카르노 타우루스
Carnotaurus| 카르노 타우루스 시간 범위:백악기 후기 OSDCPTJKPg N↓ Ꞓ 71마 PreꞒ(마스트 리히 트세)[1]. | |
|---|---|
 | |
| Chlupáč 박물관 프라하에서 뼈대만 앙상하게 남은 출연진 기마다. | |
| 과학적 분류 | |
| 왕국: | 애니멀리아 |
| Phylum: | 코다타 |
| 계통 군: | 소공니아 |
| 계통 군: | 사우리스치아 |
| 계통 군: | 테로포다류 |
| 가족: | †Abelisauridae |
| 계통 군: | †Furileusauria |
| Tribe: | †Carnotaurini |
| 지누스: | †카르노 타우루스 보나파르트, 1985년 |
| 종: | †C. sastrei |
| 이항 이름 | |
| †카르노 타우루스 sastrei 보나파르트, 1985년 | |
theropod의 공룡은 남 아메리카의 길이였으며 백악기 후기에, 7169만년 전쯤까지 카르노 타우루스 /ˌkɑːrnoʊˈtɔːrəs/은 유인원 속이다.유일한 종은 카르노 타우루스 sastrei.단일 잘 보존된 유골이 알려진, 그것이 하나의 남반구에서best-understood 수각류 공룡의.해골은 1984년에 발견된 추부트 주 아르헨티나의 라 콜로니아 구성 변화는 바위에서 발견되었다.그 Abelisauridae, 큰 수각류 공룡의 곤드와나 대륙의 남쪽 대륙의 늦은 백악기 동안 큰predatorial 틈새를 점거하고 그룹의 카르노 타우루스 파생된 회원이다.그 Abelisauridae 이내에, 이 속은 종종 Brachyrostra,short-snouted 형태 남미로 제한된 clade의 회원국으로 간주된다.
카르노 타우루스는 가볍게 지어진 두 포식자인,(관리 긴톤, 4,900 짧은톤)최소한 800만톤의 무게 7.5~8m(24.6에 26.2ft)을 측정하고 있습니다.한 theropod로서, 카르노 타우루스 고도로 독특한 전문화된 있었다.그것은 두꺼운 뿔 위에, 모든 다른 육식 공룡들은에서 형상을 뻔하고 아주 깊은 두개골은 근육 목에 앉아 있었다.카르노 타우루스 더 작은, 남아 있는 말로는, 길고 날씬한 뒷다리에 의해 특징 지어졌습니다.해골은 광범위한 피부 인상에, 작은,non-overlapping 비늘 직경이 대략 5mm의 모자이크를 보여 주는 보존 되어 있다.모자이크는 동물의 옆구리에 줄지어 있는 커다란 돌기 때문에 방해를 받았으며, 깃털의 흔적은 찾아볼 수 없다.
독특한 뿔과 근육질의 목은 서로 싸우는 데 사용되었을지도 모른다.별개의 연구에 따르면, 경쟁하는 개인은 빠른 머리를 때리거나, 두개골의 윗부분을 천천히 밀거나, 뿔을 충격 흡수제로 사용하여 서로 정면으로 부딪힘으로써 서로 싸웠을 수 있다.카르노타우루스의 먹이 습관은 여전히 불분명하다. 어떤 연구는 이 동물이 용각류와 같은 매우 큰 먹이를 사냥할 수 있었다고 제안하는 반면, 다른 연구들은 이 동물이 주로 상대적으로 작은 동물들을 먹이로 삼았다는 것을 발견했다.그것의 뇌 공동은 청각과 시력이 덜 발달한 반면, 예민한 후각을 암시한다.카노타우루스(Carnotaurus)는 아마도 달리기에 잘 적응했고 아마도 가장 빠른 큰 테로포드 중 하나였을 것이다.
디스커버리
유일한 골격(홀로타입 MACN-CH 894)은 1984년 아르헨티나의 고생물학자 호세 보나파르트가 이끄는 탐험대에 의해 발굴되었다.[A]이 탐험은 또한 독특한 가시가 있는 용각류 아마르가사우루스도 되찾았다.[3]1976년 시작돼 내셔널지오그래픽협회가 후원한 '남미의 쥬라기·백악기 육상 척추동물' 프로젝트 내 여덟 번째 원정이었다.[3][B]골격은 잘 보존되어 있고 (아직도 서로 연결되어 있으며) 꼬리 뒷부분 3분의 2만, 아랫부분의 상당 부분, 뒷발은 풍화작용에 의해 파괴된다.[C][5]이 유골은 의 융합 [6]봉합술에서 알 수 있듯이 성인 개인의 것이었다.목뼈가 몸통 위로 등을 구부린 채 전형적인 사망 자세를 보이는 등 오른쪽에 누운 채 발견됐다.[7]특이하게도 광범위한 피부인상으로 보존되어 있다.[D]이러한 인상의 의의를 고려하여 원래의 발굴현장을 재조사하기 위한 2차 탐사에 착수하여 몇 개의 추가 피부 패치를 회복하게 되었다.[7]두개골은 화석화 과정에서 변형되었는데, 왼쪽의 코뼈는 오른쪽을 기준으로 앞으로 이동했고, 코뼈는 위로 밀려 올라갔으며, 뒤로 밀린 뼈는 위쪽으로 이동했다.변형은 위턱의 위쪽으로의 곡률을 과장하기도 했다.[E]콧구멍은 두개골의 뒷부분보다 변형에 의해 더 강하게 영향을 받았으며, 아마도 후자의 더 높은 강성 때문일 것이다.위나 아래를 보면 위턱이 아래턱보다 U자형이 적어 겉보기에는 불일치가 있었다.이 불일치는 측면으로부터 작용하는 변형으로 인해 위턱에는 영향을 주었지만 아래턱에는 영향을 주지 못하는데, 후자 내 관절의 유연성이 더 크기 때문일 수 있다.[1]

이 유골은 아르헨티나 추부트 주 텔센부의 바자다 모레노 인근 농장에 모아졌다.[5]아주 단단한 종류의 암석인 큰 적혈구 결석 속에 박혀 있었기 때문에 준비는 복잡하고 천천히 진행되었다.[9][5]1985년 보나파르트는 카르노타우루스 사스트레이를 새로운 속과 종으로 제시하고 두개골과 아래턱을 간략하게 기술한 노트를 발표했다.[5]총칭 카노타우루스(Carnotaurus)는 라틴 카르노[carnis]("flesh")와 타우러스("bull")에서 유래했으며, 동물의 황소 같은 뿔을 암시하는 "고기 먹는 황소"로 번역할 수 있다.[10]구체적인 이름인 사스트레이는 유골이 발견된 목장의 주인인 엔젤 사스트레를 기린다.[11]1990년에 이어 전체 골격에 대한 포괄적인 설명이 이어졌다.[4]아벨리사우루스 다음으로 카르노타우루스는 발견된 아벨리사우루스과의 두 번째 일원이었다.[12]몇 년 동안, 그것은 그 가족 중에서 단연코 가장 잘 이해되는 구성원이었고, 또한 남반구에서 가장 잘 이해되는 종족이었다.[13][14]오카사우루스, 마장가사우루스, 스코르피오베네이터 등 유사하게 잘 보존된 아벨리사우루스(Aucasaurus), 아벨리사우루스(Majungasaurus), 스콜피오베네이터(Skorpiovenator)[F] 등이 묘사되어 과학자들이 카르노타우루스 해부학의 특정 측면을 재평가할 수 있게 되었다.이 홀로타입의 뼈대는 아르헨티나 자연과학 박물관의 베르나르디노 리바다비아에 전시되어 있다.[G] 복제품은 이 박물관과 전 세계의 다른 박물관들에서 볼 수 있다.[15]조각가 스테판과 실비아 체르카스는 이전에 로스앤젤레스 카운티의 자연사 박물관에 전시되었던 카르노타우루스 실물 크기의 조각품을 제작했다.1980년대 중반 박물관에 의해 주문된 이 조각품은 아마도 정확한 피부를 보여주는 최초의 생명체 복원품일 것이다.[7][16]
설명

