푸타멘
Putamen| 푸타멘 | |
|---|---|
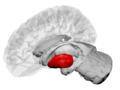
 뇌 내에 보이는 Putamen (빨간색) | |
 제3 심실의 중간 질량을 통한 뇌의 관상 부위. (상부에 Putamen 라벨이 붙어 있음) | |
| 세부 사항 | |
| 의 일부 | 도르살 선조체 |
| 식별자 | |
| 메슈 | D011699 |
| NeuroNames | 230 |
| NeuroEx ID | 버넥스_809 |
| TA98 | A14.1.09.507 |
| TA2 | 5566 |
| FMA | 61834 |
| 신경조영술의 해부학적 용어 | |
putamen(/pjutˈeɪmən/; 라틴어로 "너트껍질"이라는 뜻)은 전뇌(전뇌)의 기저부에 위치한 둥근 구조물이다. putamen과 caudate nuclear는 함께 등축조직을 형성한다. 또한 기저핵을 구성하는 구조 중 하나이다. 다양한 경로를 통해, 푸타멘은 대뇌피질의 많은 부위 외에도 실재성 니그라, 글로부스 팔리두스, 쇄골, 쇄골, 쇄골 등과 연결된다. Putamen의 주요 기능은 다양한 단계(예: 준비와 실행)에서 움직임을 조절하고 다양한 학습 유형에 영향을 미치는 것이다. GABA, 아세틸콜린, 엔케팔린 등을 채용해 기능을 수행한다. 파킨슨병 등 퇴행성 신경질환에도 푸타멘이 역할을 한다.
역사
"putamen"이라는 단어는 "putare"에서 "putare"를 의미하는 "putare"에서 "putare"에서 "putare"로, "putare"에서 "putaren"으로, "putare"에서 "putaren"으로, 또는 "생각하거나 고려하는 것"[1]을 뜻하는 라틴어에서
최근까지 대부분의 MRI 연구는 다양한 이유(예: 영상 해상도, 피타민 내 경색이나 출혈이 격리된 희귀성 등)로 인해 기저부 통풍에 광범위하게 초점을 맞췄다. 하지만, 기초적인 갱년기와 관련된 뇌 행동 관계에 대해 많은 연구가 행해져 왔다. 1970년대에 처음으로 단일 단위 녹음은 원숭이가 움직임과 관련된 완화 뉴런 활동을 감시하는 방식으로 이루어졌다. 이후 푸타멘에 대한 조사를 가능하게 하는 보다 광범위한 뉴런 추적, 자극, 영상 연구 방법(예: fMRI, DWI)이 개발되었다.
해부학

Putamen은 전뇌의 구조물이다. 그것은 입자핵과 함께 등줄기를 형성한다. 카우데이트와 푸타멘은 같은 종류의 뉴런과 회로를 포함하고 있다 – 많은 신경학자들은 등축 선조체를 하나의 구조로 간주하며, 큰 섬유조직인 내부 캡슐에 의해 두 부분으로 나뉘어져 중간을 통과한다. putamen은 globus palidus와 함께 렌즈핵을 구성한다. Putamen은 기초 갱년기의 가장 바깥쪽 부분이다. 이것들은 뇌 속의 핵으로 대뇌피질, 쇄골, 뇌계와 상호 연결된 집단이다. 기저성 갱년기(basal ganglia)는 등사 선조체, 실체성 흑색체, 측핵, 비탈사성 핵 등을 포함한다.
포유류에서 기초적인 갱년기는 운동 제어, 인지, 감정, 학습 및 도메인별 언어 지원뿐만 아니라 집행 기능에 중요한 도메인 일반 기능과 연관되어 있다. 기초적인 갱도는 쌍방향으로 위치하며, 회전과 카우달 사단을 가지고 있다. Putamen은 선조체의 일부로서 로스트랄 분할에 위치한다. 기초 강낭은 대뇌피질로부터 선조체로부터 입력을 받는다.
Putamen은 다음과 같은 구조와 상호 연결되어 있다.

