아크로칸토사우루스
Acrocanthosaurus| 아크로칸토사우루스 | |
|---|---|
.jpg) | |
| 노스캐롤라이나 자연과학 박물관의 마운트 골격(NCSM 14345). | |
| 과학적 분류 | |
| 도메인: | 진핵생물 |
| 킹덤: | 애니멀리아 |
| 문: | 초르다타 |
| 클레이드: | 공룡 |
| 클레이드: | 사우리샤 |
| 클레이드: | 테로포다 |
| 가족: | †카르카로돈토사우루스과 |
| 속: | †아크로칸토사우루스 스토볼 & 랭스턴, 1950 |
| 유형종 | |
| †아크로칸토사우루스 아토켄시스 스토볼 & 랭스턴, 1950 | |
| 동의어 | |
| |
아크로칸토사우루스(/ˌæ크로 ʊˌ크 æ ɔː르 ə스/AK-roh-KANth ə-SOR-ə스; '고가시 도마뱀')는 백악기 전기 압티안 시대에 현재의 북아메리카에 존재했던 카르카로돈토사우루스과 공룡의 한 속으로, 1억 1천만년 전부터 1억 1천만년 전까지 존재했습니다.대부분의 공룡과 마찬가지로, 아크로칸토사우루스는 A. atokensis라는 단 하나의 종만을 포함하고 있습니다.비록 아크로칸토사우루스의 것으로 추정되는 치아, 경골, 척추뼈가 메릴랜드만큼 동쪽으로 멀리서 발견되고 있기는 하지만, 이 화석의 잔해는 주로 미국의 오클라호마, 텍사스, 와이오밍 주에서 발견되고 있으며, 이는 대륙 전체의 범위를 암시하고 있습니다.
아크로칸토사우루스는 이족보행의 포식자였습니다.이름에서 알 수 있듯이, 그것은 척추뼈의 많은 부분에 있는 높은 신경 가시로 가장 잘 알려져 있는데, 이것은 아마도 동물의 목, 등, 그리고 엉덩이에 있는 근육의 능선을 지탱했을 것입니다.아크로칸토사우루스는 가장 큰 수각류 중 하나로, 알려진 가장 큰 표본은 길이가 11-11.5 미터(36-38 피트)에 달하고 무게가 약 4.4-6.6 미터(4.9-7.3 톤)에 달했습니다.텍사스에서 발견된 대형 수각류 발자국은 아크로칸토사우루스가 만들었을지도 모르지만, 해골 유해와 직접적인 연관성은 없습니다.
최근의 발견들은 그것의 뇌 구조와 앞다리 기능에 초점을 맞춘 전문적인 연구를 가능하게 하면서 그것의 해부학의 많은 세부사항들을 설명했습니다.아크로칸토사우루스는 그 생태계에서 가장 큰 수각류였고 용각류, 조류, 강직류를 잡아먹는 최상위 포식자였을 것입니다.
검색 및 이름 지정
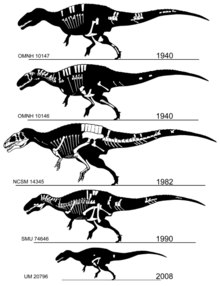
아크로칸토사우루스의 이름은 그리스의 ɑκρɑ/아크라("높다"), ɑκɑνθɑ/아칸타("가시" 또는 "가시"), σɑʊρος/사우로스("도마뱀")에서 따온 것입니다.원래 표본이 발견된 오클라호마의 아토카 카운티(Atoka County)의 이름을 따서 명명된 종(A. atokensis)이 하나 있습니다.그 이름은 1950년에 미국의 고생물학자 J. 윌리스 스토볼과 완 랭스턴 주니어에 의해 만들어졌습니다.[2]랭스턴(Langston)은 1947년 발표되지 않은 석사 논문에서 이 속과 종의 이름을 "Acracanthus atokaensis"로 제안했지만,[3][4] 공식적인 출판을 위해 이름을 "Acrocanthosaurus atokensis"로 변경했습니다.