카르노타우루스(Carnotaurus)[17]는 몸집이 크지만 가벼운 체격의유일하게 알려진 개체는 길이가 약 7.5–8m(24.6–26.2ft)[H][I][19]로 카노타우루스가 가장 큰 아벨리사우루스 중 하나가 되었다.[J][K][19]매우 불완전한 에크릭시나토사우루스와 아마도 아벨리사우루스는 크기가 비슷하거나 더 컸을 것이다.[L][M][N]2016년 한 연구에서는 8.9m(29.2ft)의 피크노네모사우루스만이 카르노타우루스보다 더 길다는 것을 발견했다; 그것은 7.8m(25.6ft)로 추정되었다.[21]Its mass is estimated to have been 1,350 kg (1.33 long tons; 1.49 short tons),[O] 1,500 kg (1.5 long tons; 1.7 short tons),[P] 2,000 kg (2.0 long tons; 2.2 short tons),[19] 2,100 kg (2.1 long tons; 2.3 short tons),[Q] and 1,306–1,743 kg (1.285–1.715 long tons; 1.440–1.921 short tons)[24] in separate studies that used different estimation methods.카르노타우루스(Carnotaurus)는 특히 두개골, 척추골, 전림부의 특징에서 볼 수 있듯이 고도의 전문화된 테로포드였다.[R]반면 골반과 뒷다리는 비교적 보수적인 상태를 유지하여 보다 기초적인 세라토사우루스의 그것과 닮았다.골반과 뒷다리 모두 길고 가늘었다.개개의 왼쪽(근골)은 길이가 103cm이지만 평균 지름은 11cm에 불과하다.[S]
스컬

길이 59.6cm(23.5인치)의 이 두개골은 다른 대형 육식공룡보다 비례적으로 짧고 깊었다.[T][U]주둥이는 적당히 넓어서, 세라토사우루스처럼 더 많은 기초적인 테로포드에서 볼 수 있는 것처럼 테이퍼링하지 않았고, 턱은 위로 구부러져 있었다.[25]눈에 띄는 뿔이 눈 위로 비스듬히 튀어나왔다.이 뿔들은 전두골에 의해 형성된 두껍고 원추형이며, 내부는 단단하며, 단면으로 다소 수직으로 평평하게 되어 있으며, 길이가 15cm(5.9인치)로 측정되었다.[V][6][1]1990년 보나파르트는 이 뿔들이 아마도 훨씬 더 긴 각질 피복의 뼈 중심부를 형성했을 것이라고 제안했다.[W]마우리시오 세로니와 동료들은 2020년 뿔이 각질 피복을 지지한다는 데는 동의했지만, 이러한 피복이 뼈 중심부보다 그리 길지는 않았을 것이라고 주장했다.[1]
다른 공룡들과 마찬가지로, 두개골은 양쪽에 6개의 주요 두개골 개구부에 의해 구멍이 뚫렸다.이 개구부의 가장 앞부분인 (뼈가 굵은 콧구멍)은 직사각형 이하였고 앞뒤로 방향을 잡았지만, 세라토사우루스와 같은 다른 세라토사우루스처럼 측면으로 경사진 것은 아니었다.이 개구부는 비강과 프리맥실라에 의해서만 형성되었고, 일부 관련 세라토룡에서는 맥실라 또한 이 개구부에 기여했다.뼈로 된 콧구멍과 궤도(눈구멍) 사이에는 항고선 페네스트라가 있었다.카르노타우루스에서는 이 오프닝이 긴 것보다 높은 반면 스코르피오베네이터, 마장가사우루스 등 관련 형식에서는 높은 편이었다.항우울증은 더 큰 우울증으로 경계되었는데 , 는 앞과 뒤에 있는 맥실라의 움푹 들어간 부분들에 의해 형성되었다.모든 아벨리사우루스처럼 이 우울증은 카르노타우루스에서는 작았다.항균포사 앞쪽 하단 모서리에 작은 개구부가 들어 있었는데, 이 구멍은 맥실라 내 공기로 채워진 공동으로 이어졌다.[1]눈은 열쇠 구멍 모양의 궤도의 상부에 위치했다.[X]이 윗부분은 비례적으로 작고 아원형이었으며, 전진 투영에 의해 궤도의 아랫부분에서 분리되었다.[1]그것은 약간 앞으로 회전했고, 아마도 어느 정도의 쌍안경도 허용했을 것이다.[Y]궤도의 열쇠구멍과 같은 모양은 표시된 두개골의 단축과 관련이 있을 가능성이 있으며, 관련된 짧은 코로 만들어진 아벨리사우루스에서도 발견된다.[1]모든 아벨리사우루스(Abelisaurids)에서와 마찬가지로 (눈 사이의 두개골 지붕에 있는)는 궤도에서 제외되었다.궤도 뒤로는 옆쪽과 두개골 위쪽에 있는 두 개의 개구부가 있었다.infratmporal fenestra는 키가 크고, 키가 작고, 신장 모양인 반면, supratemporal fenestra는 짧고 네모꼴이었다.또 다른 개구부인 , 는 아래쪽 턱에 위치해 있었다 – Carnotaurus에서, 이 개구부는 비교적 컸다.[1]

위턱의 양옆에는 전축 4개와 맥시니 12개가 달려 있었고,[Z] 아래턱에는 각 면당 15개의 의치니가 달려 있었다.[AA][1]치아는 다른 아벨리사우루스류에서 볼 수 있는 아주 짧은 치아와 달리 [9]길고 가느다란 것으로 묘사되어 있었다.[25]그러나 세로니와 동료들은 2020년 두개골에 대한 설명에서 분출된 모든 치아가 굴착 과정에서 심하게 손상되어 나중에 석고로 재건되었다고 진술했다(Bonaparte, 1990년, 일부 아래턱 치아가 조각난 것에만 주목했다).[1][AB]따라서 치아의 형태에 대한 신뢰할 수 있는 정보는 턱으로 여전히 감싸고 있는 치아와 치아뿌리 교체에 한정되며, CT 영상을 이용하여 연구할 수 있다.[1]교체된 치아는 낮고 납작한 크라운을 가지고 있었으며, 간격을 촘촘히 하고, 약 45°[1]의 앞쪽으로 기울어져 있었다.보나파르트는 1990년 묘사에서 아래턱이 얕고 약하게 만들어져 있으며 (앞턱뼈는) 맨 뒷턱 뼈와 단 두 개의 접촉점만으로 연결되어 있다고 언급했다. 이는 튼튼해 보이는 두개골과는 대조적이다.[9][AC]대신 세로니와 동료들은 틀니뼈와 맨 뒷턱뼈 사이에 여러 개 있지만 느슨한 연결부를 발견했다.그러므로 이 발음은 매우 유연했지만 반드시 약하지는 않았다.[1]의치의 아래쪽 여백은 볼록했고, 반면에 마장가사우루스에서는 곧았다.[1]

