오크나과
Ochnaceae| 오크나과 | |
|---|---|
 | |
| 브라질 남부산 소비게시아 에렉타 | |
| 과학적 분류 | |
| 왕국: | 플랜태 |
| Clade: | 기관지 식물 |
| Clade: | 혈관배양액 |
| Clade: | 유디코트 |
| Clade: | 로지드 |
| 주문: | 말피히알레스 |
| 패밀리: | 오크나과 DC.[1] |
| 속 | |
| 텍스트 참조 | |
오치나과는 말피히알레스목의 [2]꽃이 피는 식물의 과이다.APG III의 현화식물 분류 체계에서 오크나과는 대략 550여 [3]종을 포함하도록 광범위하게 정의되며, 일부 분류학자들이 별도의 과인 메두사기나과와 [1]키이나과로 취급한 것을 포함한다.2014년에 발표된 계통발생학 연구에서는 오크나과가 넓은 [4]의미로 인정되었지만, APG III 이후에 발표된 2개의 연구는 메두사기나과와 [3][5]퀴이나과를 수용했다.APG IV(2016)는 이러한 사항을 받아들이지 않았다.
이 글에서 "Ochnaceae"는 Ochnaceae sensu lato [6]또는 Ochnoids로 알려진 과의 더 큰 범위를 언급할 것이다.이런 의미에서 이 과는 약 550종의 [7]32개 속을 포함하고 있다.
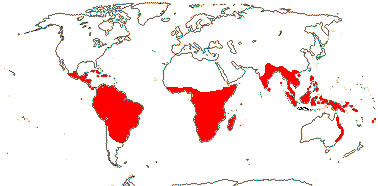
넓게 또는 좁게 정의되는 오치나과는 범열대성으로 분포하며, 이 범위 밖에서 재배되는 종도 있다.오크나과는 신생물학에서 가장 다양하며 열대 아프리카의 [4]두 번째 다양성의 중심이다.그것은 대부분 관목과 작은 나무로 이루어져 있고, 소비게시아에는 몇몇 초본 종들이 있다.많은 나무들은 나무 줄기가 직립되어 있지만 높이는 낮다.오크나과는 특이한 잎으로 유명하다.이것들은 대개 윤기가 있고, 간격이 좁고 평행한 정맥, 치아의 가장자리, 그리고 눈에 띄는 줄기가 있다.대부분의 종들은 [8]수분작용을 한다.수바게시아 부족의 8개 속에서는 꽃이 [4]핀 후 꽃 내 조직의 지속적인 성장에 의해 모양이 변한다.
오크나는 [9]관상용으로 재배되고 있습니다.Ochna Thomasiana는 아마도 가장 흔하게 심어진 식물일 것이지만, 원예 [10]문헌에서는 종종 잘못 식별된다.
세스페데시아의 잎은 때로 길이가 1m(3.3피트)에 달하며 [11]지붕에 사용된다.범열대성 잡초인 소비게시아 에렉타로 허브차를 만든다.
그 진화 과정에서, 오크나과는 조상이나 원시적인 것으로 여겨지는 특성 상태로 "복귀"한 특이한 존재였다.예를 들어, 꽃대칭 아과에서는 두 번 꽃대칭이 나타난다.또, 오치노이데아과와 키이노이데아과의 2개 군락은, 아포카르피에 매우 가까운 파생 상태를 가지고 있다.카르펠(아포카르피)의 완전한 분리는 혈관조피([12]혈관조피)
오크나과의 화석은 미시시피의 [13]초기 에오세부터 알려져 있다.그 가족의 나이는 대략 1억 [14]년으로 추정된다.
Ochnaceae에는 [15]매우 많은 속명이 발표되어 있다.오크나과의 분류학적 개정에서는 2014년 메두사기나과 1개, 퀴나과 4개, [3]오크나과 27개 등 3개 과로 분류되었다.같은 해 33번째 속인 네키아가 [4]또 다른 속인 소비게시아의 단생체를 보존하기 위해 다시 만들어졌다.
오크나과에서 가장 큰 속은 다음과 같다.오우라테아(200종), 오흐나(85종), 캄필로스페르뭄(65종), 소비게시아(39종), 퀴나(34종)[3] 등이다.어떤 큰 속도 선택된 유전자의 DNA 배열의 계통학적 분석의 대상이 되지 않았다.유전자 간 스페이서(trn L-F intergenic spacer)에 기초한 퀴이노이데아과의 한 연구에서는 이 [16]아과에서 9종만 표본 추출되었다.
속
다음 33개 속은 2014년에 [4]부활한 목과와 가장 최근의 수정판 오크나과에 [3][17][18]기술된 32개 속이다.이 분류는 Schneider et alii(2014)[4]에서 인용한 것이다.
|
묘사
아래 설명은 혈관식물의 [3][17][18]과와 속(The Families and Ochnaceae)의 Medusagynaceae, Quiinaceae 및 Ochnaceae s.s.에 대한 설명에서 발췌한 것으로, 다른 출처로부터 얻은 정보도 있습니다.
대부분 관목과 작은 나무들, 그리고 적당한 크기의 나무들, 그리고 소비게시아에는 몇 가지 허브들이 있습니다.
단순한 잎으로, 크루코비엘라와 [2]키이노이데아에서 우상복생하는 것을 제외하고, 종종 어린 식물에 우상복생하거나 화합한다.날개 모양의 잎은 유티단테라의 [4]전형이다.잎은 종종 진하고 눈에 띄게 톱니 모양이다.메두사긴을 제외하고 혈전이 있어요
정맥은 종종 평행하고 촘촘한 간격의 2차 정맥과 3차 정맥을 가진 스칼라 모양(사다리 모양)이다.꽃잎이 없거나 짧으며, 때로는 [2]심낭과 유사합니다.
단성 꽃은 메두사기네와 퀴노이데아(프로시아 제외)에서 흔히 볼 수 있지만, 오치노이데아에 속하는 3개 속에만 한정된다.단성 꽃은 슈우르만시아, 슈우르만시엘라, 에우테미스에서 [4]볼 수 있다.슈우르만시엘라에서는 꽃이 항상 남녀공용이다.[3]
일부다처제에서 꽃은 형태학만을 [16]근거로 양성애자로 평가되어 왔다.암수동물로 보이는 꽃에 의해 생성된 꽃가루는, 몇몇 경우에, 그 꽃을 기능적으로 암컷으로 만들면서, 비정상인 것으로 밝혀졌습니다.