1940년대 초에 발견되어 1950년에 동시에 기술된 완모식과 파라타입(OMNH 10146과 OMNH 10147)은 오클라호마의 앤틀러스 층에서 나온 두 개의 부분 골격과 두개골 물질로 구성되어 있습니다.[2]1990년대에 두 개의 훨씬 더 완전한 표본이 묘사되었습니다.첫 번째(SMU 74646)는 텍사스의 트윈 마운틴 층과 현재 포트워스 과학 및 역사 박물관 소장품의 일부에서 회수된 두개골의 대부분이 없어진 부분적인 골격입니다.[5]더 완벽한 골격 (NCSM 14345, 별명은 "프란")은 사우스 다코타의 블랙 힐스 연구소에서 준비한 세피스 홀과 시드 러브에 의해 오클라호마의 앤틀러스 층에서 발견되었고, 현재 롤리에 있는 노스 캐롤라이나 자연 과학 박물관에 소장되어 있습니다.표본은 가장 크고 유일하게 알려진 완전한 두개골과 앞다리를 포함합니다.OMNH 10147의 골격 요소는 NCSM 14345의 비교 가능한 뼈와 거의 동일한 크기이며, 이는 완모식체와 SMU 74646은 상당히 작은 크기임을 나타냅니다.[6]
아크로칸토사우루스의 클로블리 층에서의 존재는 2012년에 또 다른 부분 골격인 UM 20796에 대한 설명과 함께 확립되었습니다.두 개의 척추뼈, 부분 치골, 대퇴골, 부분 치골, 그리고 조각들로 구성된 이 표본은 어린 동물을 나타냅니다.와이오밍 중북부의 빅혼 분지에 있는 뼈층에서 발견되었으며, 사우로포세이돈의 어깨뼈 근처에서 발견되었습니다.이 층에서 발견된 여러 조각으로 된 수각류의 잔해들 또한 클로버리 층에서 유일하게 큰 수각류일 수도 있는 아크로칸토사우루스의 것일 수도 있습니다.[7]
아크로칸토사우루스는 오클라호마, 텍사스, 와이오밍 이외의 지역에서는 덜 완전한 유적으로 알려져 있을지도 모릅니다.애리조나 남부에서 온 치아가 이 속으로 언급되었고,[8] 같은 지역의 용각류 뼈에서 일치하는 치아 자국이 발견되었습니다.[9]메릴랜드의 아룬델 지층에서 나온 몇몇 이빨들은 아크로칸토사우루스의 이빨과 거의 동일한 것으로 묘사되어 왔으며, 아크로칸토사우루스의 동쪽 대표적인 이빨일 수도 있습니다.[10]미국 서부 전역의 다양한 지질층에서 발견된 많은 다른 치아와 뼈들도 아크로칸토사우루스로 언급되고 있지만, 이들 중 대부분은 잘못된 것으로 밝혀졌습니다.[11] 그러나 클로블리 층의 화석에 대한 이 평가에는 약간의 이견이 있습니다.[7]
묘사

아크로칸토사우루스는 알려진 가장 큰 수각류 중 하나로, 알려진 가장 큰 표본(NCSM 14345)을 기준으로 추정되는 두개골 길이는 1.23~1.29m (4.0~4.2 피트), 몸 길이는 11~11.5m (36~38 피트)였습니다.[12][6][13]연구원들은 다양한 기술을 바탕으로 이 표본의 체질량 추정치를 4.4에서 6.6 미터 톤(4.9에서 7.3 쇼트 톤)으로 산출했습니다.[12][14][15]
스컬
아크로칸토사우루스의 두개골은 다른 알로사우루스류들처럼 길고 낮고 좁았습니다.안구 소켓(안와쪽 펜스트라) 앞에 있는 체중 감소 개구부는 두개골 길이의 4분의 1 이상, 높이의 3분의 2 이상으로 상당히 컸습니다.상악골(위턱뼈)의 바깥쪽 표면과 주둥이의 지붕에 있는 코뼈의 윗면은 기가노토사우루스나 카르카로돈토사우루스처럼 거친 질감은 아니었습니다.코뼈에서 길고 낮은 융기선이 생겨났고, 콧구멍에서 눈까지 주둥이의 양쪽을 따라 달려갔고, 그곳에서 눈물뼈까지 이어졌습니다.[6]이것은 모든 알로사우로이드의 특징입니다.[16]알로사우루스와 달리 눈 앞의 눈물뼈에는 눈에 띄는 볏이 없었습니다.홍반뼈와 후안와뼈는 카르카로돈토사우루스과와 관련이 없는 아벨리사우루스과에서 볼 수 있듯이 눈 위에 두꺼운 눈썹을 형성했습니다.위턱 양쪽에는 19개의 구부러진 톱니가 늘어서 있었지만 아래턱에 대한 치아 수는 발표되지 않았습니다.아크로칸토사우루스의 이빨은 카르카로돈토사우루스의 이빨보다 넓었고 카르카로돈토사우루스과를 특징짓는 주름진 질감을 가지고 있지 않았습니다.치골(치아를 가진 아래턱뼈)은 지가노토사우루스처럼 앞쪽 가장자리가 네모지고 얕아진 반면, 뒤쪽 턱의 나머지 부분은 매우 깊어졌습니다.아크로칸토사우루스와 기가노토사우루스는 두개골과의 관절 아래 아래 아래턱의 사각형 뼈 바깥 표면에 두꺼운 수평 능선을 공유하고 있었습니다.[6]
두개후골격

