프로테아과
Proteaceae| 프로테아과 시간 범위: | |
|---|---|
 | |
| 프로테아신아로이드의 개화 | |
| 과학적 분류 | |
| 왕국: | 플랜태 |
| 클래드: | 기관식물 |
| 클래드: | 안지오스파름속 |
| 클래드: | 에우디코츠 |
| 순서: | 프로테알레스 |
| 패밀리: | 프로테아과 쥐.[2] |
| 일반 | |
| 약 80, 텍스트 참조 | |
프로테아과는 남반구에 주로 분포하는 꽃식물 과를 형성합니다.이 과는 83개 속에 약 [3]1,660종으로 알려져 있습니다.호주와 남아프리카 공화국은 다양성이 가장 많이 집중되어 있습니다.플라타나과(평면나무), 넬룸본과(신성한 연꽃), 그리고 최근의 APG IV 체계에서, 그들은 프로테아목을 구성합니다.잘 알려진 '프로테아과' 속에는 프로테아, 뱅크시아, 엠보스리움, 그레빌라, 하카, 마카다미아가 포함됩니다.뉴사우스웨일스 와라타(Telopea speciosisima), 킹 프로테아(Protea cynaroides), 그리고 다양한 종의 뱅크시아, 그레빌라, 그리고 류카덴드론과 같은 종들이 인기 있는 자른 꽃들입니다.마카다미아 인트그리폴리아의 견과류는 상업적으로 널리 재배되고 소비되며, 게부이나 아벨라나의 작은 규모의 견과류도 마찬가지입니다.
어원
프로테아과(Proteaceae)라는 이름은 1767년 여러 형태 [4][5]사이에서 변화할 수 있었던 그리스 신 프로테우스(Proteus)의 이름에서 유래한 프로테아(Protea)라는 이름에서 유래한, 1789년 앙투안 로랑 드 쥐시외(Antoine Laurent de Jussieu)가 가족을 위해 만든 프로테아(Proteae)라는 이름에서 로버트 브라운(Roberto Brown)에 의해 수정되었습니다.꽃과 잎의 놀라운 다양성과 [citation needed]다양성으로 유명하기 때문에 이것은 적절한 이미지입니다.
묘사

프로테아과의 속은 매우 다양하며, 특히 뱅크시아는 [6]식물의 적응 방사선의 놀라운 예를 제공합니다.이러한 변동성으로 인해 개별 세대를 쉽게 식별할 수 있지만, 제품군에 대한 간단한 진단 식별 키를 제공하는 것이 불가능합니다.
- 프로테아과는 엎드린 관목부터 40m 높이의 높은 숲 나무까지 다양하며, 허브인 일부 스털링기아 종을 제외하고 보통 중간 높이 또는 낮은 관목 또는 여러해살이 관목입니다.어떤 종들은 적합하게 낙엽성(Embothrium coccineum)이고, 드물게는 점액성(caulescentum)이며, 깃의 콜라인 부분이 종종 두꺼워집니다(Lignotuber).때로는 선모이고, 거의 없는 세 개의 세포가 있는 치조에서, 정첨 세포는 일반적으로 길쭉하고, 급성이며, 때로는 동등하거나 불균일하게 두 개입니다.
- 잎은 거의 향기롭지 않고, 대개 교대하며, 나선형으로, 드물게 반대되거나, 수직으로, 부식성이며, 드물게 육질이거나 가시가 있으며, 단순하거나 복합적인(비정형, 무정형 또는 드물게 구순형 또는 극치형)이며, (3-) 피노타티스트(고사리와 유사한 양상을 보임)의 전체 가장자리, 드물게 이분법적으로 분할되며, 종종 원격으로 치아가 있습니다.잎, 크린산염 또는 톱니 모양의, 앉거나, 또는 줄기 모양의, 바닥이 자주 부어오르지만 거의 피복되지 않은 (때로는 시나페아에서) 잎자루; 깃털 모양의 때로는 손바닥 모양 또는 평행한 정맥, 브로치드롬 모양 또는 하나의 돌출된 날개로 축소됨, 일반적으로 관자놀이; 잎자루; 이소필리는 다양한 성장 기간 동안 종종 발생합니다.뒤쪽 날개, 등쪽 날개, 등쪽 날개 또는 중심 날개, 보통 투명한 이배아세포, 희귀한 분비물 구멍을 가진 중엽 조직.Braky-paracy 스토마(Bellendena의 후핵구).