3번부터 5번까지, 종종 불평등하고, 때로는 증가하기도 합니다.
꽃잎 4, 5, 또는 드물게 3, 6, 7, 또는 8은 밑부분에서만 어긋나고 자유롭거나 흐트러지며 때로는 비석 위로 반사되기도 한다.
5에서 10까지의 비옥한 수술 혹은 많은 수술들, 드물게는 1개.필라멘트는 때로는 지속적이고 때로는 발열기 근처에서 좁혀진다.
아테르는 담자 또는 약간 뒤섞여 있으며, 보통 하나 또는 두 개의 꼭대기 또는 아첨기 구멍에 의해 탈지되며, 때로는 세로 구멍에 의해 역방향으로 흐트러집니다.메두사긴과 퀴노이데아에서는 확대된 격막이 테케를 분리한다.
스타미노데스는 종종 꽃잎을 형성하고, 때로는 비옥한 수술들을 감싸기도 합니다.
과즙이 생산되지 않았습니다.꽃은 보통 웅성웅성 수분한다.
난소우등, 메두사기네와 키이노이데아에서 종방향으로 갈비뼈가 나 있으며, 오치노이데아에서 갈라져 있지 않다.카르펠은 완전히 융합되거나 거의 분리됩니다. 메두사긴에서는 2-15 또는 최대 25입니다.정단 또는 산모 베이직 스타일.
때로는 날개가 달린 과일, 드물게 견과류나 드랍, 종종 베리와 같은 과일, 보통 살충 캡슐, 또는 보통 붉은 빛을 띠는 용기 위에 거무스름한 드립을 형성하기 위해 분리되는 난소.
씨앗은 알부민 또는 풍성하거나 날개가 있든 없든 간에.종자 외피에는 종종 크리스타크 세포층이 포함되어 있다.이것들은 스크레이드이며, 각각 약의 형태로 옥살산칼슘 결정을 포함하고 있습니다.
분류
20세기 말까지, 오치나과는 높은 확실성을 가지기 어려운 다소 이상한 과로 여겨졌다.심지어 21세기에도 스트라스버거속은 오크나과의 가장 가까운 친척으로 취급되었고, 일부는 심지어 스트라스버거속을 그 과에 [19]포함시켰다.APG III 체계에서 스트라스버거아는 Ixerba와 함께 장미목 크로스소마목의 [20]스트라스버거아과를 형성한다.
더 드물게, 디고덴드론속은 스트라스버거리아와 오크나과에 가까운 것으로 생각되었다.분자 계통학 연구는 장미목 말발레스에 디고덴드론을 포함시키는 것을 강하게 지지하고 있으며, 때로는 [21]단종족으로 취급되기도 한다.그렇지 않으면, 백합과와 [20]더 가깝다고 의심할 만한 이유가 있지만, 백합과로 분류되었다.
Ochnaceae와 마찬가지로 위에서 언급된 모든 속들은 오랫동안 불확실한 유사성의 질투적인 분류군으로 간주되었다.이 모든 것은 한 때 또는 다른 때 테아과 근처에 있는 오크나과와 함께 놓여 있었는데, 지금은 에리카레스목의 [20]기초에 속하는 과이다.
2012년, 엽록체 DNA의 분석은 클루시오이드로 [5]알려진 다섯 과의 자매로서 오치나과를 분해했다.이 결과 부트스트랩의 지원은 약했습니다.클루시오이드는 한때 4개 [22]과의 그룹으로 여겨졌지만, 2009년에[6] 클루시아과가 분할되었고 그 결과 생긴 분리과 [1][23]중 하나를 위해 칼로필라과라는 이름이 부활했다.
Clusioids와 Ochnaceae를 결합하는 형태학적 특성은 몇 가지밖에 없다.꽃잎의 미생은 종종 클루시오이드에서 어긋나며, 보통 오크나과에서 그러하다.두 그룹 모두 꽃은 보통 여러 개의 수술을 가지고 있고 난소에서는 태반이 대부분 축삭이다.배란에서, 누셀러스는 종종 얇고, 바깥쪽 외피는 보통 [24]안쪽보다 두껍다.
분류법
Ochnaceae는 3개의 아과로 분류된다.메두사기노이데아과, 키이노이데아과, 오치노이데아과.[4]
분자 계통학 연구는 메두사기노이데아과와 퀴이노이데아과를 자매 아과로 해결했지만, 이 결과는 통계적 뒷받침이 [5]약했다.두 아과 모두 꽃은 다계통이다.프루시아속을 제외한 많은 꽃 또는 모든 꽃은 단성화이다.다른 꽃들은 다른 잎을 제거한 후에도 지속되는 테카 사이에 거대한 중격(septum)을 포함하고 있다.그 스타일은 난소에서 바깥쪽으로 발산된다.난소 합성을 할 때, 난소는 세로 [25]갈비뼈로 조각된다.
메두사기노이데아과(Medusagyne opositolia)는 단일 종인 메두사기네 오포시폴리아로 구성되어 있다.세이셸의 [26]마헤 섬의 고유종이다.
키이노이데아과는 프로시아, 키이나, 투루리아, 라쿠나리아 등 4개 속에 약 48종으로 이루어져 있습니다.열대 [27]아메리카에 한정되어 있습니다.프로이시아는 다른 세 속과는 구별된다.그것의 꽃은 항상 양성애자이고, 열매는 서로 완전히 분리되어 있지 않다는 점을 제외하고는 엽상세포와 비슷한 세 개의 구조로 이루어져 있다.
오치노이데아과(Ochnoideae)는 2014년에 오치노이데아과(Ochnaceae sensu stricto)[3]로 개정되었다.그 처리에서는 27개의 속들이 기술되었다.분자 계통학 [4]연구 결과에 따라 같은 해 네키아라는 다른 속들이 부활했다.아마랄과 비트리히(2014)는 오치노이데아를 룩셈부르크에, 소비게에, 그리고 오치네의 세 종족으로 나눴다.테툴레아속은 소비게아족에 포함되었습니다.인식된 서브리브는 없습니다.
