대뇌피질
Cerebral cortex| 대뇌피질 | |
|---|---|
 성체 마카크 원숭이의 뇌 조직 조각(마카카 물라타).대뇌피질은 짙은 보라색으로 묘사된 바깥쪽 층이다.출처 : BrainMaps.org | |
 피질의 골지 염색 뉴런 | |
| 세부 사항 | |
| 일부 | 대뇌 |
| 식별자 | |
| 라틴어 | 대뇌피질 |
| 메쉬 | D002540 |
| 신경명 | 39 |
| NeuroLex ID | birnlex_1494 |
| TA98 | A14.1.09.003 A14.1.09.301 |
| TA2 | 5527, 5528 |
| FMA | 61830 |
| 신경해부술의 해부학적 용어 | |
대뇌 [1]맨틀로도 알려진 대뇌 피질은 인간과 다른 포유동물에서 뇌의 대뇌 신경 조직의 바깥쪽 층이다.대뇌피질은 대부분 6층 신피질로 구성되어 있으며,[2] 10%만이 할당피질로 구성되어 있습니다.그것은 대뇌를 좌우 대뇌반구로 나누는 세로방향 틈에 의해 두 개의 피질로 분리된다.두 개의 반구는 뇌량에 의해 피질 아래에서 결합된다.대뇌피질은 중추신경계에서 [3]가장 큰 신경 통합 부위이다.그것은 주의력, 지각, 인식, 사고, 기억, 언어, 그리고 의식에 중요한 역할을 합니다.대뇌피질은 인지를 담당하는 뇌의 일부분이다.
대부분의 포유동물에서, 작은 뇌를 가진 작은 포유동물을 제외하고, 대뇌 피질은 접혀 있으며, 두개골의 제한된 부피에서 더 큰 표면적을 제공한다.뇌와 두개 부피를 최소화하는 것 외에도, 피질 접힘은 뇌 회로와 뇌 기능 [4]조직에 매우 중요하다.작은 뇌를 가진 포유류는 접힘이 없고 피질이 [5][6]부드럽다.
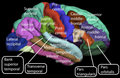
피질의 주름 또는 융기를 회(복수회), 홈을 설커스(복수회)라고 한다.이러한 표면 곡선은 태아 발달 과정에서 나타나며, 출생 후에도 회교화 과정을 통해 계속 성숙합니다.인간의 뇌에서는 대뇌피질의 대부분이 외부에서 보이지 않고 설치 [7]안에 묻힌다.주요 토막과 자이는 대뇌의 분열을 뇌의 엽으로 나타낸다.4대 엽은 전두엽, 두정엽, 후두엽, 후두엽, 측두엽이다.다른 엽은 변연엽이고, 섬피질은 종종 섬엽이라고 불립니다.
인간의 [3]대뇌피질에는 140억에서 160억개의 뉴런이 있다.이것들은 수평 피질층으로 구성되고 방사상으로 피질 기둥과 미니콜럼으로 구성됩니다.피질 영역은 운동 피질에서의 움직임과 시각 피질에서의 시각과 같은 특정한 기능을 가지고 있다.시각 피질은 후두엽에 위치해 있다.
구조.

대뇌피질은 대뇌반구 표면의 바깥쪽을 덮고 있으며 자리라고 불리는 봉우리들과 설치라고 불리는 홈으로 접혀 있다.인간의 뇌에서 그것은 2에서 3 또는 4밀리미터의 [8]두께이며, 뇌 [3]질량의 40%를 차지한다.대뇌피질의 90퍼센트는 6개의 대뇌신피질이고 나머지 10퍼센트는 [3]알로피질이다.피질에는 [3]140억에서 160억개의 뉴런이 있으며 이것들은 방사상으로 피질 기둥과 작은 반추로 구성되어 있습니다.[9][10] 피질의 수평으로 조직된 층에 있습니다.
신피질은 피질로 알려진 피질의 다른 영역으로 분리될 수 있으며 운동 피질과 시각 피질을 포함합니다.피질 표면의 약 3분의 2가 설지에 묻혀 있고 섬피질은 완전히 숨겨져 있다.피질은 회 윗부분에서 가장 두껍고 [11]구강 아랫부분에서 가장 얇다.
접다
대뇌피질은 신경조직의 넓은 표면적이 신경두개 경계 안에 들어갈 수 있도록 접혀 있다.사람에게 펼쳐질 때, 각 반구 피질의 총 표면적은 약 0.12 평방 미터(1.3 평방 피트)[12]이다.접힘은 뇌의 표면에서 안쪽으로 떨어져 있고, 세로 방향의 균열 내 각 반구의 안쪽 표면에도 있습니다.대부분의 포유류는 자이로 알려진 봉우리들과 sulci로 알려진 홈 또는 홈으로 복잡한 대뇌피질을 가지고 있다.몇몇 작은 설치류를 포함한 몇몇 작은 포유류는 [6]회전이 없는 매끄러운 뇌 표면을 가지고 있다.
로브
더 큰 sulci와 gyri는 대뇌 피질의 분열을 뇌의 [8]엽으로 표시합니다.전두엽, 두정엽, 측두엽, 후두엽의 4가지 주요 엽이 있습니다.섬피질은 종종 [13]섬엽으로 포함된다.변연엽은 각 반구의 안쪽 측면에 있는 피질의 테두리이며 종종 포함된다.[14]또한 묘사된 뇌의 세 개의 소엽이 있다: 근원소엽, 상두정소엽, 하두정소엽.
두께
포유류의 종의 경우, 더 큰 뇌는 단지 신체 크기뿐만 아니라 절대적인 관점에서 더 두꺼운 [15]피질을 갖는 경향이 있습니다.랫드와 같은 가장 작은 포유류는 신피질 두께가 약 0.5mm이고, 인간과 긴수염고래와 같이 가장 큰 뇌를 가진 포유류는 두께가 2-4mm이다.[3][8]뇌의 무게와 피질 [15]두께 사이에는 대략적인 로그 관계가 있다.뇌의 자기공명영상(MRI)은 인간 대뇌피질의 두께를 측정해 다른 측정과 연관짓는 것을 가능하게 한다.피질 부위의 두께는 다양하지만 일반적으로 감각 피질은 운동 [16]피질보다 얇다.한 연구는 피질 두께와 [17]지능 사이에 몇 가지 긍정적인 연관성을 발견했다.또 다른 연구는 편두통 환자의 체질 감각 피질이 더 두껍다는 것을 알아냈지만, 이것이 편두통 발작의 결과인지 아니면 [18][19]편두통의 원인인지는 알려지지 않았다.더 많은 환자를 사용한 최근의 연구는 [20]편두통 환자의 피질 두께에 변화가 없다고 보고했다.특정 부위의 접힘이 감소하면 6개 층이 아닌 4개 층이 있는 미세회로가 되는 대뇌피질의 유전적 질환은 어떤 경우에는 난독증과 [21]관련이 있는 것으로 보인다.
신피질층



신피질은 가장 바깥쪽 층 I에서 가장 안쪽 층 VI까지 I에서 VI까지 6개의 층으로 구성되어 있습니다.각 피질층은 다른 뉴런의 특징적인 분포와 다른 피질 및 피질하 영역과의 연결을 가진다.다른 피질 영역과 시상을 통한 간접 연결 사이에는 직접적인 연결이 있습니다.