이 설명은 초보적이며, 기본적인 확립된 피타멘 회로도 거의 소모하지 않는다. putaminal이 관여하는 cortico-subortico-cortico-cortico-cortico 회로는 조밀하고 복잡하며, 광범위한 차축, Dendritic, 화학, 분화, 효소 기질로 구성되어 있다. putamen의 출력은 출력 구조 전반에 걸쳐 매우 수목화되어 있으며, 피질 용출물은 putamen 내의 gyri와 위치에 따라 피질의 층 III-VI에서 발생한다. Putamen의 지형적 조직은 전위-후위 기능 및 섬광 구배, 횡-중간 기능 및 섬광 구배, 확산 단자 출력, 지역화된 인접 지역으로부터의 분리 단자, 원위 피질 재로부터 미세하게 분화된 단자를 결합한다.겉보기에 겹치는 듯한[citation needed] 느낌의 gions
카우다테핵
카우데이트는 대뇌피질로부터 입력을 받기 위해 피타멘과 함께 일한다. 집합적으로, 그들은 기저성 갱년기에 대한 "입구"로 간주될 수 있다. Putamen으로부터의 투영은 Caudolenticular 회색 다리를 통해 직접 Caudate에 도달한다. putamen과 caudate는 mercentia nigra와 공동으로 연결되어 있지만, caudate는 mercentia nigra pars reticulata에 더 밀도 있게 산출되는 반면 putamon은 more femfather를 internal globus pallidus에 보낸다.
스테레미아 니그라
실체형 니그라는 실체형 니그라 파스 콤팩트(SNPC)와 실체형 니그라 파스 레티쿨라타(SNPR)의 두 부분을 포함한다. SNpc는 putamen과 caudate로부터 입력을 얻어 정보를 되돌려 보낸다. SNpr은 또한 putamen과 caudate로부터 입력을 얻는다. 하지만, 그것은 머리와 눈의 움직임을 조절하기 위해 기저부 갱골 바깥으로 입력을 보낸다. SNpc는 움직임에 중요한 도파민을 생성한다. SNpc는 파킨슨병 때 변질되는 부분이다.[2]
글로부스팔리두스속
globus pallidus는 globus pars externa(GPe)와 globus pars interna(GPI)의 두 부분으로 구성되어 있다. 두 지역 모두 putamen과 caudate로부터 입력을 획득하고 subthalamic nuclear와 통신한다. 그러나 대부분 GPI는 가바에르그 억제 출력을 시상하부에 보낸다. GPI는 또한 자세 제어에 영향을 미치는 것으로 가정된 중간 두뇌의 일부에 투영을 보낸다.[2]
생리학
경로 유형
Putamen(및 일반적으로 선조체)은 코르티코-하부코르티코-코르티코 통신 루프를 허용하는 수많은 병렬 회로를 가지고 있다. 이것들은 대체로 직접, 간접, 초직접 경로로 설명되어 왔다. Putamen의 GABAergic 투영은 시상하부에 억제 효과를 가진다. 센트로메디아와 파라픽 핵에서 나오는 탈라믹 돌출부는 푸타멘에 흥분 효과를 가진다. 넓은 호혜적 연결성을 갖는 쇄골과 달리, 피질 투영과 피질 투영은 서로 다르기 때문에, 정보를 수신하는 것과는 반대로 정보를 송신한다. 피질 통신은 앞에서 설명한 바와 같이 다중 섬유 경로를 통해 이루어진다(즉, 다른 피질 구조물을 통해).
도파민