2014년에는 DNA 배열의 클래디스트 분석을 바탕으로 Ochnoideae의 2차 재분류가 발표되었다.이 논문에서 Schneider et alii는 Ochnoideae를 4개의 종족으로 나누었다.Testuleae, Luxemburgieae, Sauvagesieae 및 Ochneae.그들의 부족에 대한 제한은 테툴레아가 소비게아에서 테툴레아 부족인 테툴레아로 옮겨졌다는 점을 제외하고는 아마랄과 비트리히(2014년)와 동일했다.Sauvagesieae에 Testulea를 포함시키면 Luxemburgieae에 대한 그 부족의 측문법이 나타난다.
Testulea는 가봉의 고유종인 Testulea gabonensis라는 단일 종으로 구성되어 있습니다.잎이 브로키도롬 모양의 정맥 무늬를 가지고 있고 꽃은 4원추형이라는 점에서 오치노이데아과에서 특이하다.게다가 수술 중 하나만이 비옥하다.다른 것들은 스타미노드로 변형되어 그 길이의 최대 3µ까지 기둥으로 결합된다.
룩셈부르크 부족은 필라크라 족과 룩셈부르크 족의 두 개의 속으로 구성되어 있습니다.Philacra는 베네수엘라와 브라질 북부가 원산지입니다.룩셈부르크는 브라질 출신이다.
소비지아족은 전열대성 종으로 16개 속, 대부분 작은 것으로 이루어져 있다.지금까지 가장 큰 것은 소비게시아로, 38종이 있으며, 그 중 35종은 신트로픽에 [11]한정되어 있다.소비지아는 이질적이며, 심지어 네키아가 제거된 상태에서도 측문증일 수 있다.소비지아족의 관계는 잘 알려져 있지 않고,[4] 이러한 이유로 세분화 되어 있지 않다.
오치네 부족은 대부분의 열대 지방에서 발견되지만 아프리카와 열대 아메리카에서 가장 많이 발견됩니다.종자가 성숙하기 전에 배추를 흡수한다는 점에서 오크노이데과의 나머지 종자와 구별된다.9개 속은 3개 아강에 속합니다.Lophirinae, Elvasiinae, Ochninae.[4]
로피라아과(Lophirinae)는 단일 속인 로피라(Lophira)로 구성되어 있다.그것은 열대 아프리카에 국한된 두 가지 종을 가지고 있다.그것은 특이한 열매를 맺는데, 그 열매에서 두 개의 무덤이 크게 커지고 바람에 의한 씨앗 분배를 용이하게 하는 날개를 형성한다.
엘바시아아과(Elvasiinae)는 두 개의 속, Perisocarpa와 Elvasia로 구성되어 있으며 둘 다 미국 열대 지방에 한정되어 있다.주변 세포는 분자 계통학 연구를 위해 표본 추출된 적이 없다.
오크나과는 6개의 속으로 구성되어 있다: 캄필로스페르무움, 오우라테아, 이데르티아, 브라켄리지아, 랍도필룸, 그리고 오크나.이들 중 가장 큰 종인 Ouratea는 신대륙에 한정되어 있으며 Ochninae에 있는 모든 신대륙 종들을 포함하고 있다.Amaral과 Bittrich(2014)에 의해 정의된 대로 Ochneae의 모든 속은 단일통으로 보이지만, 한 분자 계통학 연구에서 Ouratea와 Ochna는 최대우도 [4]분석에서 약한 부트스트랩 지지만 받았다.이데르티아와 브라켄리지아는 자매속일 가능성이 높지만, 오치나과의 다른 관계는 아직 밝혀지지 않았다.
계통발생학
아래 계통수는 2014년에 [4]출판된 계통수를 개작한 것이다.약하게 지원되는 노드는 접혀 폴리토미가 형성됩니다.최대우도 부트스트랩의 지원은 75%를 넘습니다(지정된 경우는 제외).Perisocarpa와 Indosinia는 DNA를 위해 샘플링되지 않았다.계통수 위에 그들의 위치는 오직 해부학과 형태학에 기초한다.
| 중음속 키이노이데아 키보드 |
진화
오크나과의 진화는 두 개의 반동형 꽃대칭으로의 완전한 복귀와 두 개의 반동형 꽃대칭으로의 거의 완전한 복귀를 포함하고 있다는 점에서 특이하다.액티노몰피와 아포카피는 혈관배엽에서 "[28]원시적인" 성격상태로 여겨진다.2차 아포카피는 특히 희귀하며 장미과, 아포키아과, 사핀달레스,[12] 말발레스에서 가장 두드러지게 발생한다.
꽃은 메두사기네와 퀴노이데아에서는 액티노모르피이지만, 오치노이데아에서는 지고모르피가 조상 조건입니다.테스툴레아, 필라크라, 룩셈부르크에서는 꽃봉오리에 지고모형적으로 꽃이 핀다.그러나 꽃봉오리속, 고도야속, 리티단테라속, 크루코비엘라속, 세페데시아속, 플뢰리도라속, 포에실란드라속, 월라시아속 등 소비기아과의 4개 기초군에서는 꽃봉오리속에서는 꽃봉오리 중 어느 부분의 생육에 의해 형질적으로 작용하여 지고모르픽(Zygomorphic)이 된다.이러한 늦은 지고모르피는 꽃이 피는 식물에서 매우 드물다.네키아속, 슈르만시아속, 슈르만시아속, 에우테미스속, 타일러리아속, 아데나라케속, 인도시니아속, 소비게시아속 등 나머지 다섯 번째 소비게과 분류군에서는 꽃이 합성 후에도 활동하지 않고 있다.Ochneae 부족에서는 모든 종이 액티노이드 모양의 꽃을 가지고 있다.
오치나아강과 프로시아속에서는 난소(카펠)의 구성 요소가 아주 짧은 시간 동안 기저부에서 결합됩니다.그렇지 않으면, 오크나과의 난소는 완전히 녹은 카르펠로 구성된 합성암모양입니다.