피질층화의 가장 명확한 예 중 하나는 일차 시각 피질에 있는 Gennari 선이다.이것은 후두엽의 칼칼린 구근에서 육안으로 관찰할 수 있는 흰색 조직의 띠입니다.제나리 라인은 시상에서 시각 정보를 시각 피질의 IV 층으로 가져오는 축삭으로 구성되어 있습니다.
신경세포체와 피질내 축삭의 위치를 밝히기 위해 피질의 단면을 염색하는 것은 20세기 초 신경해부학자들이 다른 종의 피질 층 구조에 대한 상세한 설명을 만들 수 있게 했다.Korbinian Brodmann(1909)의 연구는 포유류의 신피질이 지속적으로 6개의 층으로 나뉘어져 있다는 것을 확립했다.
레이어 I
Layer I은 분자층이며 GABAergic Rosehip [22]뉴런을 포함한 소수의 산란 뉴런을 포함합니다.I층은 크게 피라미드 뉴런의 꼭대기 수상돌기와 수평방향 축삭,[4] 그리고 신경아교세포로 구성되어 있다.개발 중에, 이 층에는 카잘-레치우스[23] 세포와 아음경 입상층 세포가[24] 존재한다.또한, 몇몇 가시 돋친 성상 세포들이 이곳에서 발견될 수 있다.첨단에 대한 입력은 연상 학습과 [25]주의에 관여하는 대뇌 피질의 피드백 상호작용에 매우 중요한 것으로 생각된다.한때 I층에 대한 입력이 피질 [26]자체에서 왔다고 생각되었지만, 대뇌 피질 맨틀을 가로지르는 I층이 매트릭스 또는 [28]M형 시상[27] 세포로부터 상당한 입력을 받는다는 것이 실현되었습니다.
레이어 II
외부 입상층인 층 II는 작은 피라미드형 뉴런과 수많은 성상 뉴런을 포함합니다.
레이어 III
외부 피라미드층인 III층은 주로 작고 중간 크기의 피라미드형 뉴런과 수직 방향의 피질 내 축삭을 가진 비피라미드 뉴런을 포함한다. I층부터 III층은 반구간 피질 구심점의 주요 타깃이며 III층은 피질 유출물의 주요 공급원이다.
레이어 IV
내부 입상층인 레이어 IV는 다른 종류의 성상세포와 피라미드세포를 포함하고 있으며 시상피질구심뿐만 아니라 시상피질구심세포(코어형)[28]의 주요 타깃이다.레이어 IV 위의 레이어는 초립형 레이어(레이어 I-III)라고도 불리는 반면, 아래 레이어는 초립형 레이어(레이어 V 및 VI)라고 불립니다.
레이어 V
내부 피라미드층인 V층은 큰 피라미드형 뉴런을 포함하고 있다.이들로부터 축삭은 피질을 떠나 기저핵을 포함한 피질하 구조와 연결된다.전두엽의 1차 운동 피질에서, V층은 Betz 세포라고 불리는 거대한 피라미드 세포를 포함하고 있으며, 축삭은 내부 캡슐, 뇌간, 그리고 자발적인 운동 조절의 주요 경로인 피질척수관을 형성하는 척수를 통해 이동합니다.
레이어 VI
다형성 또는 다형성 층인 VI층은 몇 개의 큰 피라미드형 뉴런과 많은 작은 스핀들 모양의 피라미드형 및 다형성 뉴런을 포함합니다; VI층은 피질과 [29]시상 사이에 매우 정밀한 상호 연결을 확립하면서 시상으로 섬유질을 보냅니다.즉, 한 피질 기둥의 VI층 뉴런은 동일한 피질 기둥에 입력을 제공하는 시상 뉴런과 연결됩니다.이러한 연결은 자극적이면서도 억제적입니다.뉴런은 시상에 있는 뉴런에 흥분성 섬유를 보내고, 또한 시상에 있는 동일한 시상 뉴런 또는 그 [30]옆에 있는 시상 뉴런을 억제하는 망상핵에 부수적 섬유를 보냅니다.한 가지 이론은 억제 출력이 대뇌 피질에 대한 콜린 작동성 입력에 의해 감소하기 때문에, 이것은 뇌간에 조절 가능한 "렘니스칼 [30]입력의 릴레이를 위한 이득 제어"를 제공한다는 것입니다.
열
피질층은 단순히 다른 층 위에 쌓이는 것이 아니다; 다른 층과 피질의 모든 두께에 걸쳐 있는 뉴런 유형 사이에 특징적인 연결이 존재한다.이러한 피질 미세 회로는 피질 기둥과 미니콜럼으로 [31]분류됩니다.미니콜럼은 피질의 [32]기본적인 기능 단위라고 제안되어 왔다.1957년, 버논 마운트캐슬은 피질의 기능적 특성이 측면으로 인접한 지점들 사이에서 갑자기 변화한다는 것을 보여주었지만, 그것들은 표면과 수직인 방향으로 연속적이다.이후 연구는 시각 피질(Hubel and Wiesel, 1959),[33] 청각 피질 및 연상 피질에서 기능적으로 구별되는 피질 기둥의 존재에 대한 증거를 제공했다.
IV층이 없는 피질부위를 무과피질부위라고 한다.기본적인 층 IV만을 가진 피질 부위는 [34]입상 이상이라고 불린다.각 층 내의 정보처리는 2Hz의 느린 진동을 갖는 층 II/III와 10~15Hz의 빠른 [35]진동을 갖는 층 V와 다른 시간역학에 의해 결정된다.
피질의 종류
층상조직의 차이에 기초하여 대뇌피질은 두 가지 유형으로 분류할 수 있습니다. 즉, 6개의 세포층을 가진 신피질의 넓은 영역과 3개 또는 4개의 [2]층을 가진 훨씬 작은 영역입니다.
- 신피질은 또한 이소피질 또는 신팔륨으로 알려져 있으며 6개의 뚜렷한 층을 가진 성숙한 대뇌피질의 일부이다.신피질 영역의 예로는 입상 1차 운동 피질 및 줄무늬 1차 시각 피질이 있다.신피질에는 두 가지 서브타입이 있습니다진정한 등피질과 이등피질과 주변피질 영역 사이의 과도기 영역인 전등피질입니다
- 알로피질은 대뇌피질 중 3개 층으로 이루어진 부분이고, 3개의 피질층이 있는 고피질, 4개 또는 5개의 대피질, 그리고 알로피질과 인접한 과도기 영역인 주피질입니다.알로피질의 예로는 후각피질과 해마가 있다.
신피질과 알로피질 사이에는 2층, 3층, 4층이 합쳐진 평행피질이라고 불리는 과도기 영역이 있습니다.이 영역은 신피질의 전등피질과 알로피질의 근피질을 통합합니다.또한, 대뇌 피질은 4개의 엽으로 분류될 수 있습니다: 전두엽, 측두엽, 두정엽, 후두엽. 두개골의 위에 있는 뼈에서 이름이 붙여졌습니다.
