Norman J.; Alverson, Andrew J. (2018). "Signal, uncertainty, and conflict in phylogenomic data for a diverse lineage of microbial eukaryotes (Diatoms, Bacillariophyta)". Molecular Biology and Evolution. 35 (1): 80–93. doi:10.1093/molbev/msx268. PMC 5850769. PMID 29040712.
- ^ Medlin, L.K.; Desdevises, Y. (2020). "Review of the phylogenetic reconstruction of the diatoms using molecular tools with an analysis of a seven gene data set using multiple outgroups and morphological data for a total evidence approach" (PDF). Phycologia. in press. Archived (PDF) from the original on 20 July 2020. Retrieved 20 July 2020.
- ^ Adl, Sina M.; Bass, David; Lane, Christopher E.; Lukeš, Julius; Schoch, Conrad L.; Smirnov, Alexey; Agatha, Sabine; Berney, Cedric; Brown, Matthew W. (26 September 2018). "Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes". Journal of Eukaryotic Microbiology. 66 (1): 4–119. doi:10.1111/jeu.12691. PMC 6492006. PMID 30257078.
- ^ Mishra는, Meerambika, Arukha, Ananta P., 바시르, Tufail;야다브, Dhananjay, 프라사드, G.B.K.S.(57월 2017년)."모든 뉴 FacesDiatoms의:.잠재원 Nanomaterials과 Beyond".미생물학에 국경.둔 미디어 SA.8시 1239년. doi:10.3389/fmicb.2017.01239.ISSN 1664-302X.PMC 5496942.PMID 28725218.재료는 창조적 공용 귀인 4.0국제 라이센스 하에 1610월 2017년 Archived은 승객을 머신에 이용 가능한 이 원본에서 복사되었다.
- ^ a b Kooistra, Wiebe H.C.F.; Medlin, Linda K. (1996). "Evolution of the Diatoms (Bacillariophyta)". Molecular Phylogenetics and Evolution. 6 (3): 391–407. doi:10.1006/mpev.1996.0088. PMID 8975694.
- ^ Schieber, Jürgen; Krinsley, Dave; Riciputi, Lee (2000). "Diagenetic origin of quartz silt in mudstones and implications for silica cycling". Nature. 406 (6799): 981–5. Bibcode:2000Natur.406..981S. doi:10.1038/35023143. PMID 10984049. S2CID 4417951.
- ^ Medlin, L. K.; Kooistra, W. H. C. F.; Gersonde, R.; Sims, P. A.; Wellbrock, U. (1997). "Is the origin of the diatoms related to the end-Permian mass extinction?". Nova Hedwigia. 65 (1–4): 1–11. doi:10.1127/nova.hedwigia/65/1997/1. hdl:10013/epic.12689.
- ^ Raven, J. A.; Waite, A. M. (2004). "The evolution of silicification in diatoms: Inescapable sinking and sinking as escape?". New Phytologist. 162 (1): 45–61. doi:10.1111/j.1469-8137.2004.01022.x. JSTOR 1514475.
- ^ R. Siever; Stephen Henry Schneider; Penelope J. Boston (January 1993). "Silica in the oceans: biological-geological interplay". Scientists on Gaia. MIT Press. pp. 287–295. ISBN 978-0-262-69160-4. Retrieved 14 November 2013.
- ^ Kidder, David L.; Erwin, Douglas H. (2001). "Secular Distribution of Biogenic Silica through the Phanerozoic: Comparison of Silica-Replaced Fossils and Bedded Cherts at the Series Level". The Journal of Geology. 109 (4): 509–22. Bibcode:2001JG....109..509K. doi:10.1086/320794. S2CID 128401816.
- ^ Grenne, Tor; Slack, John F. (2003). "Paleozoic and Mesozoic silica-rich seawater: Evidence from hematitic chert (jasper) deposits". Geology. 31 (4): 319–22. Bibcode:2003Geo....31..319G. doi:10.1130/0091-7613(2003)031<0319:PAMSRS>2.0.CO;2. INIST:14692468.