아래턱은 오십자로 된 효모이드 뼈로, 동물이 살아있다면 있을 위치에서 발견되었다.혀의 근골과 다른 여러 근육을 지탱하는 이 가느다란 뼈는 공룡에서는 거의 찾아볼 수 없다. 왜냐하면 그들은 종종 장골과 연결되어 있지 않기 때문이다. 그래서 쉽게 길을 잃기 때문이다.[AD][26][1]카르노타우루스에서는 세 개의 효이형 뼈가 보존된다: 한 쌍의 곡선이 있고 막대 모양의 세라토브란치알은 한 쌍의 사다리꼴 원소인 바시알로 표현된다.카르노타우루스(Carnotaurus)는 유일하게 기초가 알려진 비(非)아비안 세로포드(non-avian)이다.[1]두개골의 뒷면에는 다른 아벨리사우루스처럼 뇌케이스를 둘러싼 잘 발달된 공기로 가득 찬 방이 있었다.두 개의 분리된 챔버 시스템, 즉 중이강과 연결된 파라티움파네틱 시스템과 목의 공기 주머니가 밖으로 자라나면서 생기는 챔버가 존재했다.[24]
두개골에는 뿔과 매우 짧고 깊은 두개골을 포함한 많은 자동형상(distogning features)이 있다.맥실라는 대뇌반동맥동(주둥이의 공기 통로)에 의해 발굴되었을 대뇌정맥동 위로 굴착이 있었다.눈의 액체를 운반하는 내측관(내측)은 기능이 불확실한 운하를 통해 외측(내측) 표면으로 빠져나갔다.다른 제안된 자동 형태로는 에서 깊고 긴 공기로 가득 찬 굴착물 및 의 의 장기화된 우울증이 있다.[1]
척추뼈

척추 기둥은 10개의 경추(목), 12개의 등골, 6개의 융접된 천골[AE], 그리고 알 수 없는 수의 카우달(꼬리) 척추뼈로 구성되어 있었다.[4]목은 다른 테로포드에서 S 커브를 보기는커녕 거의 일직선이었고, 특히 밑부분을 향해 유난히 넓었다.[27]목의 척추 윗부분에는 에피포피시스라고 불리는 두 줄로 늘어뜨리고 위쪽으로 향하는 뼈의 과정이 있어 목 척추 윗부분에 매끄러운 수조를 만들었다.이 과정들은 척추의 가장 높은 지점으로, 비정상적으로 낮은 가느다란 과정 위에 우뚝 솟아 있었다.[4][26]에피포피스는 아마도 두드러지게 강한 목 근육질의 부착 부위를 제공했을 것이다.[AF]매우 변형된 카우달 늑골에 의해 형성된 꼬리에 V자 모양으로 위쪽으로 돌출된 앞면에는 앞 꼬리 척추뼈의 매끄럽고 평평한 상단 표면을 형성하는 유사한 이중 행이 있었다.각 꼬막 늑골의 끝에는 앞 척추뼈의 꼬막 늑골과 연결된 전방 돌출 고리 모양의 확장이 제공되었다.[26][28]
포림프스


포림들은 티라노사우루스를 포함한 다른 어떤 거대한 육식 공룡들보다 비례적으로 짧았다.[AG]팔뚝은 상팔의 4분의 1밖에 되지 않았다.손에는 카팔리아가 없어서 메타카르팔들은 팔뚝과 직접 관절을 이루었다.[29]손은 네 개의 기본 숫자를 보여주었는데,[4] 분명히 이것들 중 두 개만이 손가락 뼈로 끝났을 뿐이고, 네 번째 손가락은 외부 '스푸르'를 나타낼 수 있는 하나의 부목 모양의 메타카르팔로 구성되어 있었다.손가락 자체가 융합되어 움직이지 않았고, 발톱이 부족했을지도 모른다.[30]카르노타우루스(Carnotaurus)는 다른 모든 아벨리사우루스(Abelisaurids)와는 비례적으로 짧고 튼튼한 앞다리를 가지고 있으며, 손의 가장 긴 뼈로 네 번째 부목 같은 메타카르팔(Metacarpal)을 가지고 있다는 점에서 차이가 있었다.[29]2009년의 한 연구는 자극 전달을 담당하는 신경 섬유가 오늘날의 에무스와 키위에서도 볼 수 있는 정도로 감소했기 때문에 아벨리사우루스에서는 팔이 잔재였다고 시사했다.[31]
스킨

카르노타우루스(Carnotaurus)는 상당수의 화석 피부 인상과 함께 발견된 최초의 공룡이다.[7]골격의 오른쪽 아래에서 발견되는 이러한 인상은 아래턱,[7] 목 앞쪽, 어깨 거들, 늑골 우리를 포함한 다른 신체 부위에서 온다.[AH]가장 큰 피부 조각은 꼬리의 앞부분과 일치한다.[AI]원래, 두개골의 오른쪽도 커다란 피부 조각으로 덮여 있었다. 이것은 두개골이 준비되었을 때 인식되지 않았고, 이 조각들은 우연히 파괴되었다.[7]그러나 두개골의 표면 질감은 두개골의 덮개에 대한 추론을 가능하게 한다.홈, 구덩이, 작은 개구부가 있는 험모크 표면은 주둥이의 옆면과 앞쪽에서 발견되며 비늘 모양의 덮개를 나타내며, 아마도 오늘날의 악어들처럼 평평한 비늘로 덮었을 것이다.주둥이의 윗부분은 수많은 작은 구멍과 뾰족탑으로 조각되었다. – 이 질감은 아마도 옥수수 덮개(혼합 덮개)와 상관관계가 있을 것이다.그런 패드는 마장가사우루스에서도 발생했지만 아벨리사우르스와 루고프에서는 없었다.아마도 큰 비늘이 눈을 둘러싸고 있었을 것이다. 그것은 눈물샘과 후두골에 세로 홈이 있는 험모크 표면에서 알 수 있다.[1][AJ]
피부는 직경 약 5-12 mm(0.20–0.47 in)의 겹치지 않는 다각형 비늘의 모자이크로 만들어졌다.이 모자이크는 얇고 평행한 홈으로 나뉘었다.[AK]메스는 머리를 제외한 다른 신체 부위에 걸쳐 유사했는데, 이것은 분명히 다른, 불규칙한 비늘 패턴을 보였다.[AL][15]깃털의 흔적이 없다.[7]목과 등, 꼬리 부분에 불규칙한 줄무늬로 더 큰 혹 모양의 구조물이 분포되어 있었다.이 돌기들은 지름 4~5cm(1.6~2.0인치)에 높이 5cm(2.0인치)로 중간선 능선이 낮은 경우가 많았다.그것들은 서로 8~10cm(3.1~3.9인치) 간격으로 세워졌고 동물의 윗부분을 향해 커졌다.이 돌기는 해드로사우리드("덕부리") 공룡의 몸체 중간선을 따라 흐르는 부드러운 프릴에서 볼 수 있는 것과 유사한 응축된 스쿠트 군집인 특징 척도를 나타낸다.이 구조물들은 뼈를 포함하고 있지 않았다.[AM][7][32]스테판 체르카스(1997)는 이 구조물들이 전투에서 제한된 보호를 제공하는 현대 이구아나의 목에서 유사한 구조물들을 발견할 수 있다고 주장하면서, 같은 종의 구성원들과 싸우면서 동물의 옆구리를 보호했을 수도 있다고 제안했다.[7]
2021년에 발표된 카르노타우루스의 피부에 대한 보다 최근의 연구는 신체의 비늘에 대한 이전의 묘사가 부정확하며, 더 큰 형상 비늘은 오래된 예술적 묘사와 삽화처럼 분리된 줄에 분포하지 않고 신체를 따라 무작위로 분포하였음을 시사한다.또한 신체를 따라 다른 영역을 따라 형상 눈금이 점진적으로 변화한 흔적은 없다.카르노타우루스의 지하 비늘은 크기가 작거나 길쭉한 것에서부터 큰 것, 다각형까지, 흉부, 척추, 꼬리 부분의 원형부터 발톱까지 비교적 가변적이었다.이 척도 분화는 몸집이 크고 활동적인 생활방식으로 인해 체온 조절을 통해 체열을 조절하고 과도한 열을 방출하는 것과 관련이 있을 수 있다.[33]
분류