아크로칸토사우루스의 가장 눈에 띄는 특징은 목, 등, 엉덩이 그리고 꼬리의 척추뼈 위에 위치한 높은 신경 척추들의 줄이었습니다. 이 척추뼈들은 그들이 뻗은 척추뼈의 높이의 2.5배 이상이 될 수도 있었습니다.[2]다른 공룡들도 등에 높은 가시를 가지고 있었는데, 때로는 아크로칸토사우루스의 가시보다 훨씬 높았습니다.예를 들어, 아프리카 속 스피노사우루스는 척추뼈의 몸통보다 약 11배 더 큰 거의 2m (6.6피트) 높이의 척추를 가지고 있었습니다.[17]아크로칸토사우루스의 아래쪽 가시들은 현대 들소들처럼 강력한 근육들에 대한 애착을 가졌고, 아마도 등 아래로 높고 두꺼운 능선을 형성했을 것입니다.[2]척추의 기능은 알려지지 않았지만 의사소통, 지방 저장, 근육 또는 온도 조절에 관련되었을 수 있습니다.모든 경추(목)와 등(등) 척추는 측면에 두드러진 함몰부(플뢰로쿨)를 가지고 있었고, 꼬리 척추는 더 작은 함몰부를 가지고 있었습니다.이것은 알로사우루스보다 카르카로돈토사우루스과와 더 유사합니다.[5]
척추뼈 외에도, 아크로칸토사우루스는 전형적인 알로사우루스 골격을 가지고 있었습니다.아크로칸토사우루스는 머리와 몸의 균형을 잡아주는 길고 무거운 꼬리를 가진 이족보행의 동물로 엉덩이 위로 무게중심을 유지했습니다.앞다리는 알로사우루스보다 상대적으로 짧고 튼튼했지만, 그 외에는 비슷했습니다.빠르게 달리는 많은 작은 공룡들과는 달리, 이 공룡의 대퇴골은 경골과 중족골보다 길었고,[5][6] 이는 아크로칸토사우루스가 빨리 뛰지 못했음을 암시합니다.[18]놀랄 것도 없이, 아크로칸토사우루스의 뒷다리 뼈는 더 작은 친척 알로사우루스보다 비례적으로 더 튼튼했습니다.그것의 발은 각각 네 개의 숫자를 가지고 있었지만, 수각류의 경우 전형적인 것처럼, 첫 번째 숫자는 다른 숫자보다 훨씬 작았고 땅과 접촉하지 않았습니다.[5][6]
분류와 체계

아크로칸토사우루스는 테타누라목에 속하는 알로사우로이데아과에 속합니다.이 슈퍼패밀리는 코뼈와 코뼈 위에 있는 눈물뼈와 목뼈에 있는 큰 신경 가시, 그리고 다른 특징들 중에서도 특징적입니다.[16]원래 알로사우루스과에 알로사우루스과로 분류되었는데,[2] 2000년 말에 이루어진 연구에서도 이를 뒷받침하고 있습니다.[6]대부분의 연구들은 이 동물이 근연종인 카르카로돈토사우루스과의 일원임을 밝혀냈습니다.[16][19][20]
발견 당시 아크로칸토사우루스와 다른 대형 수각류들은 단편적인 유적으로만 알려져 있었으며, 이 속의 분류는 매우 다양했습니다.J. 윌리스 스토볼(J. Willis Stovall)과 완 랭스턴(Wann Langston) 주니어(Jr.)는 처음에 이 종을 알로사우루스과에 해당하는 안트로데미대("Antrodemidae")로 분류했지만, 1956년 알프레드 셔우드 로머(Alfred Sherwood Romer)에 의해 분류학적 폐바구니 메갈로사우루스과로 옮겨졌습니다.[21]다른 저자들에게는 척추뼈에 있는 긴 가시가 스피노사우루스와 관련이 있음을 시사합니다.[22][23]아크로칸토사우루스를 스피노사우루스과(spinosaurid)로 해석하는 이러한 해석은 1980년대까지 지속되었으며,[24] 당시의 반기술 공룡 서적에서도 반복되었습니다.[25][26]
영국 백악기 전기의 큰 가시 척추뼈는 한때 아크로칸토사우루스의 것과 매우 비슷하다고 여겨졌고,[27] 1988년 그레고리 S. 폴은 이 척추뼈를 알티스피낙스속의 두 번째 종으로 명명했습니다.[28]이 뼈들은 원래 이빨로만 알려진 영국의 수각류 동물인 알티스피낙스에 할당되었고, 이 할당은 적어도 한 명의 작가가 알티스피낙스 그 자체가 아크로칸토사우루스의 동의어라고 제안하게 만들었습니다.[27]이 척추뼈들은 나중에 아크로칸토사우루스와 알티스피낙스와는 별개로 새로운 속인 베클레스스피낙스에 할당되었습니다.[29]일부 연구자들에 의해서도 알로사우루스과로 간주되었고, 아직도 가끔 알로사우루스과에 포함됩니다.[30][31]


아크로칸토사우루스를 포함한 대부분의 분류학적 분석들은 그것이 보통 아프리카의 카르카로돈토사우루스와 남아메리카의 기가노토사우루스와 비교하여 기초적인 위치에 있는 카르카로돈토사우루스과임을 발견했습니다.[16][5][32]아프리카에서도 서식하는 똑같이 기초적인 에오카르카리아의 자매 분류군으로 여겨져 왔습니다.영국에서 발견된 네오베네이터는 종종 훨씬 더 기초적인 카르카로돈토사우루스과 또는 네오베네이터과라고 불리는 자매군의 기초적인 구성원으로 여겨집니다.[18][20]이는 이 과가 유럽에서 기원한 후 남대륙(당시 초대륙 곤드와나로 통합)으로 분산되었음을 시사합니다.만약 아크로칸토사우루스가 카르카로돈토사우루스과였다면, 북아메리카에도 분산이 일어났을 것입니다.[5]알려진 모든 카로돈토사우루스과는 백악기 초중반에 살았습니다.[16]2013년 Novas et al. 이후의 다음 클래도그램은 아크로칸토사우루스의 카르카로돈토사우루스과의 위치를 보여줍니다.[33]2011년 올리버 라우헛(Oliver Rauhut)은 탄자니아의 쥐라기 시대의 텐다구루 층에서 발견된 수각류 공룡의 새로운 속을 베테루프리스티사우루스(Veterupristisaurus)라고 이름 지었고, 이 공룡이 아크로칸토사우루스의 자매 분류군이라는 것을 밝혀내 카르카로돈토사우루스과의 지위를 더욱 뒷받침했습니다.
