플라스모데스마
Plasmodesma
플라스모데스마타(plasmodesma)는 식물[2] 세포와 일부 조류 세포의 세포벽을 가로지르는 미세한 통로이며, 그들 사이의 이동과 통신을 가능하게 한다.플라스모데스마타는 여러 [3]계통에서 독립적으로 진화했고, 이러한 구조를 가진 종들은 육지식물로 [4]더 잘 알려진 모든 태생식물뿐만 아니라 Charophyceae, Charales, Coleochaetales 그리고 Phaeophyceae의 구성원들을 포함합니다.동물 세포와 달리, 거의 모든 식물 세포는 다당 세포벽으로 둘러싸여 있다.따라서 인접한 식물세포는 한 쌍의 세포벽과 중간 층에 의해 분리되어 아포플라스트로 알려진 세포외 도메인을 형성한다.세포벽은 작은 수용성 단백질과 다른 용질에도 투과할 수 있지만, 플라스모데스마타는 세포들 사이의 물질의 직접적이고, 조절되고, 상징적인 운송을 가능하게 합니다.플라스모데스마타에는 두 가지 형태가 있다: 세포 분열 중에 형성되는 1차 플라스모데스마타와 성숙한 [5]세포 사이에 형성될 수 있는 2차 플라스모데스마타.
갭[6] 접합과 막 나노튜브라고 불리는 유사한 구조는 식물 [8]세포에서 플라스티드 사이에 동물[7] 세포와 스트로뮬을 상호 연결합니다.
형성
새로운 세포벽이 2개의 새롭게 분할된 식물세포 사이에서 합성됨에 따라 내소체 일부가 중간 층을 가로질러 포착되었을 때 1차 플라스모데마타가 형성된다.이것들은 결국 세포들 사이의 세포질 결합이 된다.형성 장소에서는 벽이 더 두꺼워지지 않고 벽 안에 움푹 패인 곳 또는 피트라고 불리는 얇은 영역이 형성된다.구덩이는 보통 인접한 세포들 사이에 짝을 이룬다.플라스모데스마타는 또한 비분열 세포 사이의 기존 세포벽에 삽입될 수 있다(2차 플라스모데스마타).[9]
프라이머리 플라스마데마타
1차 플라스모데스마타의 형성은 세포분열 과정 중 내소체와 새로운 판이 융합되는 과정에서 발생하며, 이 과정은 세포질 기공(또는 세포질 슬리브)을 형성한다.압박 ER로도 알려진 데스모튜브는 피질 ER과 함께 형성됩니다.압축된 ER과 피질 ER은 모두 촘촘히 채워져 있어 내강 공간을 확보할 수 없습니다.압착된 ER이 플라스마드마타에서 막수송 경로로 기능하는 것이 제안된다.피질 ER의 필라멘트가 새로운 세포판의 형성에 얽히면 육지 식물에서 플라스모데스마타 형성이 일어난다.억제된 ER은 성장하는 세포벽의 압력과 ER 및 PM 단백질의 상호작용의 조합에 의해 형성된다는 가설이 있다.1차 플라스모데마타는 세포벽이 얇아 보이는 영역에 종종 존재한다.이는 세포벽이 팽창함에 따라 1차 플라스마마의 양이 감소하기 때문이다.세포벽 성장 중 플라스모데말 밀도를 더욱 확대하기 위해 2차 플라스모데마타를 제조한다.2차 플라스모데스마타 형성의 과정은 아직 완전히 이해되어야 하지만, 다양한 분해 효소와 ER 단백질은 [10]이 과정을 자극한다고 알려져 있다.
구조.
플라스모데스마탈 플라즈마막
일반적인 식물 세포는 1,000~100,000개의 플라스모데마타를 가지며 [12][failed verification]µm당2 1~10개의 인접 세포와[11] 접속할 수 있다.플라스모데스마타는 중간점의 직경이 약 50-60 nm이고 세 개의 주요 층, 즉 혈장막, 세포질 슬리브 및 [11]탈모관으로 구성됩니다.그들은 90 nm [12]두께의 세포벽을 가로지를 수 있다.
플라스마 막 부분은 세포막 또는 플라스마 막의 연속적인 확장이며 유사한 인지질 이중층 [13]구조를 가지고 있다.