카르노타우루스(Carnotaurus)는 아벨리사우루스과(Abelisaurae)의 가장 잘 이해되는 종 중 하나로, 고대 남부 초대륙인 곤드와나(Gondwana)에 국한된 큰 에트로포드과의 종이다.아벨리사우루스(Abelisaurids)는 곤드와나 후기 백악기의 지배적인 포식자로, 카카로돈토사우루스(carcharodontosaurids)를 대체하고 북부 대륙의 티라노사우루스(tyranhnosaurids)가 채운 생태학적 틈새를 점령했다.[17]두개골과 팔의 단축은 물론 경추와 카우달 척추뼈의 특이성 등 이 가족 내에서 진화한 몇 가지 주목할 만한 특징들이 카르노타우루스에서는 다른 아벨리사우루스보다 더 두드러졌다.[AN][AO][28]
아벨리사우루스과 내의 관계는 논의되고 있지만, 카르노타우루스는 피복재학적 분석에 의해 가족에서 가장 파생된 구성원 중 하나라는 것을 일관되게 보여주고 있다.[AP]가장 가까운 친척은 오카사우루스나[34][35][36][37] 마장가사우루스였을 것이다.[38][39][40]이와는 대조적으로 2008년 검토에서는 카르노타우루스가 어느 한 속과 밀접한 관계가 없다는 것을 시사했고, 대신 일로켈레시아를 자매 택시로 제안했다.[AQ]후안 카날과 동료들은 2009년에 카르노타우르스를 포함하되 마장가사우르스를 포함하지 않기 위해 새로운 쇄골 브라키로스트라를 설립했다. 이 분류는 그 이후로 많은 연구가 이어졌다.[34][37][41]
카르노타우루스(Carnotaurus)는 아벨리사우루스과(Carnotaurinae)와 카르노타우리니(Carnotaurini)의 두 하위집단에 해당된다.고생물학자들은 이러한 집단을 보편적으로 수용하지 않는다.카르노타우리나에(Carnotaurinae)는 대부분의 연구에서 기초 구성원으로 간주되는 아벨리사우르스를 제외한 모든 파생 아벨리사우르스를 포함하도록 정의되었다.[42]그러나 2008년 한 리뷰는 아벨리사우르스가 그 대신 파생된 아벨리사우루스라는 것을 암시했다.[AR]카르노타우리니는 카르노타우루스와 아우카사우루스가 형성한 쇄골의 이름을 붙이자고 제안되었다.[35] 오직 카노타우루스의 가장 가까운 친척으로 오카사우루스를 생각하는 고생물학자들만이 이 집단을 사용한다.[43]
아래는 카날레와 동료들이 2009년에 발표한 클래도그램이다.[34]
| 까르노타우리나과 |
| ||||||||||||||||||||||||||||||
고생물학
뿔의 기능

카르노타우루스(Carnotaurus)는 전두골에 뿔이 달린 유일한 육식성 두발 동물이다.[44]이 뿔들의 사용이 완전히 명확한 것은 아니다.비록 같은 종의 구성원에 대한 구애나 인정을 위한 전시의 사용도 가능하지만, 몇몇 해석은 상호간의 싸움이나 사냥감을 죽이는 데 사용하는 것에 중점을 두고 있다.[1]
그레그 폴(1988)은 뿔이 무기들을 버팅하고 있으며 작은 궤도가 전투 중에 눈을 다칠 가능성을 최소화했을 것이라고 제안했다.[9]제라르도 마제타와 동료(1998)는 카노타우루스가 숫양과 비슷한 방식으로 뿔을 사용했다고 제안했다.이들은 목 근육계가 각각 5.7m/s의 속도로 정면 충돌하는 두 사람의 힘을 흡수할 정도로 강하다고 계산했다.[22]페르난도 노바스(2009)는 몇 가지 골격의 특징을 머리로 타격하기 위한 적응으로 해석했다.[AS]그는 두개골이 짧으면 관성의 순간을 줄여 머리가 더 빨리 움직였을 것이고, 반면 근육질의 목은 머리를 강하게 때릴 수 있었을 것이라고 제안했다.그는 또한 머리와 목에 의한 충격에 견딜 수 있도록 진화했을 수 있는 척추의 강화된 강성과 강도에 주목했다.[AT]
다른 연구들은 경쟁 관계에 있는 카르노타우루스가 빠른 머리를 날리지 않고 두개골의 윗부분을 가지고 서로 천천히 밀었다는 것을 보여준다.[44][45]마제타와 동료들은 2009년 이 뿔이 뇌에 손상되지 않은 압축력의 분배를 위한 장치였을 수도 있다고 주장했다.이것은 뿔의 평평한 윗면, 두개골 윗부분의 강하게 융합된 뼈, 그리고 두개골의 빠른 머리 타격에서 살아남을 수 없는 능력에 의해 지탱된다.[44]2018년 라파엘 델코트는 이 뿔이 현대의 해양 이구아나에서 볼 수 있듯이 느린 박치기나 밀치기, 또는 현대의 기린에서 볼 수 있는 것처럼 상대의 목과 옆구리를 강타할 때 사용되었을 수 있다고 제안했다.[37]후자의 가능성은 2011년 회의 논문에서 관련 마장가사우루스에게 제안된 바 있다.[46]
제라르도 마제타와 동료(1998)는 뿔이 작은 먹이를 다치게 하거나 죽이기 위해 사용되었을 수도 있다고 제안한다.뿔의 중심은 뭉툭하지만 각질 덮개가 있었다면 현대의 보비드 뿔과 비슷한 형태를 가졌을지도 모른다.하지만, 이것은 뿔이 동물에서 사냥 무기로 사용되는 유일한 예일 것이다.[22]
턱 기능 및 식이요법

마제타와 동료들이 카노타우루스의 턱 구조를 분석한 결과, 1998년, 2004년, 2009년 카노타우루스는 빠른 물기는 가능했지만 강한 물기는 불가능했던 것으로 나타났다.[22][23][44]현대의 악어에 대한 연구에서 알 수 있듯이, 작은 먹이를 잡을 때는 강한 물림보다 빠른 물림이 더 중요하다.[44]이 연구원들은 또한 두개골 내, 특히 아래턱의 융통성(키네시스)이 높다는 점에 주목했는데, 이는 현대의 뱀과 다소 유사하다.턱의 탄력성이라면 카노타우루스도 작은 먹잇감을 통째로 삼킬 수 있었을 것이다.또한 아래턱의 앞부분이 경첩되어 있어 위아래로 움직일 수 있었다.아래쪽으로 눌렀을 때 치아는 앞으로 돌출되어 카르노타우루스가 작은 먹잇감을 스파이크할 수 있었고, 치아가 위쪽으로 구부러졌을 때, 지금 거꾸로 돌출되어 있는 치아는 잡힌 먹잇감이 도망치는 것을 방해했을 것이다.[22]마제타와 동료들도 이 두개골이 커다란 먹잇감을 잡아당길 때 나타나는 힘을 견딜 수 있다는 것을 발견했다.[44]따라서 카르노타우루스는 상대적으로 작은 먹이를 주로 먹었을지 모르지만, 큰 공룡을 사냥할 수도 있었다.[44]2009년, Mazzetta와 동료들은 약 3,341 뉴턴의 물린력을 추정했다.[44]
이 해석은 프랑수아 테리엔과 동료들(2005)에 의해 질문되었는데, 그는 카르노타우루스의 무는 힘이 살아있는 어떤 사다리꼴보다도 가장 강한 미국 악어의 두 배라는 것을 발견했다.이 연구자들은 또한 현대의 코모도 왕 도마뱀과의 유사점에 주목했다: 아래턱의 굽힘 세기는 끝이 선형적으로 감소하는데, 이는 턱이 작은 먹잇감을 고도로 정밀하게 잡는 데 적합하지 않고 큰 먹이를 약화시키기 위해 날카로운 상처를 주는 데 적합했음을 보여준다.결과적으로, 이 연구에 따르면, 카노타우루스는 아마도 매복에 의해 주로 큰 동물들을 잡아먹었을 것이다.[47]세로니와 동료들은 2020년 융통성이 아래턱에만 국한됐다고 주장했지만, 두개골 지붕이 두꺼워지고 여러 개의 두개골 관절이 오식화되면서 두개골에는 키네시스가 없거나 조금밖에 없었다는 것을 알 수 있다.[1]
로버트 박커(1998)는 카노타우루스가 주로 매우 큰 먹이, 특히 용각류를 먹고 산다는 사실을 발견했다.그가 지적했듯이, 두개골의 몇 가지 변형, 즉 짧은 코, 비교적 작은 이빨, 그리고 두개골의 강한 뒷면(occiput)은 알로사우루스에서 독자적으로 진화했다.이러한 특징들은 위턱이 상처를 입히기 위해 톱니바퀴처럼 사용되었다는 것을 암시한다; 큰 용각류는 반복적인 공격으로 약해졌을 것이다.[48]
이동