| |||||||||||||||||||||||||||||||||||||||||||||||||
고생물학
성장장수

완모식표본인 OMNH 10146과 NCSM 14345의 뼈 특징으로 볼 때, 아크로칸토사우루스가 완전히 자라는데 적어도 12년이 걸렸을 것으로 추정됩니다.뼈의 재생과 수강의 성장 과정에서 해리스 라인이 일부 소실되었기 때문에 이 수치는 훨씬 더 높았을 것입니다.만약 이 선들을 설명한다면, 아크로칸토사우루스가 성숙하는 데는 18-24년이 필요했습니다.[34]
무는 힘
사카모토 등(2022)은 아크로칸토사우루스의 무는 힘을 다른 공룡 33마리와 비교하여 연구하였다.그 결과에 따르면, 턱 앞 부분의 물린 힘은 8,266 뉴톤이었고, 뒤로 물린 힘은 16,894 뉴톤으로 추정되었습니다.[35]
앞다리함수
다른 대부분의 비조류 수각류들처럼 아크로칸토사우루스 앞다리는 땅과 접촉하지 않았고 이동에 사용되지 않았습니다. 대신 포식적인 역할을 했습니다.완전한 앞다리 (NCSM 14345)의 발견은 아크로칸토사우루스의 앞다리의 기능과 운동 범위를 최초로 분석할 수 있게 해주었습니다.[36]이 연구는 관절이 탈구되지 않고 얼마나 멀리 움직일 수 있는지를 알아내기 위해 다른 뼈들과 관절을 형성했을 뼈 표면을 조사했습니다.많은 관절에서 뼈가 정확히 맞지 않아 많은 살아있는 공룡들에게서 볼 수 있듯이 관절에 상당한 양의 연골이 존재함을 알 수 있습니다.다른 발견들 중에서, 그 연구는, 휴식을 취하는 자세에서, 상완골이 약간 뒤로 기울어져 있고, 팔꿈치가 구부러져 있고, 발톱이 안쪽을 향하고 있는 상태에서, 앞다리가 어깨에 매달려 있었을 것이라고 암시했습니다.[36]아크로칸토사우루스의 어깨는 인간의 어깨에 비해 움직임의 범위가 제한적이었습니다.팔은 완전한 원을 그리며 회전할 수는 없지만 수직에서 109° 뒤로 후퇴(뒤로 스윙)하여 상완골이 실제로 약간 위쪽으로 각도를 맞출 수 있습니다.보호(앞으로 흔들기)는 수직을 지나 24°로 제한되었습니다.암이 아래로 흔들릴 때 수직 위치에 도달할 수 없었지만 수평 위로 9°까지 납땜(위로 흔들림)할 수 있었습니다.팔꿈치의 움직임 또한 인간에 비해 제한적이었는데, 총 움직임 범위는 57°에 불과했습니다.팔은 완전히 뻗을 수도 없고(곧게 펴질 수도 없고), 상완골이 팔뚝과 직각을 이룰 수도 없을 정도로 멀리 휘어질 수도 없었습니다.사람의 팔뚝에서처럼 발음이나 수포(뒤틀림)의 가능성이 없도록 반지름과 척골(팔뚝뼈)이 서로 맞물려 있습니다.[36]

손목뼈(손목뼈) 중 어느 것도 정확하게 맞지 않아 손목에 연골이 다량 존재해 뻣뻣했을 것으로 추정됩니다.모든 숫자는 손목에 닿을 때까지 과도하게 (뒤로) 뻗을 수 있었습니다.구부릴 때, 가운데 자리는 첫 번째 자리 쪽으로 모이고, 세 번째 자리는 안쪽으로 꼬입니다.손의 첫 번째 손가락에는 가장 큰 집게발이 달려있는데, 이 집게발은 손의 아래쪽을 향해 뒤로 굽어지도록 영구적으로 구부려졌습니다.마찬가지로, 가운데 집게발은 영구적으로 구부려진 것일 수도 있고, 세 번째 집게발은 또한 가장 작은 집게발은 구부리고 펴는 것이 가능했습니다.[36]앞다리의 관절의 움직임 범위를 알아낸 후, 이 연구는 아크로칸토사우루스의 포식성 습성에 대해 가설을 세우기 시작했습니다.앞다리는 동물의 목을 긁을 수도 없을 정도로 앞으로 멀리 뛸 수 없었습니다.따라서, 그들은 처음에 먹이를 잡는 데 사용되었을 가능성이 낮고 아크로칸토사우루스는 아마도 사냥할 때 입으로 이끌었을 것입니다.반면에, 앞다리는 몸 쪽으로 매우 강하게 후퇴할 수 있었습니다.일단 먹이가 턱에 잡히면, 근육이 많이 발달한 앞다리가 후퇴하여 먹이를 몸에 단단히 고정시키고 탈출을 막을 수도 있습니다.먹이 동물이 도망치려고 시도했을 때, 그것은 앞 두 자리 숫자의 영구적으로 구부러진 발톱에만 더 달려 있었을 것입니다.극도의 과도한 손가락 확장성은 아크로칸토사우루스가 탈구에 대한 두려움 없이 고군분투하는 먹이를 잡을 수 있게 해주는 적응이었을지도 모릅니다.일단 먹이가 몸에 걸려들면, 아크로칸토사우루스가 턱으로 먹이를 급파했을지도 모릅니다.또 다른 가능성은 아크로칸토사우루스가 발톱으로 큰 가시를 찢고 앞다리를 반복적으로 뒤로 젖히면서 먹이를 턱에 물고 있었을 가능성입니다.[36]가능성이 낮은 다른 이론들은 용각류의 옆구리를 움켜쥐고 더 작은 키의 용각류를 쓰러뜨리기 위해 매달릴 수 있는 앞다리의 범위를 제시하고 있지만, 이것은 아크로칸토사우루스가 비슷한 구조의 다른 수각류에 비해 다리 구조가 다소 강했기 때문일 가능성은 높지 않습니다.