세포질 슬리브는 플라스마로 둘러싸인 액체로 채워진 공간이며 세포질의 연속적인 확장입니다.플라스마데마타를 통한 분자와 이온의 밀매는 이 공간을 통해 일어난다.더 작은 분자(예: 설탕 및 아미노산)와 이온은 추가적인 화학 에너지 없이도 확산에 의해 플라스마드마타를 쉽게 통과할 수 있다.단백질(예: 녹색 형광 단백질)과 RNA를 포함한 더 큰 분자도 세포질 슬리브를 [14]확산시켜 통과할 수 있습니다.일부 더 큰 분자의 플라스모데스매탈 수송은 현재 알려지지 않은 메커니즘에 의해 촉진된다.플라스모데스마타의 투과성을 조절하는 하나의 메커니즘은 목 부위 주위에 다당류 카르로스를 축적하여 칼라를 형성함으로써 물질의 [13]수송에 이용 가능한 모공의 직경을 감소시키는 것이다.확장에 의해 플라스모데즈마타의 투과성을 증가시키는 활성 게이트 또는 구조 리모델링.플라스모데즈마타 모공 투과성의 증가는 신호 분자, 전사 인자 및 RNA-단백질 복합체와 같은 더 큰 분자 또는 고분자를 다양한 세포 [10]구획으로 운반할 수 있게 합니다.
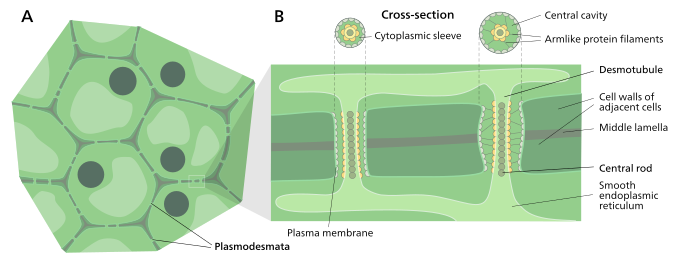
플라스모데스마타의 구조와 식물 세포 내에서의 위치

플라스모데즈마타는 분자가 심플라스틱 경로를 통해 식물 세포 사이를 이동할 수 있도록 한다.


데스모튜브
데스모티큘은 두 개의 인접한 [15]세포 사이를 흐르는 억제된(평탄화된) 소포체의 튜브입니다.일부 분자는 이 [16]채널을 통해 운반되는 것으로 알려져 있지만, 플라스마 물질 운반의 주요 경로는 아닌 것으로 생각된다.
데스모터블 주변과 전자 밀도가 높은 물질의 플라즈마 멤브레인 영역은 종종 플라스모데마를 작은 채널로 [15]분할하는 것처럼 보이는 스포크 같은 구조에 의해 결합됩니다.이러한 구조는 세포 골격의 일부인 미오신과[17][18][19] [18][20]액틴으로 구성될 수 있다.이 경우 이 단백질은 두 세포 사이의 큰 분자의 선택적 운반에 사용될 수 있다.
운송

플라스모데스마타는 단백질(전사인자 포함), 짧은 간섭 RNA, 메신저 RNA, 비로이드, 바이러스 게놈을 세포에서 세포로 운반하는 것으로 나타났다.바이러스 이동 단백질의 한 예는 담배 모자이크 바이러스 MP-30이다. MP-30은 바이러스 자신의 게놈에 결합해 플라스마데마타를 [14]통해 감염된 세포에서 감염되지 않은 세포로 이동시키는 것으로 생각된다.개화 Locus T단백질은 플라즈마드마타를 통해 잎에서 정점근막으로 이동해 [21]개화를 시작한다.
플라즈모데스마타는 인골세포에서도 사용되며, 심플라스틱 트랜스포트(symplastic transport)는 동반세포에 [citation needed]의한 체관세포의 조절에 사용된다.
플라스마드마타를 통과할 수 있는 분자의 크기는 크기 제외 한계에 의해 결정됩니다.이 제한은 매우 가변적이며 능동적으로 [5]변경될 수 있습니다.예를 들어 MP-30은 크기 제외 한계를 700 Daltons에서 9400 Daltons로 늘릴 수 있어 [22]플랜트를 통한 이동을 지원할 수 있다.또한 주입 또는 냉유도에 의해 세포질 내 칼슘 농도가 증가하면 주변 플라스마 마타의 개구부가 좁아져 [23]수송이 제한되는 것으로 나타났다.
플라스모데스마타를 통한 가능한 활성 수송을 위한 몇 가지 모델이 존재합니다.이러한 수송은 데스모터블에 국소화된 단백질과의 상호작용 및/또는 부분적으로 전개되는 단백질의 샤페론과의 상호작용에 의해 매개되어 좁은 통로를 통과할 수 있도록 하는 것이 제안되었다.유사한 메커니즘은 플라스마데마타를 [24][unreliable source?]통해 바이러스 핵산을 운반하는 데 관여할 수 있다.