도파민은 피타멘에서 지배적인 역할을 하는 신경전달물질로, 대부분이 실재아 니그라에서 공급된다. 뉴런의 세포체(putamen 또는 caudate nuclei)가 작용 전위를 발사할 때 도파민은 전시냅스 단자에서 방출된다. putamen과 caudate nuclei로부터의 투영들은 실체형 니그라의 덴드라이트를 변형시키기 때문에, 도파민은 실체형 니그라에 영향을 미치며, 이것은 운동 계획에 영향을 미친다. 이와 같은 메커니즘은 약물 중독에 관련되어 있다. 시냅스 구분의 도파민 양과 시냅스 단자 이후의 도파민 결합 양을 조절하기 위해 프리시냅스 도파민성 뉴런이 과잉 도파민을 재흡수하는 기능을 한다.
기타 신경전달물질
Putamen은 또한 다른 신경전달물질의 변조에도 역할을 한다. GABA, 엔케팔린, 물질 P, 아세틸콜린을 방출한다. 세로토닌과 글루탐산염을 받는다.
기능: 운동 기술
Putamen은 많은 다른 구조들과 상호 연결되어 있고, 그것들과 함께 작동하여 많은 종류의 운동 행동에 영향을 준다. 여기에는 운동 계획, 학습 및 실행,[3] 운동 준비,[4] 운동 진폭 지정,[5] 운동 순서 지정 등이 포함된다.[6]
일부 신경학자는 푸타멘이 운동(예: 투렛 증후군)의 선택과 이전에 학습한 운동(예: 파킨슨병)의 "자동" 수행에도 역할을 한다는 가설을 세운다.[7]
한 연구에서 피아멘이 사지 운동을 조절한다는 것이 밝혀졌다. 이 연구의 목적은 영장류의 putamen에서의 특정 세포 활동이 사지 운동 방향 또는 근력 활동의 기저 패턴과 관련이 있는지를 결정하는 것이었다. 두 마리의 원숭이가 하중의 이동과 관련된 임무를 수행하도록 훈련을 받았다. 근육 활동과 움직임을 구분할 수 있도록 과제를 만들었다. Putamen의 뉴런은 작업과 작업 외부의 팔 움직임과 관련이 있는 경우에만 모니터링을 위해 선택되었다. 모니터링한 뉴런의 50%는 하중에 관계없이 이동 방향과 관련이 있는 것으로 나타났다.[8]
또 다른 연구는 13명의 인간에서 지역 뇌혈류량의 PET 지도를 이용하여 움직임의 범위와 속도를 조사하기 위해 수행되었다. 이동 작업은 조이스틱 제어 커서로 수행되었다. 통계적 테스트는 움직임의 범위와 움직임의 상관관계를 계산하기 위해 수행되었다. "이동 범위 증가는 쌍방향 기저뇌(BG; putamen and globus pallidus)와 입자뇌에서 rCBF의 평행 증가와 관련이 있다"는 것이 밝혀졌다. 이는 Putamen이 움직임에 영향을 줄 뿐만 아니라 Putamen이 작업을 수행하기 위해 다른 구조와 통합된다는 것을 보여준다.[9]
한 가지 연구는 기초적인 갱년기가 순차적인 움직임의 학습에 어떤 영향을 미치는지 구체적으로 조사하기 위해 수행되었다. 두 마리의 원숭이가 일련의 버튼을 순서대로 누르는 훈련을 받았다. 사용된 방법은 잘 학습된 과제 대 새로운 과제를 모니터링할 수 있도록 설계되었다. 무스시몰은 기저강골의 여러 부위에 주입되었으며, "전측각막과 퍼타멘에 주입한 후 새로운 시퀀스의 학습이 부족하게 되었으나 중후반 푸타멘은 아니었다"는 사실이 밝혀졌다. 이는 순차적 움직임 학습의 다양한 측면을 수행할 때 선조체의 다른 영역이 활용된다는 것을 보여준다.[10]
배움의 역할