뇌와 내이 구조

2005년, 과학자들은 홀로타입 뇌 케이스(OMNH 10146) 내의 공간을 분석하기 위해 컴퓨터 단층 촬영(CT 스캔)을 사용하여 아크로칸토사우루스의 두개골 구멍의 내막(복제물)을 재구성했습니다.살면서, 이 공간의 많은 부분은 뇌수막과 뇌척수액, 그리고 뇌 자체로 채워졌을 것입니다.그러나 뇌와 뇌 신경의 일반적인 특징은 내생동물로부터 결정될 수 있고 내생동물이 만들어진 다른 수각류와 비교될 수 있습니다.뇌는 많은 수각류 동물들과 비슷하지만, 알로사우로이드의 뇌와 가장 유사합니다.알로사우루스나 신랍토르의 뇌보다는 카르카로돈토사우루스와 기가노토사우루스의 뇌와 가장 유사하며, 아크로칸토사우루스가 카르카로돈토사우루스과라는 가설을 뒷받침합니다.[37]
뇌는 약간의 S자형으로 대뇌 반구가 크게 확장되지 않고 새보다는 악어에 더 가까웠습니다.이것은 비코엘루로사우루스류 수각류 뇌의 전반적인 보수성과 일치합니다.아크로칸토사우루스는 후각이 뛰어나다는 것을 나타내는 크고 구근모양의 후각 전구를 가지고 있었습니다.균형을 조절하는 귀의 반고리관을 재구성해 보면 머리가 수평 아래 25° 각도로 잡혀 있었다는 것을 알 수 있습니다.이것은 동물이 경계 자세를 취할 때처럼 측면 반고리관이 지면과 평행하도록 내막을 향하도록 함으로써 결정되었습니다.[37]
발자국 가능성

텍사스 중부의 글렌 로즈 층에는 커다란 세 개의 발가락을 가진 수각류 발자국을 포함한 많은 공룡 발자국들이 보존되어 있습니다.[38]이 선로들 중 가장 유명한 것은 공룡 계곡 주립 공원의 팔룩시 강을 따라 발견되었는데, 이 구간은 현재 뉴욕시에 있는 미국 자연사 박물관에서 전시 중이지만,[39] 그 주 주변의 몇몇 다른 장소들은 문헌에 기술되어 있습니다.[40][41]어떤 동물이 발자국을 만들었는지 말하는 것은 불가능한데, 그 이유는 어떤 화석 뼈도 선로와 연관되어 있지 않기 때문입니다.하지만, 과학자들은 오랫동안 이 발자국들이 아크로칸토사우루스의 것일 가능성이 높다고 여겨왔습니다.[42]2001년의 한 연구는 글렌 로즈 발자국을 다양한 큰 수각류의 발과 비교했지만 특정한 속에 자신 있게 할당할 수 없었습니다.하지만, 이 연구는 이 흔적들이 아크로칸토사우루스에게 예상되는 크기와 모양의 범위 안에 있다는 것에 주목했습니다.글렌 로즈 층은 지리적 위치와 지질학적 시대 모두에서 앤틀러스 층과 트윈 마운틴 층에 가깝고, 이 층에서 알려진 유일한 큰 수각류는 아크로칸토사우루스뿐이기 때문에, 이 연구는 아크로칸토사우루스가 흔적을 만들었을 가능성이 가장 높다고 결론 내렸습니다.[43]
뉴욕시에 전시 중인 유명한 글렌 로즈 트랙웨이에는 용각류 공룡 12마리와 같은 방향으로 이동한 여러 개체의 수각류 발자국이 포함되어 있습니다.이 수각류 발자국은 용각류 발자국의 꼭대기에서 발견되기도 하는데, 이는 이 발자국들이 나중에 형성되었다는 것을 보여줍니다.이것은 아크로칸토사우루스의 작은 무리가 용각류 무리를 스토킹하고 있다는 증거로 제시되었습니다.[39]흥미롭고 그럴듯하지만, 이 가설은 증명하기 어렵고 다른 설명들도 존재합니다.예를 들어, 몇몇 외딴 수각류들은 용각류가 지나간 후 다른 시간대에 같은 방향으로 이동하여 무리가 먹이를 스토킹하는 모습을 연출했을 수도 있습니다.집단적으로 움직였을 수도 있고 그렇지 않았을 수도 있는 용각류의 "떼"들에게도 마찬가지라고 할 수 있습니다.[44]용각류 중 한 마리의 길을 건너는 지점에서 수각류의 궤도 중 한 곳에 발자국이 없어 공격의 증거로 지목되고 있습니다.[45]하지만, 다른 과학자들은 용각류가 큰 포식자가 옆구리에 매달려 있다면 예상되는 것처럼 걸음걸이를 바꾸지 않았기 때문에 이 해석의 타당성에 의문을 제기합니다.[44]
병리학

아크로칸토사우루스 아토켄시스 완모식표본의 두개골은 두개골에 가벼운 이물감을 보여줍니다.11번째 척추의 신경 척추는 골절되어 치유되었고, 3번째 꼬리 척추의 신경 척추는 특이한 갈고리와 같은 구조를 가지고 있었습니다.[46]
고생물학

확실한 아크로칸토사우루스 화석은 텍사스 북부의 트윈 마운틴 층, 오클라호마 남부의 앤틀러스 층, 와이오밍 중북부의 클로버 층 그리고 메릴랜드의 아룬델 층에서 발견되었습니다.이 지질학적 형성은 방사선 측정법으로 연대가 측정되지 않았지만, 과학자들은 그들의 나이를 추정하기 위해 생물 계층학을 사용해왔습니다.암모나이트 분류군의 변화에 근거하여 백악기 전기의 압티아 단계와 알비아 단계 사이의 경계는 아크로칸토사우루스의 발자국이 있을 수 있는 텍사스의 글렌 로즈 층 안에 위치해 있으며 트윈 마운틴 층 바로 위에 놓여 있습니다.이것은 트윈 마운틴 지층이 1억 2천 2백만 년 전부터 1억 1천 2백만 년 전까지 지속된 압티안 지층 안에 있다는 것을 나타냅니다.[47]앤틀러스 층은 데이노니쿠스와 테논토사우루스의 화석을 포함하고 있는데, 클로버리 층에서도 발견된 공룡의 두 속으로, 이 공룡들은 방사선 측정상으로 압티아와 알비아 단계까지 거슬러 올라가며, 앤틀러스의 시기가 비슷하다는 것을 암시합니다.[48]따라서 아크로칸토사우루스는 1억 2천 5백만 년에서 1억 년 전 사이에 존재했을 가능성이 가장 높습니다.[16]
이 시기에 트윈 마운틴과 앤틀러스 지층에 보존된 지역은 얕은 내해로 배수되는 큰 홍수터였습니다.몇 백만 년 후, 이 바다는 북쪽으로 확장되어 서부 내륙 해로가 되었고 거의 백악기 후기 전체 동안 북아메리카를 둘로 나누었습니다.글렌 로즈 층은 해안 환경을 나타내며, 아크로칸토사우루스의 흔적이 고대 해안선을 따라 갯벌에 보존되어 있을 가능성이 있습니다.아크로칸토사우루스가 큰 포식자였기 때문에, 아크로칸토사우루스는 넓은 서식 범위를 가지고 있었고 그 지역의 많은 다른 환경에서 살았을 것으로 예상됩니다.[43]잠재적인 먹잇감 동물로는 Astrodon과[49] 같은 용각류나 심지어 거대한 Sauroposeidon과 같은 거대한 조류뿐만 아니라 [50]테논토사우루스와 같은 대형 조류도 있습니다.[51]더 작은 수각류인 데이노니쿠스도 이 지역을 배회했지만 길이가 3m에 달해 아크로칸토사우루스에게 최소한의 경쟁이나 심지어 먹이를 제공했을 가능성이 높습니다.[48]