혈액 공급 및 배수

대뇌피질로의 혈액공급은 뇌순환의 일부이다.대뇌동맥은 대뇌를 관류하는 혈액을 공급한다.이 동맥혈은 산소, 포도당, 그리고 다른 영양소를 피질로 운반합니다.뇌정맥은 탈산소화된 혈액과 이산화탄소를 포함한 대사 노폐물을 심장으로 배출한다.
대뇌피질을 공급하는 주요 동맥은 전대뇌동맥, 중대뇌동맥, 후대뇌동맥이다.전대뇌동맥은 전두엽의 대부분을 포함한 뇌의 앞부분을 공급한다.중뇌동맥은 두정엽, 측두엽, 후두엽의 일부를 공급한다.중뇌동맥은 두 갈래로 갈라져 왼쪽과 오른쪽 반구에 공급되고 거기서 더 갈라진다.후대뇌동맥은 후두엽에 공급한다.
윌리스 원은 대뇌와 대뇌피질의 혈액 공급을 다루는 주요 혈액계이다.

발전
대뇌피질의 태아 발달은 유전자와 [36]환경 사이의 상호작용에 영향을 받는 피질형성이라고 불리는 복잡하고 미세하게 조정된 과정이다.
신경관
대뇌 피질은 신경관의 [37][38]가장 앞부분인 전뇌 영역에서 발달한다.신경판은 접혔다가 닫히면서 신경관을 형성합니다.신경관 내부의 공동에서 심실계를 발달시키고, 벽의 신경상피세포에서 신경계의 뉴런과 글리아를 발달시킨다.신경판의 가장 앞부분인 프로센스뇌는 신경작용이 시작되기 전에 뚜렷하게 나타나며, 뇌반구와 후기의 [39]피질을 일으킨다.
피질뉴런 발달
피질 뉴런은 심실 옆에 있는 심실 영역 내에서 생성됩니다.처음에, 이 구역은 신경줄기세포를 포함하고 있는데, 신경줄기세포는 방사상아교세포-원조세포로 이행하며, 신경줄기세포는 분열하여 아교세포와 [40]뉴런을 생성한다.
반지름 글리아

대뇌 피질은 다른 종류의 세포를 발생시키는 이종 세포 집단으로 구성되어 있다.이 세포들의 대부분은 신피질의 다른 세포 유형을 형성하는 방사상 아교 이동에서 파생되며 신경 생성의 증가와 관련된 기간입니다.마찬가지로, 신경생성의 과정은 피질의 다른 층을 형성하기 위해 적층을 조절한다.이 과정 동안 세포 운명의 제한은 피질의 어떤 세포 타입을 낳는 초기 조상들과 표면층의 뉴런만을 낳는 후기 조상들로부터 시작된다.이 분화세포의 운명은 피질에 피질 내측지형을 형성하고 표면층에는 젊은 뉴런을 형성하고 더 깊은 층에는 늙은 뉴런을 형성합니다.또한 다른 피질층을 세밀하게 구별하기 위해 층상 뉴런을 S상 또는 G2상으로 정지시킨다.층상 분화는 생후까지 완전히 이루어지지 않는다. 왜냐하면 층상 뉴런은 발달하는 동안 외인성 신호와 [41]환경적 신호에 여전히 민감하기 때문이다.
피질을 구성하는 대부분의 세포는 방사상 글리아에서 국소적으로 파생되지만, 다른 영역에서 이동하는 뉴런의 하위 집합 집단이 있습니다.요골아교세포는 피라미드 모양이고 글루탐산염을 신경전달물질로 사용하는 뉴런을 발생시키지만, 이러한 이동세포는 성상 모양의 뉴런을 기여하고 GABA를 주요 신경전달물질로 사용한다.이러한 GABAergic 뉴런은 심실 하부를 통해 피질에 접선 방향으로 이동하는 내측 신경절 융기(MGE)의 전구 세포에 의해 생성됩니다.GABA 수용체가 발달하는 동안 흥분성을 띠기 때문에 이러한 GABA 작동성 뉴런의 이동은 특히 중요하다.이 들뜸은 주로 GABA 수용체를 통한 염화물 이온의 플럭스에 의해 구동되지만, 성인의 염화물 농도 변화는 시냅스 후 [42]뉴런을 과분극시키는 염화물의 내부 플럭스를 일으킨다.전구세포의 제1분할에서 생성된 글리아섬유는 심실존에서 외측, 음경 표면까지 피질의 두께에 걸쳐 방사상으로 배향되며, [43][44]심실존에서 바깥쪽으로 뉴런의 이동을 위한 발판을 제공한다.
태어날 때 피질 뉴런의 세포체에는 수상돌기가 거의 없고 축삭은 발달하지 않았습니다.생후 1년 동안 수상돌기는 급격히 증가하여 다른 뉴런과의 시냅스 연결을 10만 개까지 수용할 수 있습니다.축삭은 세포에서 [45]멀리 뻗어나가도록 발달할 수 있다.
비대칭 분할
전구 세포의 첫 번째 분할은 대칭이며, 이것은 각 유사분열 주기에서의 전구 세포의 총 수를 복제한다.그리고 나서, 일부 전구 세포는 비대칭으로 분열하기 시작하여, 심실 영역을 떠나 방사상 신경교 섬유를 따라 이동하는 하나의 후유전 세포와, 발달이 끝날 때까지 계속 분열하는 하나의 전구 세포를 생성하며, 이것이 신경교 세포 또는 후엽 세포로 분화되면.유사 분열의 G1 단계가 길어짐에 따라, 선택적 세포 주기 연장으로 보여지는 것에서, 새로 태어난 뉴런은 [46]피질의 더 얕은 층으로 이동한다.이동하는 딸세포는 대뇌피질의 [47]피라미드 세포가 된다.발달 과정은 수백 개의 유전자와 후생적 조절 [48]메커니즘에 의해 시간 순서와 조절된다.
계층 구성

성숙한 대뇌피질의 층상구조는 발달과정에서 형성된다.생성된 첫 번째 피라미드 뉴런은 재엘린을 생성하는 카잘-레치우스 뉴런과 함께 전판에서 심실 영역과 심실 영역 밖으로 이동한다.다음으로, 전판 중앙으로 이동하는 뉴런의 코호트는 이 과도층을 성숙한 신피질의 층 I이 되는 표면적 한계 영역과 피질판이라고 불리는 중간 층을 형성하는 서브 [49]플레이트로 나눕니다.이 세포들은 성숙한 피질의 깊은 층인 5, 6층을 형성할 것이다.나중에 태어난 뉴런은 심층 뉴런을 지나 피질판으로 방사상으로 이동해 상층(2~4개)이 된다.따라서 피질의 층은 안쪽에서 [50]바깥쪽으로 순서대로 형성됩니다.이러한 신경 발생의 내부 배열에 대한 유일한 예외는 설치류와는 대조적으로, 신경 발생은 피질 [51]발생의 전체 기간 내내 계속되는 영장류의 층 I에서 발생합니다.