많은 연구에서, Putamen이 많은 종류의 학습에서 역할을 한다는 것이 명백해졌다. 몇 가지 예는 다음과 같다.
강화 및 암묵적 학습
다양한 형태의 움직임과 함께, 푸타멘은 강화 학습과 암묵적 학습에도 영향을 미친다.[11]
강화학습은 환경과의 상호 작용과 케이터링 활동을 통해 성과를 극대화하고 있다. 암묵적 학습은 사람들이 정보에 노출되고 노출을 통해 지식을 습득하는 수동적인 과정이다. 정확한 메커니즘은 알려져 있지 않지만, 도파민과 톤으로 활동한 뉴런이 여기서 핵심적인 역할을 하고 있는 것은 분명하다. 톤 활성 뉴런은 자극의 전체 지속시간 동안 발화하며 초당 약 0.5-3회 임펄스로 발화하는 콜린내강이다. 페이식 뉴런은 반대편이며 움직임이 일어날 때 오직 행동 전위를 발사한다.[12]
카테고리 학습
한 특정 연구는 범주 학습을 연구하기 위해 뇌졸중으로 인한 기저성 갱년기(특히 putamen)에 초점 병변이 있는 환자를 사용했다. 이러한 유형의 환자를 사용할 때의 이점은 전두엽 피질에 대한 도파민성 투영이 손상되지 않을 가능성이 높다는 것이다. 또한 이러한 환자들에서는 병변이 특정 장소에서만 발생하기 때문에 특정한 뇌 구조를 기능적으로 연관시키는 것이 더 쉽다. 이 연구의 목적은 이러한 병변이 규칙 기반 및 정보 통합 작업 학습에 영향을 미치는지 여부를 결정하는 것이었다. 규칙 기반 작업은 작업 메모리에 따라 가설 검정을 통해 학습된다. 정보 통합 업무는 절차 기반 시스템을 따르는 결정 전 단계에서 두 출처의 정보가 통합될 때 정확도가 극대화되는 업무다.
실험에는 9명의 대조군 참가자와 함께 기저성 골지병변을 가진 7명의 참가자가 사용되었다. 그 경계가 영향을 받지 않았다는 것에 주목하는 것이 중요하다. 참가자들은 별도의 세션 동안 각 유형의 학습에 대해 테스트를 받았기 때문에 정보 프로세스가 서로 간섭하지 않을 것이다. 각 세션이 진행되는 동안 참가자들은 컴퓨터 화면 앞에 앉아 다양한 줄이 표시되었다. 이 선들은 네 가지 범주 중 하나에서 랜덤 표본을 추출하는 무작위화 기법을 사용하여 만들어졌다. 지배 기반 시험의 경우, 이 표본들은 이 네 가지 개별 범주에 속하는 다양한 길이와 방향의 선을 구성하기 위해 사용되었다. 자극이 표시된 후 피험자들에게 4개의 버튼 중 1개를 눌러 선이 어느 범주에 속하는지 표시하도록 했다. 정보 통합 과제에 대해서도 동일한 과정을 반복했으며, 범주 경계를 45° 회전시킨 것을 제외하고는 동일한 자극이 사용되었다. 이 회전을 통해 대상자는 라인이 어떤 범주에 속하는지 결정하기 전에 라인에 대한 정량적 정보를 통합하게 된다.
실험군의 피험자는 규칙 기반 작업을 수행하면서 장애가 발생했지만 정보 통합 작업은 그렇지 않은 것으로 나타났다. 통계 테스트 후, 뇌가 규칙 기반 학습 과제를 해결하기 위해 정보 통합 기법을 사용하기 시작했다는 가설도 나왔다. 규칙 기반 작업은 뇌의 가설 검증 시스템을 사용하므로 뇌의 가설 검증 시스템이 손상/약화되었다는 결론을 내릴 수 있다. 애도와 일하는 기억은 이 시스템의 일부인 것으로 알려져 있다. 따라서, Putamen이 범주 학습, 시스템 간의 경쟁, 규칙 기반 업무에서의 피드백 처리에 관여하고 있으며, (작업 기억 및 임원 직무와 관련된) 전전면적 영역의 처리에 관여하고 있는 것으로 확인되었다. 이제 기초 학문과 교미만이 카테고리 학습에 영향을 미치는 것은 아니라고 알려져 있다.[13]