메두사기네와 키이노이데아에서는 많은 꽃들이 양성애자이며, 프로시아에서는 엄밀하게 암수동체이다.오치노이데아에서 단성 꽃은 슈우르만시아, 슈우르만시엘라, [4]에우테미스로 이루어진 군락으로 한정된다.
Medusagyne와 Quiinoideae에서는 대부분의 안지오스피름과 마찬가지로 종방향의 틈에 의해 안서가 열립니다.Ochnoideae에서, Anthher dehiscence는 조상 때부터 살상력이 있으며, 세로 구멍으로 여러 번 되돌아간다.테스툴라, 필라크라, 룩셈부르크에는 꼭대기 모공에 의해 열리는 앙테르가 있다.소비게아과에서 가장 기본적인 세 가지 분류군인 Blastemanthus, Fleurydora, 그리고 5개의 카르펠과 1개의 카르펠당 많은 배란을 가진 4개의 속 분류군(Godoya, Rhttidanthera, Krukoviella, 그리고 Cespedia)도 마찬가지입니다.Poecilandra는 살상력이 있지만 자매속인 Wallacea에서는 종으로 갈라진 틈으로 열려 있다.
소비지애의 나머지 지역은 다른 다양한 지역들이 있다.슈르만시아, 슈르만시엘라, 아데나라케에서, 다른 탈수증은 살상력이 매우 높다.즉, 종방향 슬릿이 짧고 Anter의 끝단으로부터 멀리 뻗어 있지 않습니다.소비게시아의 몇몇 종에서, 앵커는 세로로 갈라지지만, 전체 안드로슘은 꽃잎 모양의 스타미노드로 싸여 꽃가루가 앵커의 꼭대기에서만 빠져나갈 수 있습니다.이것은 마치 앵더가 진짜 살생하는 것처럼 기능하기 때문에 살생 시스템으로 알려져 있다.
Ochneae 부족에서는, 종으로 갈라진 틈에 의한 다른 해빙은 Brackenridgea와 Ochna의 몇 종에 한정되어 있다.
고환은 하나의 비옥한 수술만 가지고 있는 것이 특이하다.원추공에 의해 열리게 됩니다.다른 수술들은 무균 스타미노드로 변형되어 최대 3µ 길이의 기둥으로 융화됩니다.
프로시아속, 퀴나속, 오치네족에서는 종자 발육 초기에 배추가 완전히 흡수된다.내복의 유무가 오크나과의 조상 상태인지 아닌지는 명확하지 않다.오랫동안 오치노이데아과는 이 캐릭터만을 바탕으로 두 그룹으로 나뉘었다.이러한 분류에서 배추를 포함하는 그룹은 Testulea, Philacra 및 Luxemburgia를 포함하기 때문에 Ochneae 위에 측문된다.
카펠당 배란의 수는 오크나과에서 매우 다양하다.메두사긴과 키이노이데아는 카펠당 2개의 배란을 가지고 있다.Saubagesieae의 4개 속(Godoya, Rhttidanthera, Krukoviella, Cespedesia)의 고환과 분지에서는 배란의 수는 카르펠당 100개에서 200개입니다.에우테미스를 제외한 소비게아 지방의 나머지 지역과 필라크라 및 룩셈부르크의 경우 카르펠당 배란의 수는 4개에서 50개까지이다.에우테미스는 카펠당 2개의 배란을 가지고 있다.
오치네에서 로피라는 카펠당 4개에서 50개의 배란을 가지고 있다.Elvasiinae와 Ochninae의 아종은 카르펠 1개당 배주수가 1개이다.
역사
오치나과는 1811년 [29][30]아우구스틴 피라무스 드 칸돌레가 세웠다.그 당시, 그는 이 과의 새로운 속인 엘바시아를 기술했고, 그는 오크나, 월케라, [31]곰피아 세 가지를 포함시켰다.월케라는 1789년 요한 크리스티안 다니엘 폰 슈레버에 의해 묘사되었지만, 더 이상 인정되지 않는다.모식종은 1968년 안드리아스 카니스에 의해 [32]곰피아 세라타로 기술됐지만 현재는 캄필로스페르무속으로 [citation needed]분류되고 있다.곰피아는 오랫동안 혼란의 원인이[33] 되어 왔으며, 가장 최근의 오치나과의 [3]개정판에서는 인정되지 않았다.
고도야와 소비게시아는 드 칸돌레가 오치나과를 세운 1811년에 알려져 있었지만, 그는 그것들을 다른 과에 두었다.그의 Prodromus에서 그는 고도야를 나중에 Clusiaceae로 [34]알려진 과로 분류했다.그는 라우라디아(라브라디아)를 소비게시아와 분리하여 둘 다 비올라과로 분류하였다.그는 오크나과에 카스텔라속을 추가했지만, 지금은 시마루바과에 [35]속합니다.드 칸돌레는 시마루바과가 오크나과와 밀접하게 관련되어 있다고 믿었지만, 지금은 사핀달레스목([20]Sapindales몇몇 작가들은 21세기 [19]초까지 고도야, 소비게시아 등을 소비게시아과로 분류했다.아돌프 엥글러와 같은 다른 작가들은 그것들을 오크나과에 포함시켰다.
1874년, Engler는 성숙한 [36]씨앗에 배추가 없거나 존재함에 따라 Ochnaceae를 두 그룹으로 나누었다.배추가 없는 집단은 드 칸돌레의 오크나과의 개념과 현대의 오크네에족에 해당한다.배아를 가진 그룹은 현재 측계통으로 알려져 있으며 Testuleae, Luxemburgieae, Sauvagesieae로 구성되어 있다.1876년, 브라질의 한 식물에서, Engler는 Ochnaceae,[37] 특히 가장 큰 속인 Ouratea의 많은 새로운 종을 묘사했습니다.그는 오우라티아의 85종을 묘사했고, 그 중 17종을 당시 새로운 종으로 명명했다.그는 또한 63종을 다른 속으로부터 우라테아로 옮겼다.