피질 패턴 형성

일차 운동과 시각 피질을 포함하는 기능 피질 영역의 지도는 배아 발달 [53][54]초기에 섬유아세포 성장 인자 FGF8과 같은 분자 신호에 의해 조절되는 '프로토맵'[52]에서 유래한다.이 신호들은 부분적으로 전사인자 표현의 구배를 조절함으로써 피질 원기의 표면에 있는 피질 영역의 크기, 모양 및 위치를 피질 패턴화라고 불리는 과정을 통해 조절합니다.이러한 전사 인자의 예로는 유전자 EMX2 및 PAX6가 [55]있다.두 전사인자가 함께 반대되는 표현의 구배를 형성합니다.Pax6은 로스트랄 측극에서 높게 발현되고 Emx2는 미측극에서 높게 발현된다.적절한 개발을 위해서는 이 구배를 확립하는 것이 중요합니다.예를 들어, Pax6의 돌연변이는 Emx2의 발현 수준이 정상 발현 영역 밖으로 확장되도록 할 수 있으며, 이는 궁극적으로 시각 피질처럼 일반적으로 미간피질에서 파생된 영역의 확장을 초래할 수 있다.반대로 Emx2에서 돌연변이가 발생하면 Pax6 발현 영역이 확장되어 전두피질 영역과 운동피질 영역이 확대될 수 있다.따라서, 연구원들은 피질 옆에 있는 유사한 구배와 신호 중심이 이러한 전사 [42]인자의 지역적 표현에 기여할 수 있다고 믿는다.피질을 위한 매우 잘 연구된 두 가지 패턴닝 신호는 FGF와 레티노산을 포함한다.FGF가 발달하는 피질의 다른 영역에서 양성애자 압박을 받으면 피질 패턴화가 중단됩니다.구체적으로는 전극에서 Fgf8이 증가하면 Emx2가 하향조절되어 피질영역의 미도변동이 일어난다.이것은 궁극적으로 로스트랄 지역의 확장을 야기한다.따라서 Fgf8과 다른 FGF는 Emx2와 Pax6의 발현 조절에 역할을 하며 대뇌피질이 어떻게 [42]다른 기능에 특화될 수 있는지를 나타낸다.
피질 표면적의 급속한 팽창은 방사상 아교세포의 자가 재생량에 의해 조절되며, 부분적으로 FGF 및 노치 [56]유전자에 의해 조절된다.피질 신경 생성과 층 형성의 기간 동안, 많은 고등 포유동물들은 [57][58]대뇌 피질의 특징적인 주름들을 생성하는 회전화 과정을 시작합니다.Gyrification은 DNA 관련 단백질[59] Trnp1과 FGF 및 SH[60][61] 시그널링에 의해 조절된다.
진화
모든 다른 뇌 영역 중에서 대뇌 피질은 가장 큰 진화적 변이를 보이며 가장 [6]최근에 진화했다.예를 들어 심장의 조절과 호흡수와 같은 중요한 기능을 하는 수질의 고도로 보존된 회로와는 대조적으로, 대뇌 피질의 많은 영역은 생존을 위해 엄격히 필요하지 않습니다.따라서, 대뇌 피질의 진화는 새로운 기능 영역, 특히 [6]피질 외부에서 직접 입력을 받지 않는 연관 영역의 출현과 수정을 목격했다.
피질 [62]진화의 핵심 이론은 Rakic에 의해 최초로 제안된 방사 단위 가설과 관련된 원형 지도 가설에 구체화된다.이 이론은 줄기세포 수준에서 달성되는 새로운 방사 단위들의 추가에 의해 새로운 피질 영역이 형성된다고 말한다.프로토맵 가설은 각 피질 영역에 있는 세포와 분자 정체성과 뉴런의 특성이 원시 지도에서 방사상 신경교 세포로 알려진 피질 줄기세포에 의해 지정된다고 말한다.이 지도는 분비된 신호 단백질과 하류 전사인자에 [63][64][65]의해 제어된다.
기능.

접속
대뇌피질은 시상과 기저신경절과 같은 다양한 피질하구조와 연결되어 있으며, 그들에게 정보를 전달하고 구심성 연결을 통해 정보를 받습니다.대부분의 감각 정보는 시상을 통해 대뇌피질로 전달됩니다.그러나 후각 정보는 후구를 통해 후피피질(해체피질)로 전달된다.대부분의 연결은 피질하 영역이 아닌 피질의 한 영역에서 다른 영역으로의 연결이다; Braitenberg와 Schüz(1998)는 입력 섬유가 끝나는 피질하 수준에서 피질외 구심점에 의해 최대 20%의 시냅스가 공급되지만 다른 영역과 다른 층에 있는 시냅스가 공급된다고 주장한다.ge는 훨씬 [66]더 낮을 수 있습니다.
피질 영역
Korbinian Brodmann의 초기 발표에서 대뇌 피질 전체가 52개의 다른 영역으로 분할되었습니다.Brodmann 영역으로 알려진 이러한 영역은 세포 아키텍처를 기반으로 하지만 다양한 기능과 관련이 있습니다.예를 들어 1차 시각 피질인 Brodmann 영역 17을 들 수 있다.
보다 일반적인 용어로 피질은 일반적으로 감각, 운동 및 연상 영역의 세 부분으로 구성됩니다.
감각 영역


감각 영역은 감각으로부터 정보를 받고 처리하는 피질 영역입니다.시상으로부터 감각 입력을 받는 피질의 일부를 일차 감각 영역이라고 합니다.시각, 청각, 촉각은 각각 1차 시각 피질, 1차 청각 피질, 1차 체질 감각 피질에 의해 제공됩니다.일반적으로, 두 반구는 신체의 반대편(반구)으로부터 정보를 받습니다.예를 들어, 오른쪽 1차 체감각 피질은 왼쪽 팔다리에서 정보를 수신하고 오른쪽 시각 피질은 왼쪽 시야에서 정보를 수신합니다.피질에서 감각 지도의 구성은 지형 지도라고 알려진 것에 대응하는 감각 기관의 것을 반영한다.예를 들어, 1차 시각 피질의 인접점은 망막의 인접점에 대응합니다.이 지형도를 망막 지도라고 합니다.마찬가지로 1차 청각피질에는 강장지도가 있고 1차 감각피질에는 체질지도가 있다.이 몸의 마지막 지형도는 몸의 다른 부분의 크기가 신경의 상대적인 밀도를 반영하는 변형된 인간 표현인 체감각 호문쿨루스로 설명되었습니다.손가락 끝과 입술과 같이 감각 신경이 많이 발달한 부위는 미세한 감각을 처리하기 위해 더 많은 피질 영역을 필요로 합니다.
모터 영역
운동 영역은 피질의 양쪽 반구에 위치합니다.운동 영역은 자발적인 움직임의 제어, 특히 손이 수행하는 미세한 파편화된 움직임과 매우 밀접하게 관련되어 있습니다.모터 영역의 오른쪽 절반이 차체의 왼쪽을 제어합니다. 그 반대도 마찬가지입니다.
일반적으로 피질의 두 영역을 모터라고 합니다.
- 자발적인[citation needed] 움직임을 실행하는 일차 운동 피질
- 보조 운동 영역과 자발적인 [citation needed]움직임을 선택하는 전운동 피질.
또한 모터 기능은 다음과 같은 용도로 설명되었습니다.