- ^ Racki, G; Cordey, Fabrice (2000). "Radiolarian palaeoecology and radiolarites: Is the present the key to the past?". Earth-Science Reviews. 52 (1): 83–120. Bibcode:2000ESRv...52...83R. doi:10.1016/S0012-8252(00)00024-6.
- ^ Maldonado, Manuel; Carmona, M. Carmen; Uriz, María J.; Cruzado, Antonio (1999). "Decline in Mesozoic reef-building sponges explained by silicon limitation". Nature. 401 (6755): 785–8. Bibcode:1999Natur.401..785M. doi:10.1038/44560. S2CID 205034177. INIST:1990263.
- ^ Harper, Howard E.; Knoll, Andrew H. (1975). "Silica, diatoms, and Cenozoic radiolarian evolution". Geology. 3 (4): 175–7. Bibcode:1975Geo.....3..175H. doi:10.1130/0091-7613(1975)3<175:SDACRE>2.0.CO;2.
- ^ Falkowski, P. G.; Katz, Miriam E.; Knoll, Andrew H.; Quigg, Antonietta; Raven, John A.; Schofield, Oscar; Taylor, F. J. R. (2004). "The Evolution of Modern Eukaryotic Phytoplankton". Science. 305 (5682): 354–60. Bibcode:2004Sci...305..354F. CiteSeerX 10.1.1.598.7930. doi:10.1126/science.1095964. PMID 15256663. S2CID 451773.
- ^ Kidder, D. L.; Gierlowski-Kordesch, E. H. (2005). "Impact of Grassland Radiation on the Nonmarine Silica Cycle and Miocene Diatomite". PALAIOS. 20 (2): 198–206. Bibcode:2005Palai..20..198K. doi:10.2110/palo.2003.p03-108. JSTOR 27670327. S2CID 140584104.
- ^ a b Lazarus, David; Barron, John; Renaudie, Johan; Diver, Patrick; Türke, Andreas (2014). "Cenozoic Planktonic Marine Diatom Diversity and Correlation to Climate Change". PLOS ONE. 9 (1): e84857. Bibcode:2014PLoSO...984857L. doi:10.1371/journal.pone.0084857. PMC 3898954. PMID 24465441.
- ^ IPCC 코어 쓰기 팀, 2007.기후 변화 2007: 종합 보고서. 104.
- ^ a b Scherer, R. P.; Gladenkov, A. Yu.; Barron, J. A. (2007). "Methods and applications of Cenozoic marine diatom biostratigraphy". Paleontological Society Papers. 13: 61–83. doi:10.1017/S1089332600001467.
- ^ "What Are "Proxy" Data? National Centers for Environmental Information (NCEI) formerly known as National Climatic Data Center (NCDC)". www.ncdc.noaa.gov. Archived from the original on 8 March 2020. Retrieved 20 October 2020.
- ^ a b 스완, G.E.A.;Snelling, A.M.(1월 6일 2015년)."당신을 북서부 태평양에서 MIS4–5e에서Photic 지역 변화".과거의 기후.코페르니쿠스는 회사. 11(1):15–25.Bibcode:2015CliPa..11...15S.doi:10.5194/cp-11-15-2015.ISSN 1814-9332.재료는 창조적 공용 귀인 3.0국제 라이센스 하에 2월 23일 2011년 Archived은 승객을 머신에 이용 가능한 이 원본에서 복사되었다.
- ^ Sigman, Daniel M.; Jaccard, Samuel L.; Haug, Gerald H. (2004). "Polar ocean stratification in a cold climate". Nature. Springer Science and Business Media LLC. 428 (6978): 59–63. Bibcode:2004Natur.428...59S. doi:10.1038/nature02357. ISSN 0028-0836. PMID 14999278. S2CID 4329978. Archived from the original on 19 February 2022. Retrieved 15 December 2021.
- ^ a b Haug, Gerald H.; Ganopolski, Andrey; Sigman, Daniel M.; Rosell-Mele, Antoni; Swann, George E. A.; Tiedemann, Ralf; Jaccard, Samuel L.; Bollmann, Jörg; Maslin, Mark A.; Leng, Melanie J.; Eglinton, Geoffrey (2005). "North Pacific seasonality and the glaciation of North America 2.7 million years ago". Nature. Springer Science and Business Media LLC. 433 (7028): 821–825. Bibcode:2005Natur.433..821H. doi:10.1038/nature03332. ISSN 0028-0836. PMID 15729332. S2CID 24116155. Archived from the original on 19 February 2022. Retrieved 15 December 2021.