퀴나속과 투룰리아속은 1775년 장 베티스트 [38]오블렛에 의해 기술된 이후 알려졌으며 19세기 분류학자들에 의해 다양하게 분류되었다.자크 데니스 초이는 [39]1849년에 그들을 위해 퀴나케(Quiinaceae, 퀴나케)과를 세웠지만, 그는 식물 이름의 유효한 출판 요건을 충족하지 못했다.퀴나과라는 이름은 1888년 [30][40]Flora Brasiliensis에서 Engler에 의해 확인되었다.
메두사긴속은 1877년 존 길버트 베이커에 의해 모리셔스와 세이셸의 [41]식물군에 의해 기술되었지만 1924년이 되어서야 자신의 단일 과로 [42]분리되었다.
1893년, Ernest Friedrich Gilg는 Ochnaceae를,[43][44] 아돌프 Engler는 Quiinaceae를 다루었습니다.엥글러는 1897년 [45]DNP 초판 부록으로 메두사긴에 대한 설명을 썼다.그는 메두사긴을 "Zweifelhafte, möglicherweise zu den Guttiferae gehörige Gattung" (의심스러운, 아마도 Guttiferae에 속하는 속)이라는 제목으로 분류했다.Guttiferae는 Clusiaceae의 구식 명칭이다.
1902년 필립 반 티에헴은 현재의 오치노이데아 [46]아과인 여섯 가족을 인정했다.이들은 룩셈버기아과, 소비게시아과, 월리스과, 우테미다과, 로피라과, 오크나과였다.이들 중 3개(Wallaceae, Euthemidaceae, Lophiraceae)는 단일 유전자로 당시 반 티에헴에 의해 세워졌다.그의 Luxemburgiaceae는 오늘날 소비게이 부족의 기초 분지군을 포함했다.Van Tierghem은 1902년에 매우 많은 속들을 매우 좁은 범위로 명명했다.지금은 오치나과의 하위 분류인 오치나과에서 그는 53개의 속들을 묘사했다.가장 최근의 개정판에서는 6개의 속으로 나뉩니다.
1925년 DNP 제2판을 위해 Engler와 Gilg는 1893년에 [47][48]썼던 것과 비교하여 각각 Quiinaceae와 Ochnaceae의 처리를 확장했다.메두사기나과는 아돌프 엥글러와 한스 멜키오르가 [49]같은 부피의 DNP로 덮었다.
DNP의 두 번째 판에서 Engler는 Quiina와 Touroulia라는 두 개의 속을 Quiinaceae에 포함시켰습니다.라쿠나리아와 프루시아는 나중에 발견되었고 각각 1925년과 1948년에 이름이 붙여졌다.
같은 권에서 길그는 그의 오크누아과(현재의 오크누아과와 동일)를 인도베아, 레이트게비아, 바우사게시아, 라우라디아 등 21개의 속으로 나누었는데, 이 속들은 현재 사우바게시아의 [3]동의어로 여겨진다.현대의 8개 속(Philacra, Krukoviella, Fleurydora, Tyleria, Adenarake, Indosinia, Perisocarpa, Idertia)은 당시 식물 탐사에 의해 발견되지 않은 식물들로 구성되어 있다.Gilg는 Rhttidanthera를 Godoya의 동의어로 두었지만, Rhttidanthera는 오늘날 받아들여지고 있다.그는 곰피아의 캄필로스페르멈과 랍도필룸을 포함시켰고, 곰피아를 오라테아 아래에 동의어로 두었다.길그는 오흐나과의 개정판에서 자신의 [48]분류뿐만 아니라 판 티에헴의 분류에 대한 요약을 제공했다.반 티에헴의 3개 속(Campylospermum, Rhabdophylum, Rhabdophyla)은 [3]오늘날에도 여전히 알려져 있다.
1968년 안드리아스 카니스는 2014년 [32]개정판까지 오치나과의 후속 연구에 큰 영향을 미친 논문을 발표했다.Claude H.L. Sastre는 1970년부터 [4]2003년까지 여러 논문에서 Ochnaceae에 많은 새로운 종을 명명했다.
1991년, Ochnaceae에 [50]대한 클래디스트 분석이 발표되었다.같은 해, 4개의 엽록체 DNA 위치 및 핵 리보솜 [4]ITS에 기초한 분자 계통학 연구에서 Ochnoideae의 28번째 속인 Neckia가 부활했다.79종의 오크나과를 표본 추출하여 새로운 분류가 제시되었다.또한, 고환족은 소비게아족에서 제거되어 단형성 고환족으로 분류되었다.
레퍼런스
- ^ a b c Angiosperm Phylogeny Group (2009). "An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III" (PDF). Botanical Journal of the Linnean Society. 161 (2): 105–121. doi:10.1111/j.1095-8339.2009.00996.x. Retrieved 2013-07-06.
- ^ a b c 버논 H.헤이우드, 리처드 K.브루미트, 올레 세버그, 알라스테어 컬럼입니다세계의 꽃이 피는 식물과.파이어플라이 북스: 캐나다 온타리오.(2007).ISBN 978-1-55407-206-4.
- ^ a b c d e f g h i j k 마리아 do Carmo E.아마랄, 그리고 볼커 비트리치, 2014년.'오치나과', 253-268페이지. doi:10.1007/978-3-642-39417-1_19인치:클라우스 쿠비츠키(편집자).2014. 혈관식물 과 및 속 제11권.Springer-Verlag: 베를린, 하이델베르크, 독일.ISBN 978-3-642-39416-4(프린트).ISBN 978-3-642-39417-1(eBook).도이: 10.1007/978-3-642-39417-1
- ^ a b c d e f g h i j k l m n o p q 훌리오 5세슈나이더, 풀체리 비시엥구, 마리아 두 카르모.아마랄, 알리 타히르, 마이클 F.페이, 마르코 타인, 마크 S.M. 소세프, 게오르크 지즈카, 라스 W.채팅루2014. "5개의 DNA 영역에 기초한 범열대성 오크나과(Medusagynaceae, Ochnaceae s.str., Quiinaceae)의 계통학, 조상 상태 재구성 및 새로운 가족 분류"분자 계통학 및 진화 78:199-214.doi:10.1016/j.ympev.2014.05.018.