대뇌피질 바로 밑에는 기저핵이라 불리는 회백질 피질하 덩어리가 서로 연결되어 있다.기저신경절은 대뇌피질의 중뇌와 운동영역의 실질 흑색으로부터 입력을 받아 이 두 위치에 신호를 보냅니다.그들은 모터 제어에 관여하고 있다.시상의 측면에서 발견됩니다.기저신경절의 주요 구성 요소는 미간핵, 창자핵, 글로부스 팔리더스, 실체 흑색핵, 핵낭, 그리고 시상하핵입니다.장막과 글로부스 팔리두스는 함께 렌즈 모양의 몸을 형성하기 때문에 총칭하여 렌즈 모양의 핵으로도 알려져 있다.장막과 미간핵은 줄무늬가 있는 [67][68]모습을 따서 총칭하여 선조체라고도 불린다.
연관 영역

연관 영역은 1차 영역에 속하지 않는 대뇌 피질의 일부입니다.그들은 세계에 대한 의미 있는 지각 경험을 만들고, 우리가 효과적으로 상호작용할 수 있도록 하며, 추상적인 사고와 언어를 지원하는 기능을 합니다.두정엽, 측두엽, 후두엽은 모두 피질의 후부에 위치하여 감각 정보와 메모리에 저장된 정보를 통합합니다.전두엽 또는 전전두연상복합체는 추상적 사고뿐만 아니라 행동과 움직임을 계획하는 데 관여한다.글로벌하게 어소시에이션지역은 분산 [69]네트워크로서 편성됩니다.각 네트워크는 피질의 넓은 간격 영역에 분산된 영역을 연결합니다.개별 네트워크는 서로 인접해 배치되어 복잡한 일련의 상호 얽힌 네트워크가 서로 인접해 있습니다.어소시에이션 네트워크의 특정 조직은 네트워크 [70]간의 상호작용, 계층적 관계 및 경쟁에 대한 증거와 함께 논의됩니다.
인간에게서 연상 네트워크는 언어 기능에 특히 중요하다.과거에는 언어능력이 언어표현을 위해 왼쪽 하전두회 영역인 BA44와 BA45에서, 언어영역을 위해 베르니케 영역인 BA22에서 국지화된다는 이론이 있었다.그러나 언어 표현과 수신의 과정은 전두엽, 기저신경절, 소뇌, [71]종아리 등 외측구 주위 구조뿐만 아니라 다른 영역에서도 일어나는 것으로 나타났다.
임상적 의의

알츠하이머병이나 라포라병과 같은 신경변성 질환은 대뇌피질의 [73]회백질 위축을 나타내는 지표로 나타난다.
중추신경계의 다른 질병에는 간질, 운동 장애, 언어 장애(갑상증)와 같은 신경학적 질환이 포함된다. 뇌전증
질병이나 트라우마로 인한 뇌손상은 전두엽 장애와 같은 특정 엽에 손상을 줄 수 있으며 관련 기능에 영향을 미칠 수 있습니다.감염으로부터 뇌를 보호하는 혈액-뇌 장벽이 손상되어 병원균에 침투할 수 있다.
발육 중인 태아는 선천적 기형이나 후발달의 문제를 일으킬 수 있는 다양한 환경적 요인에 민감하다.예를 들어 모성 알코올 섭취는 태아 알코올 스펙트럼 [74]장애를 일으킬 수 있습니다.신경 발달 장애를 일으킬 수 있는 다른 요인으로는 약물과 같은 독성 물질과 엑스레이에 의한 방사선 노출이 있다.감염은 또한 피질의 발달에도 영향을 미칠 수 있다.바이러스 감염은 환각증 없이 매끄러운 피질을 만드는 발열증의 원인 중 하나이다.
피질 자극 매핑이라고 불리는 전기 피질 조영술의 한 종류는 피질의 특정 부위의 기능을 위치시키기 위해 노출된 뇌에 전극을 직접 놓는 것을 포함하는 침습적 시술이다.수술 전 [75]매핑을 포함한 임상 및 치료 애플리케이션에 사용됩니다.
피질 장애와 관련된 유전자
소두증, 정신분열증, 간뇌증 [76]등 대뇌피질의 광범위한 유전적 장애를 일으킬 수 있는 유전자 돌연변이가 많이 있다.염색체 이상은 또한 연약한 X 증후군이나 레트 증후군과 같은 많은 신경 발달 장애를 일으킬 수 있다.
MCPH1은 소두증에 대한 코드이며, 이 코드와 ASPM의 장애는 소두증과 [76]관련이 있다.니브린을 코드하는 유전자 NBS1의 돌연변이는 소두증으로 [76]특징지어지는 니메겐 파괴 증후군을 일으킬 수 있다.
EMX2,[77] COL4A1의 돌연변이는 대뇌 반구의 큰 부분이 없는 것으로 특징지어지는 정신분열증과 [78]관련이 있다.
역사
1909년, Korbinian Brodmann은 세포 구조의 차이에 기초하여 신피질의 다른 영역을 구별하고 대뇌피질을 52개의 [79]영역으로 나누었다.
산티아고 라몬 이 카할의 학생인 라파엘 로렌테 데 노는 수상돌기와 [79]축삭의 분포를 바탕으로 40종 이상의 다른 피질 뉴런을 확인했다.
기타 동물
대뇌피질은 모든 척추동물의 전뇌에서 발견되는 층상 구조인 팔리움에서 유래한다.팔륨의 기본 형태는 유체로 채워진 심실을 둘러싼 원통형 층이다.원통 둘레에는 등쪽 팔리움, 안쪽 팔리움, 복측 팔리움, 측면 팔리움 등 4개의 구역이 있으며 각각 신피질, 해마, 편도체, 후각 피질과 상동하는 것으로 생각됩니다.
최근까지 무척추동물에서 대뇌피질에 해당하는 것은 발견되지 않았다.하지만, 2010년 Cell 저널에 발표된 연구는 유전자 발현 프로파일을 바탕으로, 대뇌 피질과 누더기벌레 Platynereis dumerili의 [80]버섯 몸체 사이의 강한 친화력을 보고했다.버섯의 몸은 학습과 기억력에 중요한 역할을 하는 것으로 알려진 많은 종류의 벌레와 절지동물의 뇌에 있는 구조입니다; 유전적인 증거는 공통적인 진화적 기원을 나타내며, 따라서 대뇌 피질의 초기 전구체의 기원이 선캄브리아 시대로 거슬러 올라간다는 것을 나타냅니다.
기타 이미지
인간 대뇌피질 측면

인간 대뇌 피질의 안쪽 표면


「 」를 참조해 주세요.
레퍼런스
- ^ "cerebral mantle". TheFreeDictionary.com.
- ^ a b Strominger, Norman L.; Demarest, Robert J.; Laemle, Lois B. (2012). "Cerebral Cortex". Noback's Human Nervous System, Seventh Edition. Humana Press. pp. 429–451. doi:10.1007/978-1-61779-779-8_25. ISBN 978-1-61779-778-1.
- ^ a b c d e f Saladin, Kenneth (2011). Human anatomy (3rd ed.). McGraw-Hill. pp. 416–422. ISBN 9780071222075.
- ^ a b Shipp, Stewart (2007-06-17). "Structure and function of the cerebral cortex". Current Biology. 17 (12): R443–9. doi:10.1016/j.cub.2007.03.044. PMC 1870400. PMID 17580069.