- ^ Swann, George E. A.; Maslin, Mark A.; Leng, Melanie J.; Sloane, Hilary J.; Haug, Gerald H. (24 February 2006). "Diatom δ18O evidence for the development of the modern halocline system in the subarctic northwest Pacific at the onset of major Northern Hemisphere glaciation". Paleoceanography. American Geophysical Union (AGU). 21 (1): n/a. Bibcode:2006PalOc..21.1009S. doi:10.1029/2005pa001147. ISSN 0883-8305.
- ^ Nie, Junsheng; King, John; Liu, Zhengyu; Clemens, Steve; Prell, Warren; Fang, Xiaomin (2008). "Surface-water freshening: A cause for the onset of North Pacific stratification from 2.75 Ma onward?". Global and Planetary Change. Elsevier BV. 64 (1–2): 49–52. Bibcode:2008GPC....64...49N. doi:10.1016/j.gloplacha.2008.08.003. ISSN 0921-8181.
- ^ Swann, George E.A. (2010). "Salinity changes in the North West Pacific Ocean during the late Pliocene/early Quaternary from 2.73Ma to 2.52Ma" (PDF). Earth and Planetary Science Letters. Elsevier BV. 297 (1–2): 332–338. Bibcode:2010E&PSL.297..332S. doi:10.1016/j.epsl.2010.06.035. ISSN 0012-821X. Archived (PDF) from the original on 19 July 2018. Retrieved 15 December 2021.
- ^ Sarnthein, M.; Gebhardt, H.; Kiefer, T.; Kucera, M.; Cook, M.; Erlenkeuser, H. (2004). "Mid Holocene origin of the sea-surface salinity low in the subarctic North Pacific". Quaternary Science Reviews. Elsevier BV. 23 (20–22): 2089–2099. Bibcode:2004QSRv...23.2089S. doi:10.1016/j.quascirev.2004.08.008. ISSN 0277-3791.
- ^ Jaccard, S.L.; Galbraith, E.D.; Sigman, D.M.; Haug, G.H. (2010). "A pervasive link between Antarctic ice core and subarctic Pacific sediment records over the past 800kyrs". Quaternary Science Reviews. Elsevier BV. 29 (1–2): 206–212. Bibcode:2010QSRv...29..206J. doi:10.1016/j.quascirev.2009.10.007. ISSN 0277-3791.
- ^ Galbraith, Eric D.; Kienast, Markus; Jaccard, Samuel L.; Pedersen, Thomas F.; Brunelle, Brigitte G.; Sigman, Daniel M.; Kiefer, Thorsten (23 May 2008). "Consistent relationship between global climate and surface nitrate utilization in the western subarctic Pacific throughout the last 500 ka" (PDF). Paleoceanography. American Geophysical Union (AGU). 23 (2): n/a. Bibcode:2008PalOc..23.2212G. doi:10.1029/2007pa001518. ISSN 0883-8305. Archived (PDF) from the original on 15 July 2021. Retrieved 15 December 2021.
- ^ Brunelle, Brigitte G.; Sigman, Daniel M.; Jaccard, Samuel L.; Keigwin, Lloyd D.; Plessen, Birgit; Schettler, Georg; Cook, Mea S.; Haug, Gerald H. (2010). "Glacial/interglacial changes in nutrient supply and stratification in the western subarctic North Pacific since the penultimate glacial maximum". Quaternary Science Reviews. Elsevier BV. 29 (19–20): 2579–2590. Bibcode:2010QSRv...29.2579B. doi:10.1016/j.quascirev.2010.03.010. ISSN 0277-3791.
- ^ Kohfeld, Karen E.; Chase, Zanna (2011). "Controls on deglacial changes in biogenic fluxes in the North Pacific Ocean". Quaternary Science Reviews. Elsevier BV. 30 (23–24): 3350–3363. Bibcode:2011QSRv...30.3350K. doi:10.1016/j.quascirev.2011.08.007. ISSN 0277-3791.