- ^ a b c 진샹자이, 브래드 R.Ruhfel, Hanno Schaefer, Andre M. Amorim, Manickam Sugumaran, Kenneth J. Wurdack, Peter K.엔드레스, 메릴런 L. 매튜스, 피터 F.스티븐스, 사라 매튜스, 찰스 C.데이비스, 2012년"Phylogenomics와 사후 데이터 분할은 백악기 Angiosperm 방사선 Malpighiales를 해결합니다."PNAS(미국 국립과학원회보) 109(43):17519-17524.doi:10.1073/pnas.1205818109. (아래 외부 링크 참조)
- ^ a b Kenneth J. Wurdack; Charles C. Davis (2009), "Malpighiales phylogenetics: Gaining ground on one of the most recalcitrant clades in the angiosperm tree of life", American Journal of Botany, 96 (8): 1551–1570, doi:10.3732/ajb.0800207, PMID 21628300
- ^ Christenhusz, M. J. M.; Byng, J. W. (2016). "The number of known plants species in the world and its annual increase". Phytotaxa. 261 (3): 201–217. doi:10.11646/phytotaxa.261.3.1.
- ^ Paul A. De Luca와 Mario Vallejo-Marin, 2013."무슨 소리예요?버즈 수분작용의 생태와 진화적 의의.식물생물학 16(4):429-435.doi:10.1016/j.pbi.2013.05.002.
- ^ 앤서니 헉슬리, 마크 그리피스, 마고 레비(1992년).새로운 왕립 원예학회 정원 가꾸기 사전.맥밀런 프레스, London.Stockton Press:뉴욕.ISBN 978-0-333-47494-5 (세트)
- ^ 워렌 L. 와그너, Deren R.허브스트, 그리고 Sy H. Sohmer.1999년 개정판 하와이 현화식물 매뉴얼.비숍 박물관 프레스:호롤루루
- ^ a b 데이비드 J. 매벌리2008. 메이벌리의 식물책 제3판(2008).케임브리지 대학 출판부: 영국.ISBN 978-0-521-82071-4(아래 외부 링크 참조)
- ^ a b 피터 K.엔드레스 2011년"혈관조피 꽃의 진화적 다양화"미국 식물학 저널 98(3):370-396.doi:10.3732/ajb.1000299.(아래 외부 링크를 참조하십시오).
- ^ 다니엘 데인히, 피터 윌프, 스테판 A.작은.2007. "붉은 핫트럭 정지 지역(미시시피 메리디언)의 초기 에오세 매크로플로라." Palaeontologia Electronica 10(3):17A:31 페이지.(아래 외부 링크를 참조하십시오).
- ^ 수사나 마갈론, 키디르 W. 힐루, 디트마르 크반트 2013."토지 식물 진화 일정:유전자 효과는 연령과 치환율의 완화된 시계 추정에서 화석 제약보다 이차적이다."American Journal of Botany 100:556-573.doi:10.3732/ajb.1200416. (아래 외부 링크 참조)
- ^ 플랜트 리스트:오크나과(아래 외부 링크를 참조하십시오).
- ^ a b 훌리오 5세Schneider, Ulf Swenson, Rosabelle Samuel, Tod Stuessy, Georg Zizka, 2006."Quiinaceae(Malpighiales)의 계통학: trnL-trnF 배열 데이터와 형태학의 증거"플랜트 계통학 및 진화 257(3-4):189-203.doi:10.1007/s00606-005-0386-5.
- ^ a b 클라우스 쿠비츠키, 2014년"Quiinaceae", 277-281페이지. doi:10.1007/978-3-642-39417-1_22.인 : 클라우스 쿠비츠키 (편집자)2014. 혈관식물 과 및 속 제11권.Springer-Verlag: 베를린, 하이델베르크, 독일.ISBN 978-3-642-39416-4(프린트).ISBN 978-3-642-39417-1(eBook).도이: 10.1007/978-3-642-39417-1
- ^ a b 윌리엄 C.디키슨과 클라우스 쿠비츠키, 2014년.'메두사기나과', 249-251페이지. doi:10.1007/978-3-642-39417-1_18.인 : 클라우스 쿠비츠키 (편집자)2014. 혈관식물 과 및 속 제11권.Springer-Verlag: 베를린, 하이델베르크, 독일.ISBN 978-3-642-39416-4(프린트).ISBN 978-3-642-39417-1(eBook).도이: 10.1007/978-3-642-39417-1
- ^ a b 아르멘 L.탁타잔(탁타지안).플라워링 플랜트 제2판(2009년).Springer Science + 비즈니스 미디어.ISBN 978-1-4020-9608-2.ISBN 978-1-4020-9609-9 doi:10.1007/978-1-4020-9609-9. (아래 외부 링크 참조).
- ^ a b c d 피터 F.스티븐스(2001년 이후).오크나과: Angiosperm Phylogeny 웹사이트.주소: 미주리 식물원 웹사이트.(아래 외부 링크 참조)
- ^ 클레멘스 바이어, 2003년다이고덴드론 175-177쪽인 : 클라우스 쿠비츠키 (편집자)2003. 관상식물과 속 제5권(권편집자: K. Kubitzki, C).바이엘)Springer-Verlag: 베를린, 하이델베르크, 독일.ISBN 978-3-540-42873-2(프린트) ISBN 978-3-662-07255-4(eBook)doi: 10.1007/978-3-662-07255-4.
- ^ 피터 F.스티븐스.2007. "클루시아과 – 구티페라." 48-66페이지.인 : 클라우스 쿠비츠키 (편집자)2007. 관상식물과 속 제9권(권편집자: K. Kubitzki와 C와의 콜라보레이션).바이어와 P.F.스티븐스).Springer-Verlag: 베를린, 하이델베르크, 독일.ISBN 978-3-540-32214-6() ISBN 978-3-540-32219-1() doi:10.1007/978-3-540-32219-1.
- ^ 브래드 R. 루펠, 피터 F.스티븐스, 그리고 찰스 C.데이비스, 2013년"클루시오이드 분지(Malpighiales)의 형태학적 및 분자적 계통 발생과 고대 로지드 매크로포실 고생루시아의 위치"국제식물과학저널 174(6):910–936.doi:10.1086/670668.