참고문헌
- ^ Liddell, Henry George; Robert Scott (1980). Greek–English Lexicon, Abridged Edition. Oxford: Oxford University Press. ISBN 978-0-19-910207-5.
- ^ a b c d e Stovall, J. Willis; Langston, Wann. (1950). "Acrocanthosaurus atokensis, a new genus and species of Lower Cretaceous Theropoda from Oklahoma". American Midland Naturalist. 43 (3): 696–728. doi:10.2307/2421859. JSTOR 2421859.
- ^ Langston, Wann R. (1947). A new genus and species of Cretaceous theropod dinosaur from the Trinity of Atoka County, Oklahoma. Unpublished M.S. thesis. University of Oklahoma.
- ^ Czaplewski, Nicholas J.; Cifelli, Richard L.; Langston, Wann R. Jr. (1994). "Catalog of type and figured fossil vertebrates. Oklahoma Museum of Natural History". Oklahoma Geological Survey Special Publication. 94 (1): 1–35.
- ^ a b c d e f Harris, Jerald D. (1998). "A reanalysis of Acrocanthosaurus atokensis, its phylogenetic status, and paleobiological implications, based on a new specimen from Texas". New Mexico Museum of Natural History and Science Bulletin. 13: 1–75.
- ^ a b c d e f g Currie, Philip J.; Carpenter, Kenneth (2000). "A new specimen of Acrocanthosaurus atokensis (Theropoda, Dinosauria) from the Lower Cretaceous Antlers Formation (Lower Cretaceous, Aptian) of Oklahoma, USA". Geodiversitas. 22 (2): 207–246. Archived from the original on November 14, 2007.
- ^ a b D'Emic, Michael D.; Melstrom, Keegan M.; Eddy, Drew R. (2012). "Paleobiology and geographic range of the large-bodied Cretaceous theropod dinosaur Acrocanthosaurus atokensis". Palaeogeography, Palaeoclimatology, Palaeoecology. 333–334: 13–23. Bibcode:2012PPP...333...13D. doi:10.1016/j.palaeo.2012.03.003.
- ^ Ratkevich, Ronald P. (1997). "Dinosaur remains of southern Arizona". In Wolberg, Donald L.; Stump, Edward; Rosenberg, Gary (eds.). Dinofest International: Proceedings of a Symposium Held at Arizona State University. Philadelphia: Academy of Natural Sciences. ISBN 978-0-935868-94-4.
- ^ Ratkevich, Ronald P. (1998). "New Cretaceous brachiosaurid dinosaur, Sonorasaurus thompsoni gen. et sp. nov., from Arizona". Journal of the Arizona-Nevada Academy of Science. 31 (1): 71–82.
- ^ Lipka, Thomas R. (1998). "The affinities of the enigmatic theropods of the Arundel Clay facies (Aptian), Potomac Formation, Atlantic Coastal Plain of Maryland". In Lucas, Spencer G.; Kirkland, James I.; Estep, J.W. (eds.). Lower and Middle Cretaceous Terrestrial Ecosystems. New Mexico Museum of Natural History and Science Bulletin 14. pp. 229–234.
- ^ Harris, Jerald D. (1998). "Large, Early Cretaceous theropods in North America". In Lucas, Spencer G.; Kirkland, James I.; Estep, J.W. (eds.). Lower and Middle Cretaceous Terrestrial Ecosystems. New Mexico Museum of Natural History and Science Bulletin 14. pp. 225–228.
- ^ a b Paul, Gregory S. (2016). The Princeton Field Guide to Dinosaurs. Princeton University Press. p. 102. ISBN 978-1-78684-190-2. OCLC 985402380.