"혐오 회로"에서의 역할
잠정적인 연구들은 푸타멘이 뇌의 소위 "혐오 회로"에 역할을 할 수 있다는 것을 시사했다. 최근 한 연구는 런던에서 유니버시티 칼리지 런던의 세포와 발달 생물학과에 의해 수행되었다. fMRI는 환자들이 그들이 싫어하는 사람들과 "중립적인" 사람들의 사진을 보는 동안 행해졌다. 실험 중 사진마다 '증오 점수'가 기록됐다. 뇌의 하위 문학적 영역에서의 활동은 "혐오 회로"가 피타멘과 인슐라를 포함한다는 것을 암시했다. "푸타멘은 경멸과 혐오에 대한 인식에 역할을 하며, 행동을 취하기 위해 동원되는 운동 시스템의 일부일 수도 있다"는 이론이 나왔다. 또한 혐오회로의 활동량은 사람이 선언하는 혐오량과 상관관계가 있는 것으로 밝혀져 악성 범죄에 대한 법적 함의가 있을 수 있다.[14]
트랜스젠더 개인의 역할
피아멘은 일반적인 시스젠더 남성의 피아멘에 비해 남성에서 여성 트랜스젠더 개인에서 상당히 많은 양의 회백질을 가지고 있는 것으로 밝혀졌다. 이것은 아마도 뇌 구성의 근본적인 차이가 트랜스 여성과 시스젠더 남성 사이에 존재할 수도 있고 없을 수도 있다는 것을 암시할 수 있다.[15]
병리학
파킨슨병
Putamen의 기능을 발견한 후, 신경학자들에게 Putamen과 다른 기저성 갱년기의 부위는 파킨슨병이나 뉴런의 퇴화를 수반하는 다른 질병에 중요한 역할을 한다는 것이 명백해졌다.[16]
파킨슨병은 골수 니그라파스 콤팩타에서 도파민성 뉴런의 느리고 꾸준한 손실이다. 파킨슨병에서 푸타멘의 입력과 출력이 실체성 니그라와 글로부스 팔리두스와 상호 연결되기 때문에 푸타멘은 핵심적인 역할을 한다. 파킨슨병에서는 내부 글로부스 팔리두스로 가는 직접 경로에서의 활동이 감소하고 외부 글로부스 팔리두스로 가는 간접 경로에서의 활동이 증가한다. 파킨슨병 환자들은 운동계획 수립에 어려움을 겪는다는 점도 지적됐다.
기타 질병 및 장애
다음과 같은 질병과 장애는 푸타멘과 연관되어 있다.
- 알츠하이머병의[17] 인지력 저하
- 헌팅턴병
- 윌슨병
- 레위몸을 동반한 치매
- 피티코바살 퇴화
- 투렛 증후군
- 정신분열증
- 우울증
- 주의력결핍과잉행동장애[18]
- 안디카
- 강박-강박 장애[19][20]
- 케르니테루스
- 기타 불안장애[20]
다른 동물들에서는
인간의 파타멘은 다른 동물들과 구조와 기능이 비교적 유사하다. 따라서 푸타멘에 대한 많은 연구가 인간뿐만 아니라 동물(원숭이, 쥐, 고양이 등)에 대해서도 행해져 왔다. 그러나 종간 변동은 포유류에서 실제로 관찰되며 백색 물질 putaminal 연결성에 대해 문서화되었다. 변화는 주로 구조 연결 패턴과 관련이 있는 반면, 단조로운 조직 원칙은 유지된다. 1980년대부터 현재까지 영장류 연구는 고차 인식에 관계된 피질 영역은 주로 다른 뉴런을 putamen의 대부분에 보내는 반면, 영장류의 나머지 구조는 주로 센서-모터 기능을 제공하며 일차적 및 유연성과 밀접하게 상호 연결되어 있다는 것을 규명했다.자동차 보급 지역
추가 이미지