두 가지 종류의 반지름을 가진 식물 줄기, 넓고 다중 톱니 모양 또는 좁고 비 톱니 모양의 잎이 세 개인 삼엽절(흔히 하나의 흔적이 있는 일라쿠나는 거의 없음), 스크레리드가 자주 발생합니다; 렌티셀이 자주 수평으로 확대된 나무껍질, 코르크 캄비움이 있으며, 보통 표면적입니다.옆쪽 뿌리와 짧은 뿌리, 종종 매우 촘촘한 뿌리 털을 가진 다발(프로테오이드 뿌리)로 묶이고, 드물게는 근경련과 함께 있습니다.
- 식물들은 보통 암수동체이고, 더 드물게는 암수동체, 암수동체 또는 안드로몬체입니다.
- 꽃은 매우 다양하고, 단순하거나 복합적이며, 겨드랑이 또는 말단, 단독 또는 쌍으로, 드물게 말단 꽃, 라세미형, 패니큘레이트 또는 응결형, 보통 꽃차례가 있고, 때로는 잎 또는 사각형으로 전환되어 원추형의 한 종류를 형성하거나, 밝은 색으로 인크루시브 또는 유사안티움을 형성하며, 족자와어떤 경우에는 밀집된 꽃들이 초화서를 형성합니다; 매우 드물게는 꽃들이 나뭇가지 끝 근처에 단독이고 겨드랑이에 있습니다; 줄로 튜브를 가진 종에서는 꽃들이 때때로 이것들로부터 자라고 토양을 통과합니다.
- 꽃들은 보통 완벽하고, 활동형 또는 지모형이며, 저형성이고, 종종 크고 화려합니다.평평하거나 비스듬하며, 때로는 처녀공을 형성합니다.저생식 디스크가 있고 추가적인 종적 또는 부재.(3-)4(-8)테팔의 둘레(때로는 이량체 및 이량체로 해석됨), 1(-2)밸브 소용돌이에서 때때로 연장되고, 다른 방식으로 자유롭게 또는 융합됨(모든 융합 또는 심지어 하나의 자유 및 세 개의 기본이 완전히 융합됨), 심지어 튜브 또는 담즙 구조를 형성하는 미세하게 디지테이트 유두에 의해 묵인됨ure, zygomorphic, 때때로 다양한 방법으로 옆으로 열립니다.하플로세몬과 로키움, 대개는 너무 거대하고, (3-)4(-5) 수술과 반대이며, 모두 비옥하거나 일부는 스타미노데로 전환되며, 일반적으로 필라멘트이며, 테팔에 부분적으로 또는 완전히 융합된 필라멘트이며, 거의 자유롭지 않고, 염기성으로 고정된 다른 테트라세논산염, 다이테러스, 테트라스포란산염기, 때로는 단일원형 및 이중포라토세린산염기에 대한(희), 설명할 수 있습니다.세로로 찢어진 눈물을 따라 분리된, 보통 피클을 가진, 그리고 연결되어 있습니다.음순선(0-)1-4, 사각형 또는 길쭉한, 육질, 자유 또는 융합된, 용기 위에 월선 또는 고리 모양의 신관을 형성합니다.1(-2) 무균성 카펠, 무균성 또는 규정(약간 길쭉한 여성 홀 포함), 때로는 완전히 닫히지 않고, 보통 개발된 스타일로, 작은 스티그마 또는 말단 또는 하위 디스크의 형태로, 심지어 측면 및 사선으로, 종종 움푹 패인, 유두, 습윤 또는 건조한, 카펠당 1-100개 이상의 배주, 무균성, 무균성, 무균성, 무균성,반원형, 양성 또는 정형, 대부분 반원형, 무는 ggmic, crassinucellate, 혈관 다발의 고리가 있는 chalaza, funiculus는 때때로 없고 배주는 다양한 기질을 가진 태반, 가장자리 태반 또는 정반에 융합됩니다.