- ^ Harwood, D. M.; Nikolaev, V. A.; Winter, D. M. (2007). "Cretaceous record of diatom evolution, radiation, and expansion". Paleontological Society Papers. 13: 33–59. doi:10.1017/S1089332600001455.
- ^ Strelnikova, N. I. (1990). "Evolution of diatoms during the Cretaceous and Paleogene periods". In Simola, H. (ed.). Proceedings of the Tenth International Diatom Symposium. Koenigstein: Koeltz Scientific Books. pp. 195–204. ISBN 3-87429-307-6.
- ^ Baldauf, J. G. (1993). "Middle Eocene through early Miocene diatom floral turnover". In Prothero, D.; Berggren, W. H. (eds.). Eocene-Oligocene climatic and biotic evolution. Princeton: Princeton University Press. pp. 310–326. ISBN 0-691-02542-8.
- ^ Barron, J. A. (2003). "Appearance and extinction of planktonic diatoms during the past 18 m.y. in the Pacific and Southern oceans". Diatom Research. 18: 203–224. doi:10.1080/0269249x.2003.9705588. S2CID 84781882.
- ^ Scala, S.; Carels, N; Falciatore, A; Chiusano, M. L.; Bowler, C (2002). "Genome Properties of the Diatom Phaeodactylum tricornutum". Plant Physiology. 129 (3): 993–1002. doi:10.1104/pp.010713. PMC 166495. PMID 12114555.
- ^ Maheswari, U.; Montsant, A; Goll, J; Krishnasamy, S; Rajyashri, K. R.; Patell, V. M.; Bowler, C (2004). "The Diatom EST Database". Nucleic Acids Research. 33 (Database issue): D344–7. doi:10.1093/nar/gki121. PMC 540075. PMID 15608213.
- ^ Montsant, A.; Jabbari, K; Maheswari, U; Bowler, C (2005). "Comparative Genomics of the Pennate Diatom Phaeodactylum tricornutum". Plant Physiology. 137 (2): 500–13. doi:10.1104/pp.104.052829. PMC 1065351. PMID 15665249.
- ^ Maheswari, U.; Mock, T.; Armbrust, E. V.; Bowler, C. (2009). "Update of the Diatom EST Database: A new tool for digital transcriptomics". Nucleic Acids Research. 37 (Database issue): D1001–5. doi:10.1093/nar/gkn905. PMC 2686495. PMID 19029140.
- ^ a b Armbrust, E. V.; Berges, John A.; Bowler, Chris; Green, Beverley R.; Martinez, Diego; Putnam, Nicholas H.; Zhou, Shiguo; Allen, Andrew E.; Apt, Kirk E.; Bechner, Michael; Brzezinski, Mark A.; Chaal, Balbir K.; Chiovitti, Anthony; Davis, Aubrey K.; Demarest, Mark S.; Detter, J. Chris; Glavina, Tijana; Goodstein, David; Hadi, Masood Z.; Hellsten, Uffe; Hildebrand, Mark; Jenkins, Bethany D.; Jurka, Jerzy; Kapitonov, Vladimir V.; Kröger, Nils; Lau, Winnie W. Y.; Lane, Todd W.; Larimer, Frank W.; Lippmeier, J. Casey; et al. (2004). "The Genome of the Diatom Thalassiosira Pseudonana: Ecology, Evolution, and Metabolism". Science. 306 (5693): 79–86. Bibcode:2004Sci...306...79A. CiteSeerX 10.1.1.690.4884. doi:10.1126/science.1101156. PMID 15459382. S2CID 8593895.