- ^ Canale, J.I.; Apesteguía, S.; Gallina, P.A.; Mitchell, J.; Smith, N.D.; Cullen, T.M.; Shinya, A.; Haluza, A.; Gianechini, F.A.; Makovicky, P.J. (July 7, 2022). "New giant carnivorous dinosaur reveals convergent evolutionary trends in theropod arm reduction". Current Biology. 32 (14): 3195–3202.e5. doi:10.1016/j.cub.2022.05.057. PMID 35803271.
- ^ Snively, E.; O’Brien, H.; Henderson, D.M.; Mallison, H.; Surring, L.A.; Burns, M.E.; Holtz, T.R.; Jr, Russell, A.P.; Witmer, L.M.; Currie, P.J.; Hartman, S.A.; Cotton, J.R. (2019). "Lower rotational inertia and larger leg muscles indicate more rapid turns in tyrannosaurids than in other large theropods". PeerJ. 7: e6432. doi:10.7717/peerj.6432. PMC 6387760. PMID 30809441.
- ^ Campione, Nicolás E.; Evans, David C. (2020). "The accuracy and precision of body mass estimation in non-avian dinosaurs". Biological Reviews. 95 (6): 1759–1797. doi:10.1111/brv.12638. ISSN 1469-185X. PMID 32869488. S2CID 221404013.
- ^ a b c d e f Holtz, Thomas R.; Molnar, Ralph E.; Currie, Philip J. (2004). "Basal Tetanurae". In Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (eds.). The Dinosauria (Second ed.). Berkeley: University of California Press. pp. 71–110. ISBN 978-0-520-24209-8.
- ^ Molnar, Ralph E.; Kurzanov, Sergei M.; Dong Zhiming (1990). "Carnosauria". In Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (eds.). The Dinosauria (First ed.). Berkeley: University of California Press. pp. 169–209. ISBN 978-0-520-06727-1.
- ^ a b Naish, Darren; Hutt, Stephen; Martill, David M. (2001). "Saurischian Dinosaurs 2: Theropods". Dinosaurs of the Isle of Wight. London: The Palaeontological Association. pp. 242–309. ISBN 978-0-901702-72-2.
- ^ Brusatte, Stephen L.; Benson, Roger B. J.; Chure, Daniel J.; Xu, Xing; Sullivan, Corwin; Hone, David W. E. (2009). "The first definitive carcharodontosaurid (Dinosauria: Theropoda) from Asia and the delayed ascent of tyrannosaurids" (PDF). Naturwissenschaften. 96 (9): 1051–8. Bibcode:2009NW.....96.1051B. doi:10.1007/s00114-009-0565-2. hdl:20.500.11820/33528c2e-0c9c-4160-8693-984f077ee5d0. PMID 19488730. S2CID 25532873.
- ^ a b Benson, Roger B. J.; Carrano, Matthew T.; Brusatte, Stephen L. (2009). "A new clade of archaic large-bodied predatory dinosaurs (Theropoda: Allosauroidea) that survived to the latest Mesozoic" (PDF). Naturwissenschaften. 97 (1): 71–8. Bibcode:2010NW.....97...71B. doi:10.1007/s00114-009-0614-x. PMID 19826771. S2CID 22646156.
- ^ Romer, Alfred S. (1956). Osteology of the Reptiles. Chicago: University of Chicago Press. p. 772pp. ISBN 978-0-89464-985-1.
- ^ Walker, Alick D. (1964). "Triassic reptiles from the Elgin area: Ornithosuchus and the origin of carnosaurs". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 248 (744): 53–134. Bibcode:1964RSPTB.248...53W. doi:10.1098/rstb.1964.0009.
- ^ Romer, Alfred S. (1966). Vertebrate Paleontology (Third ed.). Chicago: University of Chicago Press. p. 468pp. ISBN 978-0-7167-1822-2.
- ^ Carroll, Robert L. (1988). Vertebrate Paleontology and Evolution. W.H. Freeman and Company. ISBN 978-0-7167-1822-2.
- ^ Lambert, David; Diagram Group (1983). "Spinosaurids". A Field Guide to Dinosaurs. New York: Avon Books. pp. 84–85. ISBN 978-0-380-83519-5.
- ^ Norman, David B. (1985). "Carnosaurs". The Illustrated Encyclopedia of Dinosaurs: An Original and Compelling Insight into Life in the Dinosaur Kingdom. New York: Crescent Books. pp. 62–67. ISBN 978-0-517-46890-6.
- ^ a b Glut, Donald F. (1982). The New Dinosaur Dictionary. Secaucus, NJ: Citadel Press. pp. 39, 48. ISBN 978-0-8065-0782-8.
- ^ Paul, Gregory S. (1988). "Genus Acrocanthosaurus". Predatory Dinosaurs of the World. New York: Simon & Schuster. pp. 314–315. ISBN 978-0-671-61946-6.
- ^ Olshevsky, George (1991). A Revision of the Parainfraclass Archosauria Cope, 1869, Excluding the Advanced Crocodylia. San Diego: Publications Requiring Research. p. 196pp.
- ^ Holtz, Thomas R. (September 1994). "The phylogenetic position of the Tyrannosauridae: implications for theropod systematics". Journal of Paleontology. 68 (5): 1100–1117. Bibcode:1994JPal...68.1100H. doi:10.1017/S0022336000026706. ISSN 0022-3360. S2CID 129684676.
- ^ Suarez, Celina A.; Frederickson, Joseph; Cifelli, Richard L.; Pittman, Jeffrey G.; Nydam, Randall L.; Hunt-Foster, ReBecca K.; Morgan, Kirsty (October 21, 2021). "A new vertebrate fauna from the Lower Cretaceous Holly Creek Formation of the Trinity Group, southwest Arkansas, USA". PeerJ. 9: e12242. doi:10.7717/peerj.12242. ISSN 2167-8359. PMC 8542373. PMID 34721970. S2CID 239497288.