- 부패하거나 쇠퇴하지 않는 과일, 포도상구 또는 분자, 난포, 드루페(연선화된 내생포) 또는 거짓 드루팔(연선화된 내부 중족포), 때로는 난소 및 고환의 벽에 융합되어 종종 착색되고 장티나게 됩니다. 동일한 꽃에서 나온 과일은 때때로 융합되어 신카프를 형성합니다.
- Bellendina에 존재하는 1-많은 씨앗, 때로는 날개가 있는 평평한 것부터 둥근 것까지, 내배아가 없는 평평한 것까지, 드물게 존재하는, 잘 분화된 배아, 곧고 쌍떡잎 모양이지만, 종종 3개 이상의 큰 자엽을 가지고 있는, 종종 청각형인, 옥살산칼슘의 결정을 포함하는 특이한 층을 가진, 엔도테스타.
- 모나드의 꽃가루는 극점에서 삼각형, (2-)3(-8)-비공형, 일반적으로 등극성과 삼극성, 엠보트리움과 뱅크시에아 부족의 쌍극성, 보프레아의 콜포이드레이트, 아울락스와 프랑클란디아의 구형 또는 퍼수니아의 일부 종의 강력한 등극성;[clarification needed] 전자의 네 가지 법칙을 따릅니다.
- 염색체 수: n=5, 7, 10–14, 26, 28; 크기는 종에 따라 매우 작은(평균 1,0 μm)부터 매우 큰(평균 14,4 μm)까지 다양합니다; x=7, 12.
꽃들

일반적으로 말해서, 프로테아과의 진단적 특징은 복합 꽃머리 또는 더 정확하게는 꽃차례입니다.많은 속에서, 가장 명백한 특징은 작고 종종 매우 화려한 꽃들로 구성된 많은 작은 꽃들이 밀집된 머리 또는 스파이크입니다.그러나 이 특징조차도 모든 프로테아과에서 발생하는 것은 아닙니다; 예를 들어, 아데난토스 종들은 단 하나의 꽃을 가지고 있습니다.대부분의 프로테아과 종에서 수분 메커니즘은 매우 전문적입니다.일반적으로 꽃가루를 꽃가루 [7]매개자에게 전달하는 스타일 끝의 영역인 "꽃가루-발표자"를 사용합니다.
프로테아과의 꽃 부분은 네 개로 이루어지지만, 네 개의 테팔은 끝이 닫힌 컵이 있는 길고 좁은 관으로 융합되고, 네 개의 수술의 필라멘트는 테팔에 융합되어 다른 것들이 컵 안에 포함됩니다.암술은 처음에는 주변 튜브의 안쪽을 따라 지나가기 때문에, 낙인 또한 컵 안에 싸여 있습니다.꽃이 자라면서 암술은 빠르게 자랍니다.낙인이 갇혔기 때문에, 스타일이 길어지기 위해서는 구부러져야 하고, 결국 그것은 하나의 솔기를 따라 끝을 갈라놓습니다.그 스타일은 꿀이 생산되기 시작할 때까지 계속해서 성장합니다.이때, 둘레는 그 구성요소의 테팔로 갈라지고, 컵은 갈라지며, 암술은 어느 정도 직립하여 스프링으로 방출됩니다.
생태학

프로테아과의 많은 부분은 특수한 프로테오이드 뿌리를 가지고 있고, 가로 뿌리와 머리카락 덩어리가 방사형 흡수 표면을 형성하며, 계절적 성장기에 잎더미 층에서 생산되고, 성장기가 끝날 때 보통 오그라듭니다.그것들은 가난하고 인이 부족한 토양에서의 성장에 대한 적응으로, 이전에는 사용할 수 없었던 인을 동원하는 카르복실산염을 배출함으로써 식물이 부족한 물과 영양분에 대한 접근을 크게 증가시킵니다.그들은 또한 뿌리의 흡수 표면을 증가시키지만, 그것은 또한 그들 자신의 뿌리 [8]클러스터에 대한 영양소 경쟁을 증가시키기 때문에 이것은 작은 특징입니다.그러나, 이러한 적응은 그들을 피토포토라 시나모미 워터 곰팡이로 인한 다이백에 매우 취약하게 만들고, 일반적으로 수정을 견딜 수 없게 만듭니다.이러한 특수화된 프로테오이드 뿌리 때문에, 프로테아과는 동맥성 균근균과 공생체를 형성하지 않는 몇 안 되는 꽃식물 과 중 하나입니다.그들은 인산의 이동과 흡수를 돕기 위해 2-3일마다 많은 양의 유기산(시트르산과 말산)을 배출합니다.많은 종들은 그들의 서식지를 휩쓰는 화재에서 살아남기 위한 전략을 가지고 있다는 것을 의미하는 불에 적응합니다.어떤 것은 뿌리가 굵고, 땅에 묻어서 불이 나면 새 줄기가 돋아나는 뿌리줄기를 가지고 있는 것도 있고, 어떤 것은 다 자란 식물이 불에 죽지만, 연기에 자극을 받아 씨앗을 흩뿌려 뿌리를 내리고 자라게 하는 것도 있습니다.그 열기는 이전에는 성장을 자극했다고 생각되었지만, 지금은 연기 속의 화학물질이 그것을 유발하는 것으로 나타났습니다.