전두엽을 통한 뇌의 관상동맥.

오른쪽 대뇌 반구의 수평 부분.

브레인

인간 뇌전면(코론) 섹션

Patamen을 보여주는 MRI 영상의 수평 슬라이스. 기저핵(caudate nucleus and globus pallidus)의 다른 핵도 볼 수 있다.

푸타멘

푸타멘
다른 아문 구조와 함께 Putamen

Coronal T1 MRI 영상에서 녹색으로 강조 표시된 Putamen

시상 T1 MRI 영상에서 녹색으로 강조 표시된 Putamen

횡단 T1 MRI 영상에서 녹색으로 강조 표시된 Putamen



_section_description_2.JPG)






참고 항목
참조
- ^ "Latin Definitions for: Putare (Latin Search) - Latin Dictionary and Grammar Resources - Latdict".
- ^ a b Alexander GE; Crutcher MD (July 1990). "Functional architecture of basal ganglia circuits: neural substrates of parallel processing". Trends in Neurosciences. 13 (7): 266–71. doi:10.1016/0166-2236(90)90107-L. PMID 1695401. S2CID 3990601.
- ^ DeLong MR; Alexander GE; Georgopoulos AP; Crutcher MD; Mitchell SJ; Richardson RT (1984). "Role of basal ganglia in limb movements". Human Neurobiology. 2 (4): 235–44. PMID 6715208.
- ^ Alexander GE; Crutcher MD (July 1990). "Preparation for movement: neural representations of intended direction in three motor areas of the monkey". Journal of Neurophysiology. 64 (1): 133–50. doi:10.1152/jn.1990.64.1.133. PMID 2388061.
- ^ Delong MR; Georgopoulos AP; Crutcher MD; Mitchell SJ; Richardson RT; Alexander GE (1984). "Functional organization of the basal ganglia: contributions of single-cell recording studies". Ciba Found. Symp. Novartis Foundation Symposia. 107: 64–82. doi:10.1002/9780470720882.ch5. ISBN 9780470720882. PMID 6389041.
- ^ Marchand WR; Lee JN; Thatcher JW; Hsu EW; Rashkin E; Suchy Y; Chelune G; Starr J; Barbera SS (June 11, 2008). "Putamen coactivation during motor task execution". NeuroReport. 19 (9): 957–960. doi:10.1097/WNR.0b013e328302c873. PMID 18521000. S2CID 11087809.
- ^ Griffiths PD; Perry RH; Crossman AR (March 14, 1994). "A detailed anatomical analysis of neurotransmitter receptors in the putamen and caudate in Parkinson's disease and Alzheimer's disease". Neuroscience Letters. 169 (1–2): 68–72. doi:10.1016/0304-3940(94)90358-1. PMID 8047295. S2CID 24324548.
- ^ Crutcher MD; DeLong MR (1984). "Single cell studies of the primate putamen. II. Relations to direction of movement and pattern of muscular activity". Exp. Brain Res. 53 (2): 244–58. doi:10.1007/bf00238154. PMID 6705862. S2CID 22320601.
- ^ Turner RS; Desmurget M; Grethe J; Crutcher MD; Grafton ST (December 2003). "Motor subcircuits mediating the control of movement extent and speed". Journal of Neurophysiology. 90 (6): 3958–66. doi:10.1152/jn.00323.2003. PMID 12954606. S2CID 7012970.
- ^ Miyachi S; Hikosaka O; Miyashita K; Kárádi Z; Rand MK (June 1997). "Differential roles of monkey striatum in learning of sequential hand movement". Exp. Brain Res. 115 (1): 1–5. doi:10.1007/PL00005669. PMID 9224828. S2CID 13541431.