- ^ Fernández, V; Llinares-Benadero, C; Borrell, V (17 May 2016). "Cerebral cortex expansion and folding: what have we learned?". The EMBO Journal. 35 (10): 1021–44. doi:10.15252/embj.201593701. PMC 4868950. PMID 27056680.
- ^ a b c d Rakic, P (October 2009). "Evolution of the neocortex: a perspective from developmental biology". Nature Reviews Neuroscience. 10 (10): 724–35. doi:10.1038/nrn2719. PMC 2913577. PMID 19763105.
- ^ Principles of neural science (4th ed.). McGraw-Hill, Health Professions Division. 2000-01-05. ISBN 978-0838577011.
- ^ a b c Roberts, P (1992). Neuroanatomy (3rd ed.). Springer-Verlag. pp. 86–92. ISBN 9780387977775.
- ^ Lodato, Simona; Arlotta, Paola (2015-11-13). "Generating Neuronal Diversity in the Mammalian Cerebral Cortex". Annual Review of Cell and Developmental Biology. 31 (1): 699–720. doi:10.1146/annurev-cellbio-100814-125353. PMC 4778709. PMID 26359774.
Functional columns were first defined in the cortex by Mountcastle (1957), who proposed the columnar hypothesis, which states that the cortex is composed of discrete, modular columns of neurons, characterized by a consistent connectivity profile.
- ^ Ansen-Wilson, LJ; Lipinski, RJ (January 2017). "Gene-environment interactions in cortical interneuron development and dysfunction: A review of preclinical studies". Neurotoxicology. 58: 120–129. doi:10.1016/j.neuro.2016.12.002. PMC 5328258. PMID 27932026.
- ^ Carpenter (1985). Core text of neuroanatomy (3rd ed.). Williams & Wilkins. pp. 348–358. ISBN 978-0683014556.
- ^ Toro, Roberto; Perron, Michel; Pike, Bruce; Richer, Louis; Veillette, Suzanne; Pausova, Zdenka; Paus, Tomáš (2008-10-01). "Brain Size and Folding of the Human Cerebral Cortex". Cerebral Cortex. 18 (10): 2352–2357. doi:10.1093/cercor/bhm261. ISSN 1047-3211. PMID 18267953.
- ^ Nieuwenhuys, R (2012). The insular cortex: a review. Progress in Brain Research. Vol. 195. pp. 123–63. doi:10.1016/B978-0-444-53860-4.00007-6. PMID 22230626.
- ^ Tortora, G; Derrickson, B (2011). Principles of anatomy & physiology (13th. ed.). Wiley. p. 549. ISBN 9780470646083.
- ^ a b Nieuwenhuys R, Donkelaar HJ, Nicholson C (1998). The central nervous system of vertebrates, Volume 1. Springer. pp. 2011–2012. ISBN 978-3-540-56013-5.
- ^ Frithjof Kruggel; Martina K. Brückner; Thomas Arendt; Christopher J. Wiggins; D. Yves von Cramon (2003). "Analyzing the neocortical fine-structure". Medical Image Analysis. 7 (3): 251–264. doi:10.1016/S1361-8415(03)00006-9. hdl:11858/00-001M-0000-0010-9C60-3. PMID 12946467.
- ^ Katherine L. Narr; Roger P. Woods; Paul M. Thompson; Philip Szeszko; Dilbert Robinson; Teodora Dimtcheva; Mala Gurbani; Arthur W. Toga; Robert M. Bilder (2007). "Relationships between IQ and Regional Cortical Grey Matter Thickness in Healthy Adults". Cerebral Cortex. 17 (9): 2163–2171. doi:10.1093/cercor/bhl125. PMID 17118969.
- ^ Alexandre F.M. DaSilva; Cristina Granziera; Josh Snyder; Nouchine Hadjikhani (2007). "Thickening in the somatosensory cortex of patients with migraine". Neurology. 69 (21): 1990–1995. doi:10.1212/01.wnl.0000291618.32247.2d. PMC 3757544. PMID 18025393.
- ^ Catharine Paddock (2007-11-20). "Migraine Sufferers Have Thicker Brain Cortex". Medical News Today. Archived from the original on 2008-05-11.
- ^ Datte R, Detre JA, et al. (Oct 2011). "Absence of changes in cortical thickness in patients with migraine". Cephalalgia. 31 (14): 1452–8. doi:10.1177/0333102411421025. PMC 3512201. PMID 21911412.
- ^ Habib M (2000). "The neurological basis of developmental dyslexia: an overview and working hypothesis". Brain. 123 (12): 2373–99. doi:10.1093/brain/123.12.2373. PMID 11099442.
- ^ "Scientists identify a new kind of human brain cell". Allen Institute. 27 August 2018.
- ^ Meyer, Gundela; Goffinet, André M.; Fairén, Alfonso (1999). "Feature Article: What is a Cajal–Retzius cell? A Reassessment of a Classical Cell Type Based on Recent Observations in the Developing Neocortex". Cerebral Cortex. 9 (8): 765–775. doi:10.1093/cercor/9.8.765. PMID 10600995.
- ^ Judaš, Miloš; Pletikos, Mihovil (2010). "The discovery of the subpial granular layer in the human cerebral cortex". Translational Neuroscience. 1 (3): 255–260. doi:10.2478/v10134-010-0037-4. S2CID 143409890.
- ^ Gilbert CD, Sigman M (2007). "Brain states: top-down influences in sensory processing". Neuron. 54 (5): 677–96. doi:10.1016/j.neuron.2007.05.019. PMID 17553419.
- ^ Cauller L (1995). "Layer I of primary sensory neocortex: where top-down converges upon bottom-up". Behav Brain Res. 71 (1–2): 163–70. doi:10.1016/0166-4328(95)00032-1. PMID 8747184. S2CID 4015532.
- ^ Rubio-Garrido P, Pérez-de-Manzo F, Porrero C, Galazo MJ, Clascá F (2009). "Thalamic input to distal apical dendrites in neocortical layer 1 is massive and highly convergent". Cereb Cortex. 19 (10): 2380–95. doi:10.1093/cercor/bhn259. PMID 19188274.
- ^ a b Jones EG (1998). "Viewpoint: the core and matrix of thalamic organization". Neuroscience. 85 (2): 331–45. doi:10.1016/S0306-4522(97)00581-2. PMID 9622234. S2CID 17846130.
- ^ 크로이츠펠트, O. 1995대뇌피질입니다스프링거-벨라그.
- ^ a b Lam YW, Sherman SM (2010). "Functional Organization of the Somatosensory Cortical Layer 6 Feedback to the Thalamus". Cereb Cortex. 20 (1): 13–24. doi:10.1093/cercor/bhp077. PMC 2792186. PMID 19447861.
- ^ Suzuki, IK; Hirata, T (January 2013). "Neocortical neurogenesis is not really "neo": a new evolutionary model derived from a comparative study of chick pallial development" (PDF). Development, Growth & Differentiation. 55 (1): 173–87. doi:10.1111/dgd.12020. PMID 23230908. S2CID 36706690.
- ^ Mountcastle V (1997). "The columnar organization of the neocortex". Brain. 120 (4): 701–722. doi:10.1093/brain/120.4.701. PMID 9153131.