4개의 암수성 속(올락스, 딜로베이아, 헬리콥시스, 류카덴드론)과 11개의 암수성 속 및 일부 다른 속들은 비밀스럽게 암수성을 가진 종들을 가지고 있습니다: 2개의 종들은 무균 상태이고 식물적으로만 번식합니다(로마티아 태즈메이카, 하카 풀비니페라).이 종들은 자동 호환성과 자동 호환성 사이에서 중간 상황에 따라 다양합니다. 이러한 상황은 때때로 동일한 종에서 발생합니다.그 꽃들은 보통 원형입니다.합성하기 직전에, 다른 것들은 꽃가루를 방출하고, 많은 경우에 특히 자신의 꽃가루의 축적을 위해 확장된 살찐 영역을 가지고 있습니다.꿀을 먹는 사람들은 다른 사람들과 직접 접촉할 가능성은 낮지만 낙인과 접촉하는 것을 피할 수 없습니다. 따라서 낙인은 꽃가루 전달자 역할을 하여 꿀을 먹는 사람들이 꽃가루 매개자 역할을 하도록 보장합니다.이 수분 전략의 단점은 자가 수정의 가능성이 크게 증가한다는 것입니다. 많은 프로테아과는 이를 원시화, 자가 비호환성 또는 자가 씨앗의 우선적인 낙태와 같은 전략으로 반박합니다.꽃가루를 전달하는 시스템은 보통 꽃가루 매개자의 다양화에 따라 매우 다양합니다.수분은 벌, 딱정벌레, 파리, 나방, 새(꿀벌새, 태양새, 설탕새, 벌새)와 포유류( 설치류, 작은 유대류, 코끼리땃쥐, 박쥐)에 의해 수행됩니다.후자의 두 가지 평균은 진화적으로 서로 다른 독립적인 사건에서 곤충학에서 파생되었습니다.일부 종의 분산은 그들의 화생적 행동과 관련된 세로티니의 기이한 현상을 보여줍니다: 이 나무들은 바깥 층이나 보호 구조(브랙트)가 매우 정렬되고 불에 잘 타지 않는 가지에 과일을 축적합니다.열매는 불에 타서 땅에 재를 바르고 경쟁자가 없을 때에만 씨를 뿌립니다.많은 종들은 개미에 의해 분산된 엘라이오좀을 가진 씨앗을 가지고 있습니다; 날개를 가진 씨앗이나 엉겅퀴가 있는 씨앗은 에모코리를 나타내는 반면, 드럼과 다른 살찐 과일들은 포유동물과 새들이 그것들을 섭취할 때 엔도조코리를 보입니다.몇몇 아프리카와 호주 설치류들은 그들을 먹이로 먹기 위해 이 식물들의 열매와 씨앗을 둥지에 축적하는 것으로 알려져 있지만, 일부는 발아에 성공합니다.