마제타와 동료(1998년, 1999년)는 달리는 동안 허벅지 뼈가 높은 굽힘 순간에도 견딜 수 있도록 개조되었다고 주장하면서 카르노타우루스가 빠른 달리기 선수라고 추정했다.그 힘을 견디는 동물의 다리의 능력은 최고 속도를 제한한다.카르노타우루스의 달리기 적응은 타조의 적응에 못 미치지만 인간의 적응보다는 나았을 것이다.[AU][49]과학자들은 카르노타우루스의 최고 속도가 시속 48~56km(30~35mi)에 달했다고 계산한다.[50]
공룡에서는 가장 중요한 기관차 근육이 꼬리 부분에 위치했다.카우도페모랄리스라고 불리는 이 근육은 허벅지 뼈의 두드러진 능선인 네 번째 트로이칸터에 붙어서 수축되면 허벅지 뼈를 뒤로 당긴다.스콧 피플스와 필 커리(2011년)는 카르노타우루스의 꼬리 척추뼈에서 카우달 늑골이 수평으로 돌출되지 않고("T자형") 척추뼈의 수직축에 대해 각도를 이루며 'V'를 형성하고 있다는 점에 주목한다.이것은 다른 어떤 세로포드보다 큰 카우도페모랄리스 근육을 위한 추가 공간을 제공했을 것이다. 즉, 근육량은 다리당 111 ~ 137kg(245 ~ 302lb)으로 계산되었다.따라서 카르노타우루스는 가장 빠른 대형 테로포드 중 하나일 수도 있었다.[28]카우도페모랄리스 근육이 확대된 반면, 카우달 늑골 위에 위치한 후축 근육은 비례적으로 더 작았을 것이다.긴디시무스와 척수근이라고 불리는 이 근육들은 꼬리의 움직임과 안정성을 담당했다.이러한 근육의 감소에도 불구하고 꼬리의 안정성을 유지하기 위해 코달늑골은 앞으로 돌출하는 과정을 통해 척추뼈와 골반이 서로 맞물려 꼬리가 뻣뻣해진다.그 결과 다른 회전목마와는 달리 엉덩이와 꼬리가 동시에 회전해야 했기 때문에 빡빡한 회전을 할 수 있는 능력이 떨어졌을 것이다.[28]
뇌와 감각
세로니와 폴리나-카라바할은 2019년 CT 스캔을 통해 뇌가 들어 있는 내분비강을 연구했다.뇌는 이 공간의 일부분만 채웠을 것이지만, 내분비 충치의 용적은 168.8cm³이었다.저자들은 두 개의 다른 뇌 크기 추정치를 사용했는데, 뇌 크기가 각각 50%와 37%라고 가정했다.이로 인해 파충류 뇌화 지수(지성의 척도)는 관련 마장가사우루스보다 크지만 티라노사우루스에서는 작다.호르몬을 생성하는 소나무샘은 다른 아벨리사우루스보다 작았을지도 모르는데, 이는 솔샘이 위치한 것으로 생각되는 전뇌 위에 있는 공간이다.[24]
후각을 수용하는 후각 전구는 큰 반면 시력을 담당하는 광엽은 상대적으로 작았다.이것은 후각이 시력감각보다 더 발달했을지도 모른다는 것을 나타내는 반면, 현대 조류는 그 반대다.후각과 전구의 앞쪽 끝은 아래쪽으로 휘어져 있었는데, 이는 인디오사우루스만이 공유하는 특징이며, 다른 아벨리사우루스에서는 이러한 구조물이 수평 방향으로 향하였다.세로니, 파울리나-카라바잘에 의해 가설된 바와 같이, 이 하향곡선은 전구의 큰 크기와 함께 카르노타우루스가 다른 아벨리사우루스보다 후각에 더 의존했음을 나타낼지도 모른다.시력 안정화(눈과 몸의 조화)와 상관관계가 있다고 생각되는 뇌엽인 플로쿨루스는 카르노타우루스(Carnotaurus)와 다른 남미 아벨리사우루스(Abelisaurids)에서 컸다.이것은 이러한 형태들이 머리와 몸의 빠른 움직임을 자주 사용했음을 나타낼 수 있다.카르노타우루스나 다른 아벨리사우루스에서는 내이의 짧은 라게나(lagena)로 알 수 있듯이 청력이 제대로 발달되지 않았을지도 모른다.청력 범위는 3kHz 미만으로 추정되었다.[24]
노화 및 연근개념