플라스마 모드마에 걸친 수송을 추정하기 위해 다수의 수학적 모델이 제안되었다.이 모델들은 주로 운송을 확산 문제로 취급하고 있으며, 일부 장애물이 추가되어 있다.[25][26][27]
플라스모데스마타의 세포골격 구성 요소
플라스모데스마는 식물의 거의 모든 세포를 연결하는데, 이것은 바이러스의 확산과 같은 부정적인 영향을 일으킬 수 있다.이것을 이해하기 위해서 우리는 먼저 액틴 미세 필라멘트, 미세관, 그리고 미오신 단백질과 같은 세포 골격 요소들과 그것들이 세포와 어떻게 관련이 있는지 살펴봐야 한다.액틴 마이크로필라멘트는 플라스모데마타를 통한 세포 이동을 가능하게 하는 플라스모데마타에 대한 바이러스 이동 단백질의 운반과 연결되어 있다.담뱃잎의 공동발현을 위한 형광태깅은 액틴 필라멘트가 플라스마드마타로 바이러스 이동 단백질을 운반하는 역할을 한다는 것을 보여주었다.액틴 중합이 차단되면 담배의 이동 단백질을 대상으로 하는 플라스모데스마타의 감소를 유발하고 담배 중엽 세포 사이를 10kDa(16-kDa가 아닌) 성분이 이동할 수 있었다.이것은 또한 담배 식물 [28]내 분자의 세포 대 세포 이동에 영향을 미쳤다.
바이러스
바이러스는 식물 내에서 움직이기 위해 플라스마데마타 채널 내의 액틴 필라멘트를 분해한다.예를 들어, 오이 모자이크 바이러스가 식물에 들어갔을 때, 그것은 플라스마 마타를 통해 그들 자신을 운반하기 위해 바이러스 이동 단백질을 이용하여 거의 모든 세포를 통과할 수 있다.담뱃잎을 액틴 필라멘트를 안정시키는 약인 팔로이딘으로 처리하면 오이모자이크 바이러스 이동 단백질은 플라스모데스마타 크기 제외 한계(SEL)[28]를 높일 수 없다.
묘신
많은 양의 미오신 단백질이 플라스마데마타 부위에서 발견됩니다.이 단백질들은 바이러스 화물을 플라스마데마로 유도하는 데 관여한다.담배 식물에서 돌연변이 형태의 미오신을 검사했을 때 플라스모데즈마를 표적으로 하는 바이러스 단백질이 부정적인 영향을 받았다.약물에 의해 유도된 미오신의 액틴에 대한 영구 결합은 세포 대 세포 이동의 감소를 야기했다.바이러스는 또한 선택적으로 미오신 [28]단백질과 결합할 수 있다.
미소관
미세관은 바이러스 RNA의 세포 전달에 중요한 역할을 한다. 바이러스는 세포에서 세포로 자신을 운반하는 많은 다른 방법들과 미세관을 통해 플라스모데마타에 국소화하기 위해 RNA의 N 말단 도메인을 연관짓는 방법들 중 하나를 사용한다.고온에서 보관된 담배 모자이크 바이러스가 주입된 담배 식물에서는 GFP 라벨이 부착된 TMV 이동 단백질과 미세관 사이에 강한 상관관계가 있었다.이것은 담배를 [28]통한 바이러스 RNA의 확산으로 이어졌다.
플라스모데스마타와 굳은살
플라스모데스마타 조절 및 구조는 굳은살로 알려진 베타 1,3-글루칸 폴리머에 의해 조절된다.굳은살은 세포판의 세포에서 발견되지만, 이 과정이 완료됨에 따라 굳은살 수치는 [citation needed]감소한다.세포에서 굳은살이 풍부한 유일한 부분은 플라스마데마타가 존재하는 세포벽의 부분을 포함합니다.플라스모데스마를 통해 운반되는 것을 조절하기 위해서는 굳은살이 존재해야 한다.캘로스는 플라스마데마타 투과성을 조절하는 메커니즘을 제공한다.서로 다른 조직 간에 수송되는 것을 제어하기 위해 플라스모데스마는 몇 가지 특수한 구조 변화를 [10]겪는다.