분배
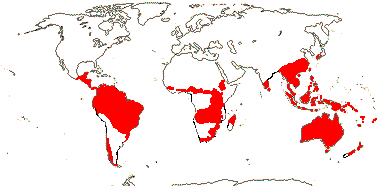
프로테아과(Proteaceae)는 주로 남반구에 분포하는 과의 하나로, 오스트레일리아와 남아프리카에 분포합니다.중앙 아프리카, 남아메리카와 중앙 아메리카, 인도, 동아시아와 동남아시아,[8] 오세아니아에서도 발생합니다.비록 화석 꽃가루 증거가 이전에 [9]더 많은 종이 있었다는 것을 암시하지만, 뉴질랜드에서 오직 두 종만이 알려져 있습니다.
남극을 제외한 사실상 모든 육지에서 택시가 발생하는 것은 고대 초대륙 곤드와나의 잔재로 여겨지는 곤드와나 가족의 좋은 예입니다.이 과와 아과는 곤드와나가 분열되기 훨씬 전에 다양화된 것으로 생각되며, 이는 이들 모두가 9천만 년이 훨씬 넘었음을 암시합니다.이것에 대한 증거는 뉴질랜드 남섬의 백악기 석탄 퇴적물에서 발견된 풍부한 단백질 꽃가루를 포함합니다.그것은 [10]해양 간격을 가로질러 분산되기보다는 주로 대륙 이동에 의해 현재의 분포를 달성한 것으로 생각됩니다.
식물화학

이 광범위한 과에 존재하는 화학 물질에 대한 결정적인 연구는 수행되지 않았습니다.프로테아속과 포레아속은 자일로스를 과즙의 주당으로 사용하고 폴리갈락톨 농도가 높은 반면, 수크로스는 그레빌라에 존재하는 주당이기 때문에 특이합니다.프로안토시아니딘(델피니딘 및 시아니딘), 플라보놀(켐페롤, 케르세틴 및 미리세틴) 및 아르부틴과 마찬가지로 티로신에서 파생된 시안성 글리코사이드가 종종 존재합니다.알칼로이드는 보통 없습니다.이리도이드와 엘라그산 또한 없습니다.사포닌과 사포게닌은 다른 종에 존재하거나 존재하지 않을 수 있습니다.많은 종들이 알루미늄을 축적합니다.

사용 및 재배

많은 전통적인 문화는 동물의 가죽을 치료하기 위한 영양, 약, 염료, 장작 그리고 건설을 위한 나무로 프로테아과를 사용했습니다.호주 원주민들은 퍼수니아 열매를 먹고, 게부이나와 마카다미아를 포함한 다른 속의 종들의 씨앗은 원주민들의 식단의 일부를 형성하지만 전 세계적으로 판매되기도 합니다.헬리키아 종의 부드러운 새싹은 자바에서 사용되고, 많은 종의 꽃에서 나오는 꽃꿀은 호주에서 마십니다.전통적인 약은 피부 상태의 국소적인 적용으로 사용되거나 두통, 기침, 이질, 설사, 소화불량, 위궤양 및 신장 질환을 치료하기 위한 강장제, 진통제, 그리고 갈락토겐으로 사용되는 많은 종의 뿌리, 나무껍질, 잎 또는 꽃의 주입으로부터 얻을 수 있습니다.이 과의 나무에서 나오는 나무는 건축과 장식과 같은 내부 용도로 널리 사용됩니다. 프로테아, 류카덴드론, 그레빌라의 나무는 특히 인기가 있습니다.많은 종들이 정원 가꾸기에 사용되는데, 특히 뱅크시아, 엠보트리움, 그레빌라, 텔로페아속이 그렇습니다.이러한 사용은 외래종의 침입을 초래했습니다; 예를 들어 포르투갈의 하카 버드나무와 실키 하카를 포함합니다.
마카다미아의 두 종은 식용 견과류를 위해 상업적으로 재배됩니다.게부이나 아벨라나(칠레 헤이즐)는 칠레와 뉴질랜드에서도 식용 견과류로 재배되며, 보습 특성과 자외선 차단제의 성분으로 제약 산업에서도 사용됩니다.그것은 [citation needed]견과류를 생산하는 나무과 중에서 가장 추위에 강합니다.그것은 또한 열대의 모습과 더 시원한 기후에서 자라는 능력을 위해 영국 제도와 미국의 태평양 연안에 심어져 있습니다.