- ^ Hubel DH, Wiesel TN (October 1959). "Receptive fields of single neurones in the cat's striate cortex". The Journal of Physiology. 148 (3): 574–91. doi:10.1113/jphysiol.1959.sp006308. PMC 1363130. PMID 14403679.
- ^ S.M. 돔브로스키, C.C.힐게타그, 그리고 H. 바바.Wayback Machine에서 아카이브된 2008-08-29 Rhesus Monkey의 양적 아키텍처는 전전두피질계를 구별합니다.Cereb. Cortex 11: 975~988. "그것들은 부족하거나 기초적인 입상층 IV(입상층)만을 가지고 있다."
- ^ Sun W, Dan Y (2009). "Layer-specific network oscillation and spatiotemporal receptive field in the visual cortex". Proc Natl Acad Sci U S A. 106 (42): 17986–17991. Bibcode:2009PNAS..10617986S. doi:10.1073/pnas.0903962106. PMC 2764922. PMID 19805197.
- ^ Pletikos, Mihovil; Sousa, Andre MM; et al. (22 January 2014). "Temporal Specification and Bilaterality of Human Neocortical Topographic Gene Expression". Neuron. 81 (2): 321–332. doi:10.1016/j.neuron.2013.11.018. PMC 3931000. PMID 24373884.
- ^ Wolpert, Lewis (2015). Principles of development (Fifth ed.). UK: Oxford University Press. p. 533. ISBN 9780199678143.
- ^ Warren N, Caric D, Pratt T, Clausen JA, Asavaritikrai P, Mason JO, Hill RE, Price DJ (1999). "The transcription factor, Pax6, is required for cell proliferation and differentiation in the developing cerebral cortex". Cerebral Cortex. 9 (6): 627–35. doi:10.1093/cercor/9.6.627. PMID 10498281.
- ^ Larsen, W. J. 인간배아학 제3판 2001. 페이지 421-422 ISBN 0-443-06583-7
- ^ Stephen C. Noctor; Alexander C. Flint; Tamily A. Weissman; Ryan S. Dammerman & Arnold R. Kriegstein (2001). "Neurons derived from radial glial cells establish radial units in neocortex". Nature. 409 (6821): 714–720. Bibcode:2001Natur.409..714N. doi:10.1038/35055553. PMID 11217860. S2CID 3041502.
- ^ Sur, Mriganka; Leamey, Catherine A. (2001). "Development and Plasticity of Cortical Areas and Networks". Nature Reviews Neuroscience. 2 (4): 251–262. doi:10.1038/35067562. PMID 11283748. S2CID 893478.
- ^ a b c Sanes, Dan H.; Reh, Thomas A.; Harris, William A. (2012). Development of the Nervous System. Elsevier Inc. ISBN 978-0-12-374539-2.
- ^ Rakic, P (October 2009). "Evolution of the neocortex: a perspective from developmental biology". Nature Reviews Neuroscience. 10 (10): 724–35. doi:10.1038/nrn2719. PMC 2913577. PMID 19763105.
- ^ Rakic, P (November 1972). "Extrinsic cytological determinants of basket and stellate cell dendritic pattern in the cerebellar molecular layer". The Journal of Comparative Neurology. 146 (3): 335–54. doi:10.1002/cne.901460304. PMID 4628749. S2CID 31900267.
- ^ Gilbert, Scott (2006). Developmental biology (8th ed.). Sinauer Associates Publishers. pp. 394–395. ISBN 9780878932504.
- ^ Calegari, F; Haubensack W; Haffner C; Huttner WB (2005). "Selective lengthening of the cell cycle in the neurogenic subpopulation of neural progenitor cells during mouse brain development". The Journal of Neuroscience. 25 (28): 6533–8. doi:10.1523/jneurosci.0778-05.2005. PMC 6725437. PMID 16014714.
- ^ P. Rakic (1988). "Specification of cerebral cortical areas". Science. 241 (4862): 170–176. Bibcode:1988Sci...241..170R. doi:10.1126/science.3291116. PMID 3291116.
- ^ Hu, X.L.; Wang, Y.; Shen, Q. (2012). "Epigenetic control on cell fate choice in neural stem cells". Protein & Cell. 3 (4): 278–290. doi:10.1007/s13238-012-2916-6. PMC 4729703. PMID 22549586.
- ^ Kostović, Ivica (1990). "Developmental history of the transient subplate zone in the visual and somatosensory cortex of the macaque monkey and human brain". Journal of Comparative Neurology. 297 (3): 441–470. doi:10.1002/cne.902970309. PMID 2398142. S2CID 21371568.
- ^ Rakic, P (1 February 1974). "Neurons in rhesus monkey visual cortex: systematic relation between time of origin and eventual disposition". Science. 183 (4123): 425–7. Bibcode:1974Sci...183..425R. doi:10.1126/science.183.4123.425. PMID 4203022. S2CID 10881759.
- ^ Zecevic N, Rakic P (2001). "Development of layer I neurons in the primate cerebral cortex". The Journal of Neuroscience. 21 (15): 5607–19. doi:10.1523/JNEUROSCI.21-15-05607.2001. PMC 6762645. PMID 11466432.
- ^ Rakic, P (8 July 1988). "Specification of cerebral cortical areas". Science. 241 (4862): 170–6. Bibcode:1988Sci...241..170R. doi:10.1126/science.3291116. PMID 3291116.
- ^ Fukuchi-Shimogori, T; Grove, EA (2 November 2001). "Neocortex patterning by the secreted signaling molecule FGF8". Science. 294 (5544): 1071–4. Bibcode:2001Sci...294.1071F. doi:10.1126/science.1064252. PMID 11567107. S2CID 14807054.
- ^ Garel, S; Huffman, KJ; Rubenstein, JL (May 2003). "Molecular regionalization of the neocortex is disrupted in Fgf8 hypomorphic mutants". Development. 130 (9): 1903–14. doi:10.1242/dev.00416. PMID 12642494.
- ^ Bishop, KM; Goudreau, G; O'Leary, DD (14 April 2000). "Regulation of area identity in the mammalian neocortex by Emx2 and Pax6". Science. 288 (5464): 344–9. Bibcode:2000Sci...288..344B. doi:10.1126/science.288.5464.344. PMID 10764649.
- ^ Rash, BG; Lim, HD; Breunig, JJ; Vaccarino, FM (26 October 2011). "FGF signaling expands embryonic cortical surface area by regulating Notch-dependent neurogenesis". The Journal of Neuroscience. 31 (43): 15604–17. doi:10.1523/jneurosci.4439-11.2011. PMC 3235689. PMID 22031906.
- ^ Rajagopalan, V; Scott, J; Habas, PA; Kim, K; Corbett-Detig, J; Rousseau, F; Barkovich, AJ; Glenn, OA; Studholme, C (23 February 2011). "Local tissue growth patterns underlying normal fetal human brain gyrification quantified in utero". The Journal of Neuroscience. 31 (8): 2878–87. doi:10.1523/jneurosci.5458-10.2011. PMC 3093305. PMID 21414909.
- ^ Lui, Jan H.; Hansen, David V.; Kriegstein, Arnold R. (2011-07-08). "Development and evolution of the human neocortex". Cell. 146 (1): 18–36. doi:10.1016/j.cell.2011.06.030. ISSN 1097-4172. PMC 3610574. PMID 21729779.