- ^ 피터 K.엔드레스, 찰스 C.데이비스, 그리고 메릴런 L. 매튜스.2013. "꽃식물 중 가장 큰 목 중 하나인 말피기아목의 주요 아계층의 꽃 구조 특성화 진전"식물연보 111(5): 969~985.doi:10.1093/aob/mct056. (아래 외부 링크 참조).
- ^ Merran L. Matthews, Maria do Carmo E.아마랄, 그리고 피터 K.엔드레스. 2012년."Ochnaceae s.l. (Ochnaceae, Quiinaceae, Medusagynace; Malpighiales)의 비교 꽃 구조와 계통학"Linnean Society 식물학회지 170(3):299-392.doi:10.111/j.1095-8339.2012.01299.x.
- ^ 윌리엄 C.디키슨, 1990년메두사긴과(Medusagyne)의 형태와 관계.플랜트 계통학 및 진화 171(1-4): 27-55.
- ^ 훌리오 5세슈나이더와 게오르크 지즈카입니다'퀴나과'.플로라 네오트로피카 모노그래프?
- ^ 더글러스 E.솔티스, 파멜라 S 솔티스, 피터 K엔드레스, 그리고 마크 W. 체이스, 2005년.혈관배엽의 계통발생과 진화.시나우어 어소시에이츠 주식회사:선덜랜드, 미국, ISBN 978-0-8793-817-9.
- ^ 국제식물명인덱스의 Ochnaceae. (아래 외부 링크 참조).
- ^ a b 제임스 L. 디플로2008년 이후."현존 혈관식물의 과명 및 상과명 체크리스트"주소: James L. Shellow 및 C 홈페이지.로즈 브룸.(아래 외부 링크 참조).
- ^ 아우구스틴 피라무스 드 칸돌레 1811년Nouveau bulletin des sciences par la Société philomathique de Paris 2 (40) : 208. (아래 외부 링크 참조).
- ^ a b 안드리아스 카니스, 1968년"인도-태평양 지역의 오크나과의 개정판"Blumea 16 (1):1-83.
- ^ 베르너 그로이터와 로사 랭킨 로드리게스입니다2014. "곰피아(오키아과)의 한 종류 – 다시 한 번"분류 63(5):1122-1123.doi:10.12705/635.5
- ^ 아우구스틴 피라무스 드 칸돌레, 1824년"Violaceae", 287-316페이지, Clusiaceae, 557-564페이지, "Ochnaceae", 735-738페이지.In: Prodromus Systematis Naturalis Regni Vegetabilis volume 1. (아래 외부 링크 참조)
- ^ 조슈아 W.클레이튼, 2011년408-423쪽의 '시마루바과'인 : 클라우스 쿠비츠키 (편집자)2011. 혈관식물 과 및 속 제X권.Springer-Verlag: 베를린, 하이델베르크, 독일.ISBN 978-3-642-14396-0 (프린트).ISBN 978-3-642-14397-7(전자책).도이: 10.1007/978-3-642-14397-7
- ^ H.G. 아돌프 엥글러 1874년"에베르 베그렌중 und systematische Stellung der naturlicen Familie der Ochnaceae"Nova Acta Academiae Caesaremieae Leopoldino - Carolinae Germannae Naturae Curiosum 37 (2) : 1 - 28 (아래 외부 링크 참조).
- ^ H.G. 아돌프 엥글러. 1876. 오크나과: 칼 F.P. 폰 마르티우스와 아우구스트 빌헬름 아이클러(편집자).Flora Brasiliensis 12 (2): 297-366 (아래 외부 링크 참조).
- ^ 장 침례교 C.F.오블렛.1775. 기앙 프랑수아즈 역사: · · · · · · Quiina :제1권 및 제2권 보충(19페이지), 투르울리아: 1:492, 오우라테아: 1:397 (아래 외부 링크 참조)
- ^ 자크 데니스 초이시, 1849년des Guttiferes de l'Indie, · · · · :12.
- ^ H.G. 아돌프 엥글러 1888년"Quiinaceae", 475-486페이지와 표 109-110.인: 칼 F.P. 폰 마르티우스와 아우구스트 빌헬름 아이클러(편집자)Flora Brasiliensis 12 (1) (아래 외부 링크 참조).
- ^ 존 길버트 베이커 1877년모리셔스와 세이셸의 플로라: 16. L. 리브 & Co.:런던, 영국(아래 외부 링크 참조).
- ^ H.G.아돌프 엥글러와 어니스트 F.길그 1924년Icllabus der Pfranzenfamilien : eine übersicht · · neunte und zehnt auflage : 280 . (식물과의 개요 · ··판 9 및 10, 280페이지)
- ^ 어니스트 F.길그 1893년"오키나과", 131-153페이지입력: H.G. 아돌프 엥글러와 칼 A.E. 프란틀(편집자)1895년 다이 나튀리첸 플란젠파밀린 제1판:테일 3권, 압테룽 6권 (3권, 6편)Verlag von Wilhelmn: 라이프치히, 독일.(아래 외부 링크를 참조하십시오).
- ^ H.G. 아돌프 엥글러 1893년"Quiinaceae", 165-167페이지입력: H.G. 아돌프 엥글러와 칼 A.E. 프란틀(편집자)1895년 다이 나튀리첸 플란젠파밀린 제1판:테일 3권, 압테룽 6권 (3권, 6편)Verlag von Wilhelmn: 라이프치히, 독일.(아래 외부 링크를 참조하십시오).
- ^ H.G. 아돌프 엥글러, 1897년메두사긴 250쪽H.G. 아돌프 엥글러, 1897년'구티페레', 247-250페이지H.G. 아돌프 엥글러와 칼 A.E. 프란틀 1897년Die Natürichen Pfranzenfamilien: 나흐트래게 [I] 줌 II-IV 틸 (자연식물군:볼륨 2-4에 대한 첫 번째 보충 자료). (아래 외부 링크를 참조하십시오.)
- ^ 필립 E.L. 반 티켐.1902년 '오치나세 수르'Annales des Sciences Naturelles – botanique, séries 8 16:161-416.