많은 프로테아과 종들은 방벽 식물로서 그리고 두드러지고 독특한 꽃과 잎을 위해 보육 산업에 의해 재배됩니다.일부 종은 절단된 꽃 산업, 특히 일부 뱅크시아와 프로테아 종에 중요합니다.마카다미아속의 두 종은 식용 견과류를 위해 상업적으로 재배됩니다.
탑(미메테스), 아울락스, 홍조 신부(세루리아)와 같은 다른 것들뿐만 아니라 슈가부시(프로테아), 핀쿠션(레우카덴드론)은 케이프 꽃 왕국의 일부를 이루는 핀보스의 세 가지 주요 식물 그룹 중 하나입니다.크기로는 가장 작지만 풍부한 식물 왕국이자 단일 국가 내에 포함된 유일한 왕국입니다.핀보스의 다른 주요 식물군은 에리쿠스과와 레스티온과입니다.남아프리카의 프로테아는 다양한 형태와 특이한 꽃들로 인해 널리 재배되고 있습니다.그들은 아름다움과 새들과 유용한 곤충들을 유인하기 위한 야생 정원에서의 유용성으로 남아프리카에서 인기가 있습니다.
장식품으로 가장 가치 있는 종은 온대 기후의 풍경을 열대처럼 보이게 하기 때문에 남위도에서 자라는 나무입니다; 로마티아 페루기네아 (푸인케), 로마티아 히르수타 (라달)는 서유럽과 미국 서부에 도입되었습니다.Mbothrium coccineum (칠레의 불나무 또는 노트로)는 검붉은 꽃으로 인해 영국 제도에서 높은 평가를 받고 있으며 북위 62°에 있는 페로 제도만큼 북쪽에서도 찾을 수 있습니다.
온대와 지중해성 기후에서 자라는 뱅크시아 중 대다수는 관목이고, 키 때문에 가치가 있는 나무는 소수에 불과합니다.가장 높은 종들 중에는 다음과 같은 것들이 있습니다: B. integripolia와 B. integripolia subsp. monticola는 아종을 형성하는 식물들이 뱅크시아에서 가장 키가 크고 다른 뱅크시아, B. seminuda, B. littoralis, B. serrata보다 서리에 더 강하기 때문에 주목할 만합니다.큰 관목: B. grandis, B. prionotes, B. marinata, B. coccinea, B. speciosa; 이 모든 것들은 크기 때문에 공원과 정원, 심지어 길가에 심어져 있습니다.이 속의 나머지 종들, 약 170종은 관목이지만, 그들 중 일부는 꽃 때문에 가치가 있습니다.
더 작지만 세계의 일부 지역에서 재배되는 또 다른 종은 호주 뉴사우스웨일스의 산에서 온 텔로페아 스페시오시시마입니다.
일부 온대 기후 종들은 매력적인 외모 때문에 호주에서 더 현지적으로 재배됩니다.소나무 잎 지붕(Persoonia pinifolia)은 선명한 노란색 꽃과 포도와 같은 과일로 가치가 있습니다.Adenanthos sericeus (양털 덤불)은 매력적인 부드러운 잎과 작은 빨간색 또는 오렌지색 꽃을 위해 심어져 있습니다.힉스 비치아 피나티폴리아(쇠고기 견과류, 빨간 바우플 견과류)는 일반적으로 잎과 식용 견과류를 위해 심습니다.
기생충

프로테아과는 특히 특정 기생충, 특히 지중해성 기후에서 자라는 식물에서 심각한 뿌리 썩음을 일으키는 옴균 피토포라 계피노미에 취약합니다.푸사리움 옥시스포룸은 뿌리에 푸사리오시스라고 불리는 질병을 유발하여 노랗게 변하고 시들게 하며, 삼림 식물에 심각한 생태학적 손상과 상업적 관심이 있는 식물의 경제적 손실을 초래합니다.다른 일반적인 감염은 보트리오스페리아, 리조코니아, 아르밀리아, 보트리스, 칼로넥트리아와 다른 곰팡이들에 의해 발생합니다.
보존상태
국제 자연 보전 연맹(IUCN)은[11] 47종의 프로테아과가 위협을 받고 있으며, 그 중 한 종인 1935년 누벨칼레도니에서 온 스테노카르푸스 덤베엔시스 기욤이 멸종된 것으로 생각됩니다.이 과의 종들은 특히 그들의 서식지의 파괴 또는 파편화, 화재, 기생충 질병, 도입된 식물과의 경쟁, 토양 퇴화 및 인간과 그들의 가축에 의해 유발되는 다른 피해에 취약합니다.그 종은 또한 기후 변화의 영향을 받습니다.