원래 카르노타우루스가 발견된 바위는 약 1억년(알비안 또는 세노마니아 단계)으로 여겨졌던 고르로 프리지오 형성 상부에 배속되었다.[5][AV]후에, 그들은 캄파니아와 마스트리히트 무대(8360만 년 전 ~ 6600만 년 전)[13]에 이르는 훨씬 더 젊은 라 콜로니아 포메이션과 관련이 있다는 것을 깨달았다.[1]노바스는 2009년 한 책에서 720만 년 전에서 6990만 년 전(하위 마스트리히트 무대)의 더 좁은 시간을 주었다.[AW]따라서 카르노타우루스(Carnotaurus)는 가장 최근에 알려진 남미 아벨리사우루스였다.[28]백악기 후반에 이르러 남미는 이미 아프리카와 북아메리카 양쪽에서 고립되었다.[51]
라 콜로니아 포메이션은 북파타고니아 마시프의 남쪽 경사면 위에 노출되어 있다.[52]카르노타우루스(Carnotaurus)를 포함한 척추동물 화석은 대부분 형성의 중간 부분(미들 페이시스 협회라고 한다)에서 나온다.[52]이 부분은 하구, 갯벌 또는 해안 평야로 이루어진 환경의 퇴적물을 대표할 가능성이 있다.[52]기후는 건조하고 습한 기간과 함께 계절적이었을 것이다.[52]가장 흔한 척추동물로는 세라토돈트 폐어, 거북이, 악어, 플레시오사우르스, 공룡, 도마뱀, 뱀, 포유류가 있다.[53]발견된 뱀 중 일부는 알라미토피스 아르젠티누스와 같은 보아과와 마드소과과에 속한다.[54]거북이는 적어도 5개의 세사로 대표되며, 4개는 첼로과(Pleurodira)에서, 1개는 메이올라니과(Cryptodira)에서 대표된다.[55]해양 파충류 중에는 폴리코틸레과과의 플레시오사우루스 술쿠스쿠스 에레레니도 있다.[55]포유류는 남미 도코돈트 최초의 기록으로 여겨졌던 레이지트리움 부노돈툼과 [52]멀티투버쿨라타일 가능성이 있는 아르헨티나산 콜로니엔시스로 대표된다.[56]2011년 라 콜로니아 형성의 새로운 에반티오르니틴 새 발견이 발표되었다.[57]
참고 항목
메모들
- ^ 페이지 276 노바스(2009)[2]
- ^ 페이지 2 보나파르트 (1990)[4]
- ^ 페이지 2 보나파르트 (1990)[4]
- ^ 페이지 2 보나파르트 (1990)[4]
- ^ 페이지 191 카라노와 샘프슨(2008)[8]
- ^ 페이지 191 카라노와 샘프슨(2008)[8]
- ^ 페이지 3 보나파르트 (1990)[4]
- ^ 페이지 38 보나파르트 (1990)[4]
- ^ P. 162 후아레스 발레리 외 연구진(2010)[18]
- ^ 페이지 191 카라노와 샘프슨(2008)[8]
- ^ P. 162 후아레스 발레리 외 연구진(2010)[18]
- ^ 페이지 163 후아레스 발레리 외 연구진(2010)[18]
- ^ 페이지 556 칼보 외(2004)[20]
- ^ 페이지 191 카라노와 샘프슨(2008)[8]
- ^ 페이지 30 보나파르트 (1990)[4]
- ^ 페이지 187 Mazzetta 외 연구진(1998)[22]
- ^ 페이지 79 Mazzetta 등.(2004)[23]
- ^ 페이지 276 노바스(2009)[2]
- ^ 페이지 28-32 보나파르트 (1990)[4]
- ^ 페이지 8 보나파르트 (1990)[4]
- ^ 페이지 191 카라노와 샘프슨(2008)[8]
- ^ 페이지 4–5 보나파르트 (1990)[4]
- ^ 페이지 5 보나파르트 (1990)[4]
- ^ 페이지 3 보나파르트 (1990)[4]
- ^ 페이지 191 Mazzetta 외 연구진(1998)[22]
- ^ 페이지 255 in Novas(2009)[2]
- ^ 6페이지 보나파르트 (1990)[4]
- ^ 6페이지 보나파르트 (1990)[4]
- ^ 6페이지 보나파르트 (1990)[4]
- ^ 6페이지 보나파르트 (1990)[4]
- ^ 페이지 191 카라노와 샘프슨(2008)[8]
- ^ 페이지 257 노바스(2009)[2]
- ^ 페이지 1276 (Ruiz 등)[29]
- ^ 페이지 32 보나파르트 (1990)[4]
- ^ 페이지 32 보나파르트 (1990)[4]
- ^ 페이지 3 보나파르트 (1990)[4]
- ^ 페이지 264–299(Novas(2009)[2]
- ^ 페이지 264–299(Novas(2009)[2]
- ^ 페이지 32 보나파르트 (1990)[4]
- ^ 페이지 276–277 in Novas(2009)[2]
- ^ Novas(2009)에서 페이지 256–[2]261
- ^ 페이지 188–189 및 카라노 및 샘프슨(2008)[8]
- ^ 페이지 202 카라노와 샘프슨(2008)[8]
- ^ 페이지 202 카라노와 샘프슨(2008)[8]
- ^ 페이지 259–261 노바스(2009)[2]
- ^ 페이지 260–261 노바스(2009)[2]
- ^ 페이지 186 및 190, Mazzetta 등(1998)[22]
- ^ 페이지 3 보나파르트 (1990)[4]
- ^ 페이지 276 노바스(2009)[2]
참조
- ^ a b c d e f g h i j k l m n o p q r s t u Cerroni, M. A.; Canale, J. I.; Novas, F. E. (2020). "The skull of Carnotaurus sastrei Bonaparte 1985 revisited: insights from craniofacial bones, palate and lower jaw". Historical Biology: 1–42. doi:10.1080/08912963.2020.1802445.
- ^ a b c d e f g h i j k Novas, Fernando E. (2009). The age of dinosaurs in South America. Bloomington: Indiana University Press. ISBN 978-0-253-35289-7.
- ^ a b Salgado, Leonardo; Bonaparte, José F. (1991). "Un nuevo sauropodo Dicraeosauridae, Amargasaurus cazaui gen. et sp. nov., de la Formacion La Amarga, Neocomiano de la Provincia del Neuquén, Argentina". Ameghiniana (in Spanish). 28 (3–4): 334.
- ^ a b c d e f g h i j k l m n o p q r s t u v w x Bonaparte, José F.; Novas, Fernando E.; Coria, Rodolfo A. (1990). "Carnotaurus sastrei Bonaparte, the horned, lightly built carnosaur from the Middle Cretaceous of Patagonia" (PDF). Contributions in Science. 416: 1–41. Archived from the original (PDF) on July 21, 2010.
- ^ a b c d e Bonaparte, José F. (1985). "A horned Cretaceous carnosaur from Patagonia". National Geographic Research. 1 (1): 149–151.
- ^ a b Paulina Carabajal, Ariana (2011). "The braincase anatomy of Carnotaurus sastrei (Theropoda: Abelisauridae) from the Upper Cretaceous of Patagonia". Journal of Vertebrate Paleontology. 31 (2): 378–386. doi:10.1080/02724634.2011.550354. S2CID 129933997.
- ^ a b c d e f g h i Czerkas, Stephen A.; Czerkas, Sylvia J. (1997). "The Integument and Life Restoration of Carnotaurus". In Wolberg, D. I.; Stump, E.; Rosenberg, G. D. (eds.). Dinofest International. Academy of Natural Sciences, Philadelphia. pp. 155–158.
- ^ a b c d e f g h i Carrano, Matthew T.; Sampson, Scott D. (January 2008). "The Phylogeny of Ceratosauria (Dinosauria: Theropoda)". Journal of Systematic Palaeontology. 6 (2): 183–236. doi:10.1017/S1477201907002246. S2CID 30068953.
- ^ a b c d Paul, Gregory S. (1988). Predatory Dinosaurs of the World. pp. 284–285. ISBN 978-0-671-61946-6.
- ^ Yong, Ed (October 18, 2011). "Butch tail made Carnotaurus a champion dinosaur sprinter". National Geographic. Retrieved July 31, 2019.
- ^ Headden, Jaime A. (September 19, 2006). "Re: Carnotaurus sastrei etymology". Dinosaur Mailing List.
- ^ Bonaparte, José F. (1991). "The gondwanian theropod families Abelisauridae and Noasauridae". Historical Biology. 5: 1. doi:10.1080/10292389109380385.
- ^ a b Bonaparte, José F. (1996). "Cretaceous tetrapods of Argentina". Münchener Geowissenschaftliche Abhandlung. A (30): 89.
- ^ Glut, Donald F. (1997). "Carnotaurus". Dinosaurs, the encyclopedia. Jefferson, North Carolina: McFarland & Company, Inc. Publishers. pp. 256–259. ISBN 978-0-375-82419-7.