플라스모데스마타의 활동은 식물 내의 생리적, 발달적 과정과 관련이 있다.1차 세포 신호를 플라스마데마타를 통해 전달하는 호르몬 신호 경로가 있습니다.또한 플라스모데스마타 기능과 관련된 환경적, 생리학적, 발달적 단서의 패턴도 있다.플라스모데스마타의 중요한 메커니즘은 채널을 게이트하는 능력이다.굳은살은 플라스마데마타 조리개 [29]크기를 변경하는 방법인 것으로 입증되었습니다.굳은살 퇴적물은 형성된 새로운 세포벽의 플라스모데스마타의 목에서 발견됩니다.플라스모데스마타의 퇴적물 수준은 변동할 수 있으며, 이는 플라스모데스마타에 굳은살이 축적되어 플라스모데스마타가 게이트 또는 더 열리게 하는 신호가 있음을 나타냅니다.베타 1,3-글루칸 합성효소 및 가수분해효소의 효소 활성은 플라스마데마타 셀룰로오스 수준의 변화에 관여한다.일부 세포외 신호는 이 합성효소와 가수분해효소의 활성 전사를 변화시킨다.아라비도시스 탈리아나는 B-1,3-글루칸의 촉매 서브유닛을 코드하는 굳은살 합성효소 유전자를 가지고 있다.이 유전자 풀에서 기능 돌연변이의 이득은 [28]혈장에서의 굳은살 침착의 증가와 발달 중의 고분자 수송의 감소를 나타낸다.
「 」를 참조해 주세요.
레퍼런스
- ^ Maule, Andrew (December 2008). "Plasmodesmata: structure, function and biogenesis". Current Opinion in Plant Biology. 11 (6): 680–686. doi:10.1016/j.pbi.2008.08.002. PMID 18824402.
- ^ Oparka, K. J. (2005). Plasmodesmata. Blackwell Pub Professional. ISBN 978-1-4051-2554-3.
- ^ Zoë A. Popper; Gurvan Michel; Cécile Hervé; David S. Domozych; William G.T. Willats; Maria G. Tuohy; Bernard Kloareg; Dagmar B. Stengel (2011). "Evolution and Diversity of Plant Cell Walls: From Algae to Flowering Plants" (PDF). Annual Review of Plant Biology. 62: 567–590. doi:10.1146/annurev-arplant-042110-103809. hdl:10379/6762. PMID 21351878. S2CID 11961888.
- ^ Graham, LE; Cook, ME; Buse, JS(2000), 국립과학원회보 97, 4535-4540.
- ^ a b Jan Traas; Teva Vernoux (29 June 2002). "The shoot apical meristem: the dynamics of a stable structure". Philosophical Transactions of the Royal Society B: Biological Sciences. 357 (1422): 737–747. doi:10.1098/rstb.2002.1091. PMC 1692983. PMID 12079669.
- ^ Bruce Alberts (2002). Molecular Biology of the Cell (4th ed.). New York: Garland Science. ISBN 978-0-8153-3218-3.
- ^ Gallagher KL, Benfey PN (15 January 2005). "Not just another hole in the wall: understanding intercellular protein trafficking". Genes & Development. 19 (2): 189–95. doi:10.1101/gad.1271005. PMID 15655108.
- ^ Gray JC, Sullivan JA, Hibberd JM, Hansen MR (2001). "Stromules: mobile protrusions and interconnections between plastids". Plant Biology. 3 (3): 223–33. doi:10.1055/s-2001-15204.
- ^ Lucas, W.; Ding, B.; Van der Schoot, C. (1993). "Tansley Review No.58 Plasmodesmata and the supracellular nature of plants". New Phytologist. 125 (3): 435–476. doi:10.1111/j.1469-8137.1993.tb03897.x. JSTOR 2558257. PMID 33874589.
- ^ a b c Sager, Ross (June 7, 2018). "Plasmodesmata at a Glance". Journal of Cell Science. 131 (11): jcs209346. doi:10.1242/jcs.209346. PMID 29880547.
- ^ a b Robards, AW (1975). "Plasmodesmata". Annual Review of Plant Physiology. 26: 13–29. doi:10.1146/annurev.pp.26.060175.000305.
- ^ a b Lodish, Berk, Zipursky, Matsudaira, Baltimore, Darnell (2000). "22". Molecular Cell Biology (4 ed.). pp. 998. ISBN 978-0-7167-3706-3. OCLC 41266312.
{{cite book}}: CS1 maint: 작성자 파라미터 사용(링크) - ^ a b AW Robards (1976). "Plasmodesmata in higher plants". In BES Gunning; AW Robards (eds.). Intercellular communications in plants: studies on plasmodesmata. Berlin: Springer-Verlag. pp. 15–57.
- ^ a b A. G. Roberts; K. J. Oparka (1 January 2003). "Plasmodesmata and the control of symplastic transport". Plant, Cell & Environment. 26 (1): 103–124. doi:10.1046/j.1365-3040.2003.00950.x.