- ^ Stahl, Ronny; Walcher, Tessa; De Juan Romero, Camino; Pilz, Gregor Alexander; Cappello, Silvia; Irmler, Martin; Sanz-Aquela, José Miguel; Beckers, Johannes; Blum, Robert (2013-04-25). "Trnp1 regulates expansion and folding of the mammalian cerebral cortex by control of radial glial fate". Cell. 153 (3): 535–549. doi:10.1016/j.cell.2013.03.027. ISSN 1097-4172. PMID 23622239.
- ^ Wang, Lei; Hou, Shirui; Han, Young-Goo (2016-05-23). "Hedgehog signaling promotes basal progenitor expansion and the growth and folding of the neocortex". Nature Neuroscience. 19 (7): 888–96. doi:10.1038/nn.4307. ISSN 1546-1726. PMC 4925239. PMID 27214567.
- ^ Rash, Brian G.; Tomasi, Simone; Lim, H. David; Suh, Carol Y.; Vaccarino, Flora M. (2013-06-26). "Cortical gyrification induced by fibroblast growth factor 2 in the mouse brain". The Journal of Neuroscience. 33 (26): 10802–10814. doi:10.1523/JNEUROSCI.3621-12.2013. ISSN 1529-2401. PMC 3693057. PMID 23804101.
- ^ Rakic, P (8 July 1988). "Specification of cerebral cortical areas". Science. 241 (4862): 170–6. Bibcode:1988Sci...241..170R. doi:10.1126/science.3291116. PMID 3291116.
- ^ Fukuchi-Shimogori, T; Grove, EA (2 November 2001). "Neocortex patterning by the secreted signaling molecule FGF8". Science. 294 (5544): 1071–4. Bibcode:2001Sci...294.1071F. doi:10.1126/science.1064252. PMID 11567107. S2CID 14807054.
- ^ Bishop, KM; Goudreau, G; O'Leary, DD (14 April 2000). "Regulation of area identity in the mammalian neocortex by Emx2 and Pax6". Science. 288 (5464): 344–9. Bibcode:2000Sci...288..344B. doi:10.1126/science.288.5464.344. PMID 10764649.
- ^ Grove, EA; Fukuchi-Shimogori, T (2003). "Generating the cerebral cortical area map". Annual Review of Neuroscience. 26: 355–80. doi:10.1146/annurev.neuro.26.041002.131137. PMID 14527269. S2CID 12282525.
- ^ Braitenberg, V와 Schüz, A 1998.「Cortex: 신경 접속의 통계와 기하학.완전 개정판 2탄"뉴욕: Springer-Verlag
- ^ 살라딘, 케네스해부학 및 생리학:형태와 기능의 통일성, 제5판뉴욕: 맥그로힐 컴퍼니즈, 2010.인쇄.
- ^ 돌랜드의 건강소비자를 위한 의학사전, 2008.
- ^ Yeo BT, Krienen FM, Sepulcre J, Sabuncu MR, Lashkari D, Hollinshead M, Roffman JL, Smoller JW, Zöllei L, Polimeni JR, Fischl B, Liu H, Buckner RL (2011). "The organization of the human cerebral cortex estimated by intrinsic functional connectivity". Journal of Neurophysiology. 106 (3): 1125–1165. doi:10.1152/jn.00338.2011. PMC 3174820. PMID 21653723.
- ^ Rupesh Kumar Srivastava; Jürgen Schmidhuber (2014). "Understanding Locally Competitive Networks". arXiv:1410.1165 [cs.NE].
- ^ Cathy J. Price (2000). "The anatomy of language: contributions from functional neuroimaging". Journal of Anatomy. 197 (3): 335–359. doi:10.1046/j.1469-7580.2000.19730335.x. PMC 1468137. PMID 11117622.
- ^ Kentar, Modar; Mann, Martina; Sahm, Felix; Olivares-Rivera, Arturo; Sanchez-Porras, Renan; Zerelles, Roland; Sakowitz, Oliver W.; Unterberg, Andreas W.; Santos, Edgar (2020-01-15). "Detection of spreading depolarizations in a middle cerebral artery occlusion model in swine". Acta Neurochirurgica. 162 (3): 581–592. doi:10.1007/s00701-019-04132-8. ISSN 0942-0940. PMID 31940093. S2CID 210196036.
- ^ Ortolano S, Vieitez I, et al. (2014). "Loss of cortical neurons underlies the neuropathology of Lafora disease". Mol Brain. 7: 7. doi:10.1186/1756-6606-7-7. PMC 3917365. PMID 24472629.
- ^ Mukherjee, Raja A.S.; Hollins, Sheila (2006). "Fetal Alcohol Spectrum Disorder: An Overview". Journal of the Royal Society of Medicine. 99 (6): 298–302. doi:10.1177/014107680609900616. PMC 1472723. PMID 16738372.
- ^ Tarapore, PE; et al. (August 2012). "Preoperative multimodal motor mapping: a comparison of magnetoencephalography imaging, navigated transcranial magnetic stimulation, and direct cortical stimulation". Journal of Neurosurgery. 117 (2): 354–62. doi:10.3171/2012.5.JNS112124. PMC 4060619. PMID 22702484.
- ^ a b c Walsh, Christopher A.; Mochida, Ganeshwaran H. (1 May 2004). "Genetic Basis of Developmental Malformations of the Cerebral Cortex". Archives of Neurology. 61 (5): 637–640. doi:10.1001/archneur.61.5.637. PMID 15148137.
- ^ "EMX2 empty spiracles homeobox 2 [Homo sapiens (human)] – Gene – NCBI". www.ncbi.nlm.nih.gov.
- ^ Smigiel, R; Cabala, M; Jakubiak, A; Kodera, H; Sasiadek, MJ; Matsumoto, N; Sasiadek, MM; Saitsu, H (April 2016). "Novel COL4A1 mutation in an infant with severe dysmorphic syndrome with schizencephaly, periventricular calcifications, and cataract resembling congenital infection". Birth Defects Research. Part A, Clinical and Molecular Teratology. 106 (4): 304–7. doi:10.1002/bdra.23488. PMID 26879631.
- ^ a b Principles of neural science. Kandel, Eric R. (5th ed.). New York. 2013. pp. 347–348. ISBN 9780071390118. OCLC 795553723.
{{cite book}}: CS1 유지보수: 기타 (링크) - ^ Tomer, R; Denes, AS; Tessmar-Raible, K; Arendt, D; Tomer R; Denes AS; Tessmar-Raible K; Arendt D (2010). "Profiling by image registration reveals common origin of annelid mushroom bodies and vertebrate pallium". Cell. 142 (5): 800–809. doi:10.1016/j.cell.2010.07.043. PMID 20813265. S2CID 917306.
외부 링크
- NeuroNames의 hier-20
- BrainMaps 프로젝트의 "대뇌 피질"을 포함하는 염색된 뇌 슬라이스 이미지
- "주요 시각 피질", 웹비전:일차 시각 피질의 구조와 기능에 대한 포괄적인 기사입니다.
- "기본 셀 유형", Webvision:원숭이 대뇌 피질의 기본 세포 유형 이미지.
- 대뇌 피질 - 세포 중심 데이터베이스



{kind=link}