- ^ a b Glut, Donald F. (2003). "Carnotaurus". Dinosaurs: The Encyclopedia. 3rd Supplement. Jefferson, North Carolina: McFarland & Company, Inc. pp. 274–276. ISBN 978-0-7864-1166-5.
- ^ Glut, Donald F. (2000). "Carnotaurus". Dinosaurs: The Encyclopedia. 1st Supplement. Jefferson, North Carolina: McFarland & Company, Inc. pp. 165–167. ISBN 978-0-7864-0591-6.
- ^ a b Candeiro, Carlos Roberto dos Anjos; Martinelli, Agustín Guillermo. "Abelisauroidea and carchardontosauridae (theropoda, dinosauria) in the cretaceous of south america. Paleogeographical and geocronological implications". Uberlândia. 17 (33): 5–19.
- ^ a b c Juárez Valieri, Rubén D.; Porfiri, Juan D.; Calvo, Jorge O. (2010). "New information on Ekrixinatosaurus novasi Calvo et al. 2004, a giant and massively-constructed Abelisauroid from the 'Middle Cretaceous' of Patagonia". Paleontologıa y Dinosaurios en América Latina: 161–169.
- ^ a b c Paul, Gregory S. (2010). The Princeton Field Guide to Dinosaurs (1st ed.). Princeton University Press. ISBN 9780691137209.
- ^ Calvo, Jorge O.; Rubilar-Rogers, David; Moreno, Karen (2004). "A new Abelisauridae (Dinosauria: Theropoda) from northwest Patagonia". Ameghiniana. 41 (4): 555–563.
- ^ Grillo, O.N.; Delcourt, R. (2016). "Allometry and body length of abelisauroid theropods: Pycnonemosaurus nevesi is the new king". Cretaceous Research. 69: 71–89. doi:10.1016/j.cretres.2016.09.001.
- ^ a b c d e f g Mazzetta, Gerardo V.; Fariña, Richard A.; Vizcaíno, Sergio F. (1998). "On the palaeobiology of the South American horned theropod Carnotaurus sastrei Bonaparte" (PDF). Gaia. 15: 185–192.
- ^ a b Mazzetta, Gerardo V.; Christiansen, Per; Fariña, Richard A. (June 2004). "Giants and Bizarres: Body Size of Some Southern South American Cretaceous Dinosaurs". Historical Biology. 16 (2–4): 71–83. CiteSeerX 10.1.1.694.1650. doi:10.1080/08912960410001715132. S2CID 56028251.
- ^ a b c d Cerroni, Mauricio A.; Paulina-Carabajal, Ariana (2019). "Novel information on the endocranial morphology of the abelisaurid theropod Carnotaurus sastrei". Comptes Rendus Palevol. 18 (8): 985–995. doi:10.1016/j.crpv.2019.09.005.
- ^ a b Sampson, Scott D.; Witmer, Lawrence M. (2007). "Craniofacial Anatomy of Majungasaurus crenatissimus (Theropoda: Abelisauridae) From the Late Cretaceous of Madagascar". Journal of Vertebrate Paleontology. 27 (sp8): 95–96. doi:10.1671/0272-4634(2007)27[32:CAOMCT]2.0.CO;2.
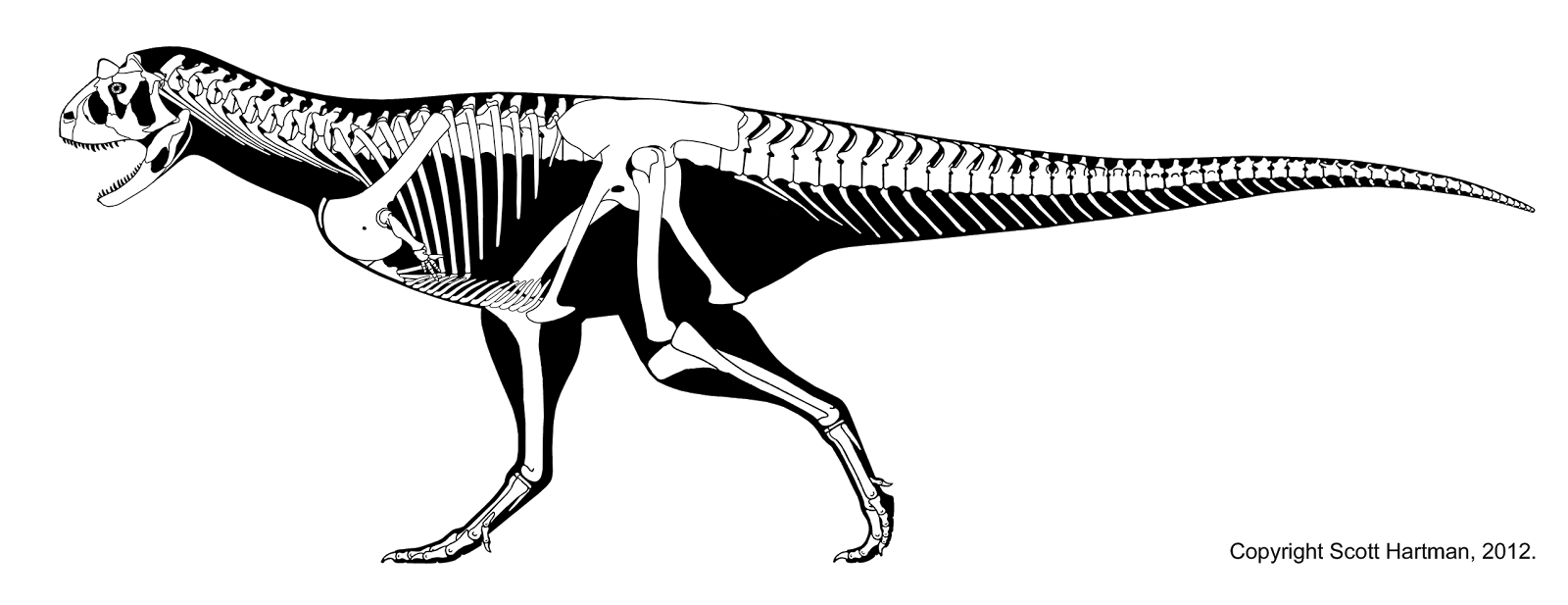
- ^ a b c Hartman, Scott (2012). "Carnotaurus – delving into self-parody?". Retrieved December 7, 2012.
- ^ Méndez, Ariel (2014). "The cervical vertebrae of the Late Cretaceous abelisaurid dinosaur Carnotaurus sastrei". Acta Palaeontologica Polonica. 59 (1): 99–107. doi:10.4202/app.2011.0129. S2CID 67792084.
- ^ a b c d e Persons, W.S.; Currie, P.J. (2011). Farke, Andrew Allen (ed.). "Dinosaur Speed Demon: The caudal musculature of Carnotaurus sastrei and implications for the evolution of South American abelisaurids". PLOS ONE. 6 (10): e25763. Bibcode:2011PLoSO...625763P. doi:10.1371/journal.pone.0025763. PMC 3197156. PMID 22043292.
- ^ a b c d Ruiz, Javier; Torices, Angélica; Serrano, Humberto; López, Valle (2011). "The hand structure of Carnotaurus sastrei (Theropoda, Abelisauridae): implications for hand diversity and evolution in abelisaurids" (PDF). Palaeontology. 54 (6): 1271–1277. doi:10.1111/j.1475-4983.2011.01091.x.
- ^ Agnolin, Federico L.; Chiarelli, Pablo (June 2010). "The position of the claws in Noasauridae (Dinosauria: Abelisauroidea) and its implications for abelisauroid manus evolution". Paläontologische Zeitschrift. 84 (2): 293–300. doi:10.1007/s12542-009-0044-2. S2CID 84491924.
- ^ Senter, P. (2010). "Vestigial skeletal structures in dinosaurs". Journal of Zoology. 280 (4): 60–71. doi:10.1111/j.1469-7998.2009.00640.x.
- ^ Campione, Nicolás E.; Barrett, Paul M.; Evans, David C. (2020). "On the ancestry of feathers in Mesozoic dinosaurs". In Christian Foth; Oliver W.M. Rauhut (eds.). The Evolution of Feathers. Springer. pp. 213–243. ISBN 978-3-030-27222-7.
- ^ Hendrickx, Christophe; Bell, Phil R. (August 2021). "The scaly skin of the abelisaurid Carnotaurus sastrei (Theropoda: Ceratosauria) from the Upper Cretaceous of Patagonia". Cretaceous Research: 104994. doi:10.1016/j.cretres.2021.104994.
- ^ a b c Canale, Juan I.; Scanferla, Carlos A.; Agnolin, Federico; Novas, Fernando E. (2009). "New carnivorous dinosaur from the Late Cretaceous of NW Patagonia and the evolution of abelisaurid theropods". Naturwissenschaften. 96 (3): 409–14. Bibcode:2009NW.....96..409C. doi:10.1007/s00114-008-0487-4. hdl:11336/52024. PMID 19057888. S2CID 23619863.
- ^ a b Coria, Rodolfo A.; Chiappe, Luis M.; Dingus, Lowell (2002). "A new close relative of Carnotaurus sastrei Bonaparte 1985 (Theropoda: Abelisauridae) from the Late Cretaceous of Patagonia". Journal of Vertebrate Paleontology. 22 (2): 460. doi:10.1671/0272-4634(2002)022[0460:ANCROC]2.0.CO;2.