- ^ a b c d Bowler, Chris; Allen, Andrew E.; Badger, Jonathan H.; Grimwood, Jane; Jabbari, Kamel; Kuo, Alan; Maheswari, Uma; Martens, Cindy; Maumus, Florian; Otillar, Robert P.; Rayko, Edda; Salamov, Asaf; Vandepoele, Klaas; Beszteri, Bank; Gruber, Ansgar; Heijde, Marc; Katinka, Michael; Mock, Thomas; Valentin, Klaus; Verret, Fréderic; Berges, John A.; Brownlee, Colin; Cadoret, Jean-Paul; Chiovitti, Anthony; Choi, Chang Jae; Coesel, Sacha; De Martino, Alessandra; Detter, J. Chris; Durkin, Colleen; et al. (2008). "The Phaeodactylum genome reveals the evolutionary history of diatom genomes". Nature. 456 (7219): 239–44. Bibcode:2008Natur.456..239B. doi:10.1038/nature07410. PMID 18923393. S2CID 4415177.
- ^ Roy, S. W.; Penny, D. (2007). "A Very High Fraction of Unique Intron Positions in the Intron-Rich Diatom Thalassiosira pseudonana Indicates Widespread Intron Gain". Molecular Biology and Evolution. 24 (7): 1447–57. doi:10.1093/molbev/msm048. PMID 17350938.
- ^ Maumus, Florian; Allen, Andrew E; Mhiri, Corinne; Hu, Hanhua; Jabbari, Kamel; Vardi, Assaf; Grandbastien, Marie-Angèle; Bowler, Chris (2009). "Potential impact of stress activated retrotransposons on genome evolution in a marine diatom". BMC Genomics. 10: 624. doi:10.1186/1471-2164-10-624. PMC 2806351. PMID 20028555.
- ^ Moustafa, A.; Beszteri, B.; Maier, U. G.; Bowler, C.; Valentin, K.; Bhattacharya, D. (2009). "Genomic Footprints of a Cryptic Plastid Endosymbiosis in Diatoms" (PDF). Science. 324 (5935): 1724–6. Bibcode:2009Sci...324.1724M. doi:10.1126/science.1172983. PMID 19556510. S2CID 11408339. Archived from the original (PDF) on 21 April 2014. Retrieved 13 January 2019.
- ^ a b Kroth, Peter G.; Bones, Atle M.; et al. (October 2018). "Genome editing in diatoms: achievements and goals". Plant Cell Reports. 37 (10): 1401–1408. doi:10.1007/s00299-018-2334-1. PMID 30167805. S2CID 52133809. Archived from the original on 19 February 2022. Retrieved 20 May 2021.
- ^ a b Falciatore, Angela; Casotti, Raffaella; et al. (May 2015). "Tranformation of Nonselectablporter Genes in Marine Diatomse Re". Marine Biotechnology. 1 (3): 239–251. doi:10.1007/PL00011773. PMID 10383998. S2CID 22267097.
- ^ a b c d e Karas, Bogumil J.; Diner, Rachel E.; et al. (21 April 2015). "Designer diatom episomes delivered by bacterial conjugation". Nature Communications. 6 (1): 6925. Bibcode:2015NatCo...6.6925K. doi:10.1038/ncomms7925. ISSN 2041-1723. PMC 4411287. PMID 25897682.
- ^ a b c d Slattery, Samuel S.; Diamond, Andrew; et al. (16 February 2018). "An Expanded Plasmid-Based Genetic Toolbox Enables Cas9 Genome Editing and Stable Maintenance of Synthetic Pathways in Phaeodactylum tricornutum". ACS Synthetic Biology. 7 (2): 328–338. doi:10.1021/acssynbio.7b00191. PMID 29298053.
- ^ Nymark, Marianne; Sharma, Amit Kumar; et al. (July 2016). "A CRISPR/Cas9 system adapted for gene editing in marine algae". Scientific Reports. 6 (1): 24951. Bibcode:2016NatSR...624951N. doi:10.1038/srep24951. PMC 4842962. PMID 27108533.
- ^ Mishra, M; Arukha, AP; Bashir, T; Yadav, D; Gbks, Prasad (2017). "All New Faces of Diatoms: Potential Source of Nanomaterials and Beyond". Front Microbiol. 8: 1239. doi:10.3389/fmicb.2017.01239. PMC 5496942. PMID 28725218.