- ^ Eddy, Drew R.; Clarke, Julia A. (2011). Farke, Andrew (ed.). "New Information on the Cranial Anatomy of Acrocanthosaurus atokensis and Its Implications for the Phylogeny of Allosauroidea (Dinosauria: Theropoda)". PLoS ONE. 6 (3): e17932. Bibcode:2011PLoSO...617932E. doi:10.1371/journal.pone.0017932. PMC 3061882. PMID 21445312.
- ^ Novas, Fernando E. (2013). "Evolution of the carnivorous dinosaurs during the Cretaceous: The evidence from Patagonia". Cretaceous Research. 45: 174–215. Bibcode:2013CrRes..45..174N. doi:10.1016/j.cretres.2013.04.001.
- ^ D'Emic, Michael; Melstrom, Keegan; Eddy, Drew (May 15, 2012). "Paleobiology and geographic range of the large-bodied Cretaceous theropod dinosaur Acrocanthosaurus atokensis". Palaeogeography, Palaeoclimatology, Palaeoecology. 333–334: 13–23. Bibcode:2012PPP...333...13D. doi:10.1016/j.palaeo.2012.03.003.
- ^ Sakamoto, Manabu (July 12, 2022). "Estimating bite force in extinct dinosaurs using phylogenetically predicted physiological cross-sectional areas of jaw adductor muscles". PeerJ. 10: e13731. doi:10.7717/peerj.13731. ISSN 2167-8359. PMC 9285543. PMID 35846881.
- ^ a b c d e Senter, Phil; Robins, James H. (2005). "Range of motion in the forelimb of the theropod dinosaur Acrocanthosaurus atokensis, and implications for predatory behaviour". Journal of Zoology. 266 (3): 307–318. doi:10.1017/S0952836905006989.
- ^ a b Franzosa, Jonathan; Rowe, Timothy. (2005). "Cranial endocast of the Cretaceous theropod dinosaur Acrocanthosaurus atokensis". Journal of Vertebrate Paleontology. 25 (4): 859–864. doi:10.1671/0272-4634(2005)025[0859:CEOTCT]2.0.CO;2. S2CID 10173542.
- ^ "Texas drought uncovers dinosaur footprints from 113 million years ago".
- ^ a b Bird, Roland T. (1941). "A dinosaur walks into the museum". Natural History. 43: 254–261.
- ^ Rogers, Jack V. (2002). "Theropod dinosaur trackways in the Lower Cretaceous (Albian) Glen Rose Formation, Kinney County, Texas". Texas Journal of Science. 54 (2): 133–142.
- ^ Hawthorne, J. Michael; Bonem, Rena M.; Farlow, James O.; Jones, James O. (2002). "Ichnology, stratigraphy and paleoenvironment of the Boerne Lake Spillway dinosaur tracksite, south-central Texas". Texas Journal of Science. 54 (4): 309–324.
- ^ Langston, Wann (1974). "Non-mammalian Comanchean tetrapods". Geoscience and Man. 3: 77–102.
- ^ a b Farlow, James O. (2001). "Acrocanthosaurus and the maker of Comanchean large-theropod footprints". In Tanke, Darren; Carpenter, Ken (eds.). Mesozoic Vertebrate Life. Bloomington: Indiana University Press. pp. 408–427. ISBN 978-0-253-33907-2.
- ^ a b Lockley, Martin G. (1991). Tracking Dinosaurs: A New Look at an Ancient World. Cambridge: Cambridge University Press. p. 252pp. ISBN 978-0-521-39463-5.
- ^ Thomas, David A.; Farlow, James O. (1997). "Tracking a dinosaur attack". Scientific American. 266 (6): 48–53. Bibcode:1997SciAm.277f..74T. doi:10.1038/scientificamerican1297-74.
- ^ Molnar, R.E., 2001, 절지동물 고병리학: 문헌 조사:In: 중생대 척추동물 생명, Tanke, D. H. and Carpenter, K. 편집, 인디애나 대학 출판부, p. 337-363.
- ^ Jacobs, Louis L.; Winkler, Dale A.; Murry, Patrick A. (1991). "On the age and correlation of Trinity mammals, Early Cretaceous of Texas, USA". Newsletters on Stratigraphy. 24 (1–2): 35–43. doi:10.1127/nos/24/1991/35.
- ^ a b Brinkman, Daniel L.; Cifelli, Richard L.; Czaplewski, Nicholas J. (1998). "First occurrence of Deinonychus antirrhopus (Dinosauria: Theropoda) from the Antlers Formation (Lower Cretaceous: Aptian – Albian) of Oklahoma". Oklahoma Geological Survey Bulletin. 146: 1–27.
- ^ [온라인 published]
- ^ Wedel, Matthew J.; Cifelli, Richard L.; Kent Sanders, R. (2000). "Sauroposeidon proteles, a new sauropod from the Early Cretaceous of Oklahoma" (PDF). Journal of Vertebrate Paleontology. 20 (1): 109–114. doi:10.1671/0272-4634(2000)020[0109:SPANSF]2.0.CO;2. S2CID 55987496.
- ^ Winkler, Dale A.; Murry, Patrick A.; Jacobs, Louis L. (1997). "A new species of Tenontosaurus (Dinosauria: Ornithopoda) from the Early Cretaceous of Texas". Journal of Vertebrate Paleontology. 17 (2): 330–348. Bibcode:1997JVPal..17..330W. doi:10.1080/02724634.1997.10010978. Archived from the original on September 27, 2007.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||

.jpg)