화석

프로테아과는 진단적 특징을 보이지 않는 유해를 식별하는 데 내재된 어려움에도 불구하고 풍부한 화석 기록을 가지고 있습니다.식별은 일반적으로 brachy-paracy 기공과 특이한 트리홈 염기 또는 다른 경우에는 꽃가루 사선의 [citation needed]특이한 구조의 조합을 사용하여 이루어집니다.자일로카리온은 현존하는 [12]에도테아속과 과일의 유사성으로 인해 프로테아과의 일원으로 확인되었습니다.이 과에 속하는 화석은 곤드와나 초대륙을 형성한 대부분의 지역에서 발견되었습니다.호주 동남부의 백악기 후기 (샹파뉴-마스트리흐트)로 거슬러 올라가는 이 과에 속하는 다양한 꽃가루와 북아프리카와 페루의 백악기 중기 (세노마니아-투로니안)의 꽃가루는 트리오리테스 아프리카넨시스로 묘사됩니다.최초의 매크로 화석은 2천만 년 후에 남아메리카의 팔레오세와 호주의 북동쪽에서 나타납니다.뉴질랜드와 태즈메이니아와 같은 일부 지역의 화석 기록은 현재 존재하는 것보다 더 큰 생물 다양성을 보여주는데, 이는 많은 분류군의 분포가 시간의 흐름에 따라 급격하게 변화했고 신생대 동안 높은 수준의 멸종을 포함하여 전반적인 감소를 겪었다는 사실을 뒷받침한다.[필요 이상]
분류학

프랑스의 식물학자 Antoine Laurent de Jussieu에 의해 처음 기술된, 프로테아과는 약 80개 속으로 이루어진 꽤 큰 과이지만, 2000종 미만입니다.사실상 모든 분류학자들에게 인정받고 있습니다.고전적인 린네 분류 체계 하에서 확고하게 확립되어 있으며, 분지학 기반의 APG와 APG II 분류 체계에서도 인정받고 있습니다.그것은 프로테알레스의 순서로 배치되는데, 그 배치 자체가 다양했습니다.
프로테아과에 속하는 속들의 분류는 1975년 로리 존슨과[13] 바바라 브릭스에 의해 그들의 영향력 있는 논문 "프로테아과에 대하여: 남부 [14]과의 진화와 분류"에서 이루어졌고, 피터 H의 분자 연구로 대체되었습니다.2006년 웨스턴과 나이절 바커.프로테아과는 현재 5개의 아과로 나뉩니다.Bellendenoideae, Persoonioideae, Symphionematoideae, Proteoideae 및 Grevilleoideae.[14]2008년에 Mast와 동료들은 마카다미아와 마카다미아 부족의 관련 속들을 업데이트했습니다.게다가, 오리아테스 메가카르푸스는 마카다미에아 부족 대신에 오리아테스속이나 루팔레아 부족에 속하지 않는 것으로 밝혀졌고, 따라서 새로운 종 이름인 노토라이트 메가카르푸스가 [15]붙여졌습니다.Weston과 Barker(2006)에 따르면, Mast et al.의 속에 대한 업데이트와 함께 전체 배열.(2008)은 다음과 같습니다.
.jpg)
- 프로테아과
레퍼런스
- ^ "Proteales". www.mobot.org. Retrieved 16 June 2023.
- ^ Angiosperm Phylogeny Group logo m moi lo (2009). "An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III". Botanical Journal of the Linnean Society. 161 (2): 105–121. doi:10.1111/j.1095-8339.2009.00996.x.
- ^ Christenhusz, M. J. M. & Byng, J. W. (2016). "The number of known plants species in the world and its annual increase". Phytotaxa. 261 (3): 201–217. doi:10.11646/phytotaxa.261.3.1.
- ^ 브라운, 로버트."쥐시외의 프로테아과에 관하여".런던 린네 학회의 거래 10:15–226, 1811.