- ^ Reka, Arianit A.; Pavlovski, Blagoj; Makreski, Petre (October 2017). "New optimized method for low-temperature hydrothermal production of porous ceramics using diatomaceous earth". Ceramics International. 43 (15): 12572–12578. doi:10.1016/j.ceramint.2017.06.132. Archived from the original on 19 February 2022. Retrieved 8 April 2020.
- ^ Reka, Arianit; Anovski, Todor; Bogoevski, Slobodan; Pavlovski, Blagoj; Boškovski, Boško (29 December 2014). "Physical-chemical and mineralogical-petrographic examinations of diatomite from deposit near village of Rožden, Republic of Macedonia". Geologica Macedonica. 28 (2): 121–126. ISSN 1857-8586. Archived from the original on 18 April 2020. Retrieved 8 April 2020.
- ^ Reka, Arianit A.; Pavlovski, Blagoj; Ademi, Egzon; Jashari, Ahmed; Boev, Blazo; Boev, Ivan; Makreski, Petre (31 December 2019). "Effect Of Thermal Treatment Of Trepel At Temperature Range 800-1200˚C". Open Chemistry. 17 (1): 1235–1243. doi:10.1515/chem-2019-0132.
- ^ Auer, Antti (1991). "Qualitative Diatom Analysis as a Tool to Diagnose Drowning". The American Journal of Forensic Medicine and Pathology. 12 (3): 213–8. doi:10.1097/00000433-199109000-00009. PMID 1750392. S2CID 38370984.
- ^ Bradbury, J. (2004). "Nature's Nanotechnologists: Unveiling the Secrets of Diatoms". PLOS Biology. 2 (10): 1512–1515. doi:10.1371/journal.pbio.0020306. PMC 521728. PMID 15486572.

- ^ Drum, Ryan W.; Gordon, Richard (2003). "Star Trek replicators and diatom nanotechnology". Trends in Biotechnology. 21 (8): 325–8. doi:10.1016/S0167-7799(03)00169-0. PMID 12902165.
- ^ Johnson, R.C. (9 April 2009). "Diatoms could triple solar cell efficiency". EE Times. Archived from the original on 31 July 2012. Retrieved 13 April 2009.
- ^ Ramachandra, T. V.; Mahapatra, Durga Madhab; b, Karthick; Gordon, Richard (2009). "Milking Diatoms for Sustainable Energy: Biochemical Engineering versus Gasoline-Secreting Diatom Solar Panels". Industrial & Engineering Chemistry Research. 48 (19): 8769–88. doi:10.1021/ie900044j.
- ^ a b c Pierella Karlusich, Juan José; Ibarbalz, Federico M; Bowler, Chris (2020). "Exploration of marine phytoplankton: from their historical appreciation to the omics era". Journal of Plankton Research. 42: 595–612. doi:10.1093/plankt/fbaa049.
- ^ Dolan, John R. (1 August 2019). "Unmasking "The Eldest Son of The Father of Protozoology": Charles King". Protist. 170 (4): 374–384. doi:10.1016/j.protis.2019.07.002. ISSN 1434-4610. PMID 31479910. S2CID 201207778. Archived from the original on 10 August 2021. Retrieved 10 August 2021.
- ^ Darwin, Richard (1866). On the Origin of Species by Means of Natural Selection: Or the Preservation of Favoured Races in the Struggle for Life.
외부 링크
- Diatom EST 데이터베이스, Ecole Normale Supérieure
- 규조류 이미지를 포함하는 분류 데이터베이스인 Flanken*Net
- 캘리포니아 대학 고생물학 박물관, 규조류의 생활사와 생태
- 규조류: 베르겐 대학 유레카 유적지 '자연의 대리석'
- 규조생물사와 생태학, 학습과 교육을 위한 Microfossil 이미지 복구 및 순환(MIRACLE), University College London
- Diatom 페이지 2009년 10월 8일 로열 식물원 에든버러 웨이백 머신에서 아카이브
- 자연 3의 기하학과 패턴: 방사성 물질과 규조류 테스트의 구멍
- 몬터레이만 수족관 연구소, Diatom QuickFacts
- 조류 이미지 데이터베이스 필라델피아 자연과학 아카데미(ANSP)
- 필라델피아 자연과학 아카데미(ANSP)