- ^ Packard MG; Knowlton BJ (2002). "Learning and memory functions of the Basal Ganglia". Annu Rev Neurosci. 25 (1): 563–93. doi:10.1146/annurev.neuro.25.112701.142937. PMID 12052921.
- ^ Yamada H; Matsumoto N; Kimura M (April 7, 2004). "Tonically active neurons in the primate caudate nucleus and putamen differentially encode instructed motivational outcomes of action". Journal of Neuroscience. 24 (14): 3500–10. doi:10.1523/JNEUROSCI.0068-04.2004. PMC 6729748. PMID 15071097.
- ^ Ell SW; Marchant NL; Ivry RB (2006). "Focal putamen lesions impair learning in rule-based, but not information-integration categorization tasks". Neuropsychologia. 44 (10): 1737–51. doi:10.1016/j.neuropsychologia.2006.03.018. PMID 16635498. S2CID 16312162.
- ^ Zeki S; Romaya JP (2008). Lauwereyns, Jan (ed.). "Neural Correlates of Hate". PLOS ONE. 3 (10): e3556. doi:10.1371/journal.pone.0003556. PMC 2569212. PMID 18958169.
- ^ Luders E; Sánchez FJ; Gaser C; et al. (July 2009). "Regional gray matter variation in male-to-female transsexualism". NeuroImage. 46 (4): 904–7. doi:10.1016/j.neuroimage.2009.03.048. PMC 2754583. PMID 19341803.
- ^ DeLong MR; Wichmann T (January 2007). "Circuits and circuit disorders of the basal ganglia". Arch. Neurol. 64 (1): 20–4. doi:10.1001/archneur.64.1.20. PMID 17210805.
- ^ de Jong LW; van der Hiele K; Veer IM; Houwing JJ; Westendorp RG; Bollen EL; de Bruin PW; Middelkoop HA; van Buchem MA; van der Grond J (December 2008). "Strongly reduced volumes of putamen and thalamus in Alzheimer's disease: an MRI study". Brain. 131 (12): 3277–85. doi:10.1093/brain/awn278. PMC 2639208. PMID 19022861.
- ^ Martin H. Teicher; Carl M. Anderson; Ann Polcari; Carol A. Glod; Luis C. Maas; Perry F. Renshaw (2000). "Functional deficits in basal ganglia of children with attention-deficit/hyperactivity disorder shown with functional magnetic resonance imaging relaxometry". Nature Medicine. 6 (12): 470–3. doi:10.1038/74737. PMID 10742158. S2CID 32093789.
- ^ Radua, Joaquim; Mataix-Cols, David (November 2009). "Voxel-wise meta-analysis of grey matter changes in obsessive–compulsive disorder". British Journal of Psychiatry. 195 (5): 393–402. doi:10.1192/bjp.bp.108.055046. PMID 19880927.
- ^ a b Radua, Joaquim; van den Heuvel, Odile A.; Surguladze, Simon; Mataix-Cols, David (5 July 2010). "Meta-analytical comparison of voxel-based morphometry studies in obsessive-compulsive disorder vs other anxiety disorders". Archives of General Psychiatry. 67 (7): 701–711. doi:10.1001/archgenpsychiatry.2010.70. PMID 20603451.
외부 링크
| 위키미디어 커먼즈에는 푸타멘과 관련된 미디어가 있다. |
- BrainMaps 프로젝트의 "putamen"을 포함하는 얼룩진 뇌 조각 이미지
- "Anatomy diagram: 13048.000-2". Roche Lexicon - illustrated navigator. Elsevier. Archived from the original on 2012-07-22.
- 다이어그램(uni-tuebingen.de
- Atlas 이미지: eye_38, University of Michigan Health System – "아래의 시각 경로"



{kind=link}