- ^ Jussieu, Antoine Laurent de, General plantarum secundum ordines naturales disposita, juxta methodum in Horto Regio Parisiensi exaratum, anno M.DCC.LXXIV.파리, 1789. 페이지 78-80 2016년 2월 16일 웨이백 머신에서 보관.
- ^ Mast, A. R. & Givnish, T. J. (2002). "Historical Biogeography and the Origin of Stomatal Distributions in Banksia & Dryandra (Proteaceae) Based on Their cpDNA Phylogeny". American Journal of Botany. 89 (8): 1311–1323. doi:10.3732/ajb.89.8.1311. PMID 21665734. S2CID 2835600.
- ^ Watson, L. & Dallwitz, M. J. (1992). "Proteaceae". The Families of Flowering Plants: Descriptions, Illustrations, Identification, Information retrieval. Retrieved 26 June 2006.
- ^ a b Orchard, Anthony E. (ed.). "Proteaceae". Flora of Australia, Volume 16: Elaeagnaceae, Proteaceae 1. Melbourne: Australian Biological Resources Study / CSIRO Publishing. Archived from the original on 31 October 2006. Retrieved 26 June 2006.
{{cite book}}:author=일반 이름 있음(도움말) - ^ Pole M (1998). "The Proteaceae record in New Zealand". Australian Systematic Botany. 11 (4): 343–372. doi:10.1071/SB97019.
- ^ Weston, P. H. & Crisp, M. D. (1996). "Trans-Pacific biogeographic patterns in the Proteaceae". In Keast, A. & Miller, S. E. (eds.). The origin and evolution of Pacific Island Biotas, New Guinea to eastern Polynesia: Patterns and processes. Amsterdam: SPB Academic Publishing. pp. 215–232. ISBN 978-90-5103-136-2.
- ^ IUCN 2006.2006 IUCN 적색 멸종위기종 목록.<www.iucnredlist.org >.2007년 2월 22일에 다운로드됨
- ^ Rozefelds, Andrew C.; Dettmann, Mary E.; Clifford, H. Trevor. "Xylocaron lockii F.Muell. (Proteaceae) fruits from the Cenozoic of south eastern Australia". Kanunnah. 1: 91–102.
- ^ L. A. S. Johnson and Briggs, B. G. (1975). "On the Proteaceae: the evolution and classification of a southern family". Journal of the Linnean Society of London. Botany. 70 (2): 83–182. doi:10.1111/j.1095-8339.1975.tb01644.x.
- ^ a b Weston, Peter H.; Barker, Nigel P. (2006). "A new suprageneric classification of the Proteaceae, with an annotated checklist of genera" (PDF). Telopea. 11 (3): 314–44. doi:10.7751/telopea20065733. Retrieved 30 March 2010.
- ^ Mast, Austin R.; Willis, Crystal L.; Jones, Eric H.; Downs, Katherine M.; Weston, Peter H. (July 2008). "A smaller Macadamia from a more vagile tribe: inference of phylogenetic relationships, divergence times, and diaspore evolution in Macadamia and relatives (tribe Macadamieae; Proteaceae)". American Journal of Botany. 95 (7): 843–870. doi:10.3732/ajb.0700006. ISSN 1537-2197. PMID 21632410. Retrieved 4 April 2013.
- Weston, P.H. (2007). "Proteaceae". Kubitzki, K. (Editor). The Families and Genera of Vascular Plants. IX. Flowering Plants - Eudicots. Springer-Verlag: Berlín. ISBN 978-3-540-32214-6.
- Hoot, S.B. & Douglas, A.W. (1998). "Phylogeny of the Proteaceae based on atpB and atpB-rbcL intergenic spacer region sequences". Australian Systematic Botany. 11 (4): 301–320. doi:10.1071/sb98027.
- Ramsey, H.P.. (1963). "Chromosome numbers in the proteaceae". Australian Journal of Botany. 11 (1): 1–20. doi:10.1071/BT9630001.
- Watson, L. & Dallwitz, M.J. (1992). "The families of flowering plants: descriptions, illustrations, identification, and information retrieval. Version: 29th July 2006". Retrieved 31 January 2007.
- 브라운, R. 쥐시외의 프로테아과에 관하여 린네 학회 10:15-216.


{kind=link}