- ^ a b Overall, RL; Wolfe, J; Gunning, BES (1982). "Intercellular communication in Azolla roots: I. Ultrastructure of plasmodesmata". Protoplasma. 111 (2): 134–150. doi:10.1007/bf01282071. S2CID 5970113.
- ^ Cantrill, LC; Overall, RL; Goodwin, PB (1999). "Cell-to-cell communication via plant endomembranes". Cell Biology International. 23 (10): 653–661. doi:10.1006/cbir.1999.0431. PMID 10736188. S2CID 23026878.
- ^ Radford, JE; White, RG (1998). "Localization of a myosin‐like protein to plasmodesmata". Plant Journal. 14 (6): 743–750. doi:10.1046/j.1365-313x.1998.00162.x. PMID 9681037.
- ^ a b Blackman, LM; Overall, RL (1998). "Immunolocalisation of the cytoskeleton to plasmodesmata of Chara corallina". Plant Journal. 14 (6): 733–741. doi:10.1046/j.1365-313x.1998.00161.x.
- ^ Reichelt, S; Knight, AE; Hodge, TP; Baluska, F; Samaj, J; Volkmann, D; Kendrick-Jones, J (1999). "Characterization of the unconventional myosin VIII in plant cells and its localization at the post-cytokinetic cell wall". Plant Journal. 19 (5): 555–569. doi:10.1046/j.1365-313x.1999.00553.x. PMID 10504577.
- ^ White, RG; Badelt, K; Overall, RL; Vesk, M (1994). "Actin associated with plasmodesmata". Protoplasma. 180 (3–4): 169–184. doi:10.1007/bf01507853. S2CID 9767392.
- ^ Corbesier, L., Vincent, C., Jang, S., Fornara, F., Fan, Q.; et al. (2007). "FT protein movement contributes to long distance signalling in floral induction of Arabidopsis". Science. 316 (5827): 1030–1033. Bibcode:2007Sci...316.1030C. doi:10.1126/science.1141752. hdl:11858/00-001M-0000-0012-3874-C. PMID 17446353. S2CID 34132579.
{{cite journal}}: CS1 maint: 여러 이름: 작성자 목록(링크) - ^ Shmuel, Wolf; William, J. Lucas; Carl, M. Deom (1989). "Movement Protein of Tobacco Mosaic Virus Modifies Plasmodesmatal Size Exclusion Limit". Science. 246 (4928): 377–379. Bibcode:1989Sci...246..377W. doi:10.1126/science.246.4928.377. PMID 16552920. S2CID 2403087.
- ^ Aaziz, R.; Dinant, S.; Epel, B. L. (1 July 2001). "Plasmodesmata and plant cytoskeleton". Trends in Plant Science. 6 (7): 326–330. doi:10.1016/s1360-1385(01)01981-1. ISSN 1360-1385. PMID 11435172.
- ^ 식물생리학 강의, 5장 2010-02-16 Wayback Machine 아카이브
- ^ Rutschow, Heidi L.; Baskin, Tobias I.; Kramer, Eric M. (2011-04-01). "Regulation of Solute Flux through Plasmodesmata in the Root Meristem". Plant Physiology. 155 (4): 1817–1826. doi:10.1104/pp.110.168187. ISSN 0032-0889. PMC 3091107. PMID 21325566.
- ^ Deinum, Eva E; Mulder, Bela M; Benitez-Alfonso, Yoselin (2019-11-22). Hardtke, Christian S; Bergmann, Dominique C; Liesche, Johannes; Knoblauch, Michael; Couvreur, Valentin (eds.). "From plasmodesma geometry to effective symplasmic permeability through biophysical modelling". eLife. 8: e49000. doi:10.7554/eLife.49000. ISSN 2050-084X. PMC 6994222. PMID 31755863.
- ^ Hughes, Nathan; Faulkner, Christine; Morris, Richard J.; Tomkins, Melissa (2021). "Intercellular communication as a series of narrow escape problems". IEEE Transactions on Molecular, Biological and Multi-Scale Communications. 7 (2): 89–93. doi:10.1109/TMBMC.2021.3083719. ISSN 2332-7804.
- ^ a b c d e Sager, Ross (September 26, 2014). "Plasmodesmata in integrated cell signalling: insights from development and environmental signals and stresses". Journal of Experimental Botany. 65 (22): 6337–58. doi:10.1093/jxb/eru365. PMC 4303807. PMID 25262225.
- ^ Storme, Nico (April 21, 2014). "Callose homeostasis at plasmodesmata: molecular regulators and developmental relevance". Frontiers in Plant Science. 5: 138. doi:10.3389/fpls.2014.00138. PMC 4001042. PMID 24795733.