- ^ Ezcurra, Martín D.; Agnolin, Federico L.; Novas, Fernando E. (May 10, 2010). "An abelisauroid dinosaur with a non-atrophied manus from the Late Cretaceous Pari Aike Formation of southern Patagonia". Zootaxa. 2450 (1): 1. doi:10.11646/zootaxa.2450.1.1.
- ^ a b c Delcourt, Rafael (2018). "Ceratosaur palaeobiology: new insights on evolution and ecology of the southern rulers". Scientific Reports. 8 (1): 9730. Bibcode:2018NatSR...8.9730D. doi:10.1038/s41598-018-28154-x. PMC 6021374. PMID 29950661.
- ^ Sereno, Paul C.; Wilson, Jeffrey A.; Conrad, Jack L. (July 7, 2004). "New dinosaurs link southern landmasses in the Mid-Cretaceous". Proceedings of the Royal Society B: Biological Sciences. 271 (1546): 1325–1330. doi:10.1098/rspb.2004.2692. PMC 1691741. PMID 15306329.
- ^ Tykoski, Ronald B.; Rowe, Timothy (2004). "Ceratosauria". In Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (eds.). The Dinosauria (Second ed.). Berkeley: University of California Press. p. 65. ISBN 978-0-520-24209-8.
- ^ Wilson, Jeffrey A.; Sereno, Paul C.; Srivastava, Suresh; Bhatt, Devendra K.; Khosla, Ashu; Sahni, Ashok (2003). "A new abelisaurid (Dinosauria, Theropoda) from the Lameta Formation (Cretaceous, Maastrichtian) of India". Contributions from the Museum of Paleontology. 31 (1): 25. hdl:2027.42/48667.
- ^ Wang, Shuo; Stiegler, Josef; Amiot, Romain; Wang, Xu; Du, Guo-hao; Clark, James M.; Xu, Xing (January 2017). "Extreme Ontogenetic Changes in a Ceratosaurian Theropod". Current Biology. 27 (1): 144–148. doi:10.1016/j.cub.2016.10.043. PMID 28017609.
- ^ Sereno, Paul (2005). "Carnotaurinae". Taxon Search. Archived from the original on May 16, 2012. Retrieved December 29, 2012.
- ^ Sereno, Paul (2005). "Carnotaurini". Taxon Search. Archived from the original on May 16, 2012. Retrieved December 29, 2012.
- ^ a b c d e f g h Mazzetta, Gerardo V.; Cisilino, Adrián P.; Blanco, R. Ernesto; Calvo, Néstor (2009). "Cranial mechanics and functional interpretation of the horned carnivorous dinosaur Carnotaurus sastrei". Journal of Vertebrate Paleontology. 29 (3): 822–830. doi:10.1671/039.029.0313. S2CID 84565615.
- ^ Chure, Daniel J. (1998). "On the orbit of theropod dinosaurs". Gaia. 15: 233–240.
- ^ Snively, Eric; Cotton, John R.; Witmer, Lawrence; Ridgely, Ryan; Theodor, Jessica (2011). "Finite element comparison of cranial sinus function in the dinosaur Majungasaurus and head-clubbing giraffes". Summer Bioengineering Conference. Vol. 54587. American Society of Mechanical Engineers. pp. 1075–1076.
- ^ Therrien, François; Henderson, Donald; Ruff, Christopher (2005). "Bite Me – Biomechanical Models of Theropod Mandibles and Implications for Feeding Behavior". In Carpenter, Kenneth (ed.). The carnivorous dinosaurs. Indiana University Press. pp. 179–198, 228. ISBN 978-0-253-34539-4.
- ^ Bakker, Robert T. (1998). "Brontosaur killers: Late Jurassic allosaurids as sabre-tooth cat analogues" (PDF). Gaia. 15: 145–158.
- ^ Mazzetta, Gerardo V.; Farina, Richard A. (1999). "Estimacion de la capacidad atlética de Amargasaurus cazaui Salgado y Bonaparte, 1991, y Carnotaurus sastrei Bonaparte, 1985 (Saurischia, Sauropoda-Theropoda)". XIV Jornadas Argentinas de Paleontologia de Vertebrados, Ameghiniana (in Spanish). 36 (1): 105–106.
- ^ "Predatory dinosaur was fearsomely fast". CBC News. October 21, 2011. Retrieved April 22, 2017.
- ^ Le Loeuff, Jean (1997). "Biogeography". In Padian, Kevin; Currie, Philip J. (eds.). Encyclopedia of dinosaurs. San Diego: Academic Press. pp. 51–56. ISBN 978-0-12-226810-6.
- ^ a b c d e Pascual, Rosendo; Goin, Francisco J.; González, Pablo; Ardolino, Alberto; Puerta, Pablo F. (2000). "A highly derived docodont from the Patagonian Late Cretaceous: evolutionary implications for Gondwanan mammals". Geodiversitas. 22 (3): 395–414.
- ^ Sterli, Juliana; De la Fuente, Marcelo S. (2011). "A new turtle from the La Colonia Formation (Campanian–Maastrichtian), Patagonia, Argentina, with remarks on the evolution of the vertebral column in turtles". Palaeontology. 54 (1): 65. doi:10.1111/j.1475-4983.2010.01002.x.
- ^ Albino, Adriana M. (2000). "New record of snakes from the Cretaceous of Patagonia (Argentina)". Geodiversitas. 22 (2): 247–253.
- ^ a b Gasparini, Zulma; De la Fuente, Marcelo (2000). "Tortugas y Plesiosaurios de la Formación La Colonia (Cretácico Superior) de Patagonia, Argentina". Revista Española de Paleontología (in Spanish). 15 (1): 23.
- ^ Kielan−Jaworowska, Zofia; Ortiz−Jaureguizar, Edgardo; Vieytes, Carolina; Pascual, Rosendo; Goin, Francisco J. (2007). "First ?cimolodontan multi−tuberculate mammal from South America" (PDF). Acta Palaeontologica Polonica. 52 (2): 257–262.
- ^ Lawver, Daniel R.; Debee, Aj M.; Clarke, Julia A.; Rougier, Guillermo W. (January 1, 2011). "A New Enantiornithine Bird from the Upper Cretaceous La Colonia Formation of Patagonia, Argentina". Annals of Carnegie Museum. 80 (1): 35–42. doi:10.2992/007.080.0104. S2CID 85015365.
외부 링크
| 위키미디어 커먼스는 카르노타우루스 관련 매체를 보유하고 있다. |
- Nacional de Mar del Plata의 카르노타우루스에게 물린 상처.(스페인어)
- 스콧 하트먼의 골격 재건














| 국립도서관 | |
|---|---|
| 기타 | |


{kind=link}