- ^ H.G. 아돌프 엥글러 1925년"Quiinaceae", 106~108페이지입력: H.G. 아돌프 엥글러와 칼 A.E. 프란틀(편집자)1925. Die Naturlichen Pfranzanfamilien: zweite auflage 21 Band. (자연식물군: 제2판, 제21권).Verlag von Wilhelmn: 라이프치히, 독일.
- ^ a b 어니스트 F.1925년 길그"Ochnaceae", 53-87페이지입력: H.G. 아돌프 엥글러와 칼 A.E. 프란틀(편집자)1925. Die Naturlichen Pfranzanfamilien: zweite auflage 21 Band. (자연식물군: 제2판, 제21권).Verlag von Wilhelmn: 라이프치히, 독일.
- ^ 1925년 한스 멜키오르"메두사기나과", 50-52페이지.입력: H.G. 아돌프 엥글러와 칼 A.E. 프란틀(편집자)1925. Die Naturlichen Pfranzanfamilien: zweite auflage 21 Band. (자연식물군: 제2판, 제21권).Verlag von Wilhelmn: 라이프치히, 독일.
- ^ 마리아 do Carmo E.아마랄, 1991년"Phylogenetische Systematik der Ochnaceae."Botanische Jahrbücher für Systematik, Pfranzengeschichte, und Pfranzengographie 113 (1):105-196.
외부 링크
- Xi2012 위치: 퍼브드 센트럴 위치: 국립생명공학정보센터
- 세스페데시아 그리고. 오흐나 그리고. 소비게시아 인 메이벌리의 식물책
- 엔드리스 (2011) 그리고. 육지 식물 진화 연표 위치: 기록 보관소 위치: 미국 식물학 저널
- 미시시피산 Eocene 매크로플로라(프린트) 위치: 목차. 위치: 제10권, 제3호 위치: 문제를 커버별로 정리하다 위치: 기사 참조 위치: Palaeontologia Electronica / 콘텐츠
- 오크나과 위치: 혈관 배지 위치: 둘러보다 위치: 플랜트 리스트
- 오크나과 인 탁타잔 2009
- 오치노이데아 분포도 그리고.오크노이데아과의 속 목록 앳 오치노이데아과 앳말피히알레스 앳 나무들 위치: AP웹 위치: 미주리 식물원
- 클루시오이드 군락과 고생루시아 입력: 제174권, 제6호 대상: IJPS
- 말피히알레스의 꽃 구조[데드링크] 입력: 기록 보관소 위치: 식물 연보
- 오크나과 위치: 플랜트명 위치: IPNI
- OA-OZ 그리고. Q 앳 현존 혈관식물의 초속명 체크리스트 앳 James L. Shellow 및 C 홈페이지로즈 브룸
- 곰피아 앳 분류군
- 모리셔스와 세이셸의 플로라 앳 Archive.org
- BHL(생물다양성 유산 라이브러리):
- 오크나과, 208페이지 인 제40호 앳 책 보기 앳 Nouveau bulletin des sciences par la Société philomathique de Paris, tome 2
- 라브라디아 그리고. 소비게시아 그리고. 고도야 그리고. 카스텔라 앳 책 보기 앳 Prodromus v.1(1824)
- 제37권, 제2호 오치나과 인 제37권 앳 노바악타아카데미 레오폴디노카롤리아게게르마니아나투라내추럴래 큐리오룸
- 오크나과 인 제12권 제2부 앳 플로라브라질리엔시스
- 퀴나 인 제2권 보충 인 제2권 그리고.투루리아 그리고. 우라테아 인 제1권 의 기앙 프랑수아즈 미술관
- 퀴이나과 인 제12권 제1 플로라 브라질리엔시스
- 오크나과 그리고. 퀴이나과 인 제3권 제6부 의 디 나튀리첸 플란젠파밀린
- 메두사긴 인 Nachtraege (보조) Teil 2-4 앳 디 나튀리첸 플란젠파밀린
- 수르 레 오슈나세 인 시리즈 8, 볼륨 16 의 Annales des sciences natelles, Huitieme série – Botanique.
- Ochnaceae(정확하게 검색) 위치: 이름 위치: 트로피코스 위치: 과학과 보존 위치: 미주리 식물원
- L. 왓슨과 M.J. Dallwitz(1992년 이후). 꽃이 피는 식물의 과: 설명, 일러스트, 식별, 정보 검색.http://delta-intkey.com
- 오크나과의 속 목록 위치: 쌍떡잎식물 위치: 패밀리의 속 목록 위치: 관상식물과 및 속 위치: 체크리스트에 대해서 위치: 선택한 식물군의 세계 체크리스트 위치: 플랜트명 위치: 데이터 소스 위치: ePIC 위치: 데이터베이스 위치: 자원 및 데이터베이스 위치: 과학적 연구 및 데이터 위치: 과학과 보존 위치: 큐 가든스
- 신열대성 오크나과 위치: 패밀리 인덱스 위치: 네오트로피키 위치: 프로젝트 및 프로그램 위치: 열대 아메리카 프로젝트 위치: 큐의 깊이 위치: 과학 연구 및 데이터 위치: 큐 가든스
- 오크나과 위치: 말피히알레스 위치: 목련목 위치: 기관지학 위치: 식물 위치: 글로벌 종
- 오크나과 위치: 혈관배양액 위치: 둘러보다 위치: 플랜트 리스트
- 결과 표시 위치: 식물 및 속(種)이 선택된 계층 보고서 위치: 데이터 액세스 위치: 그렇다.
- 말피히알레스 위치: 장미 위치: 핵심 유디코트(Gunneridae) 위치: 유디코트 위치: 혈관 배지 위치: 스펠마탑시다속 위치: 태생 식물 위치: 녹색 식물 위치: 진핵생물 위치: 지구상의 생명(나무의 뿌리) 위치: 트리 오브 라이프 웹 프로젝트(ToL)
- 전체 속 목록 위치: 오크나과 위치: 가족 목록 위치: GRIN의 패밀리 및 속 위치: 쿼리 위치: 식물에 대한 GREAN 분류법
- 오크나과 위치: 말피히알레스 위치: 패비드 위치: 장미 위치: 펜타페탈레 위치: 군네루스과 위치: 우디코티레돈 위치: 메산지오스페르마에 위치: 목련식물(꽃식물) 위치: 정조식물 위치: 유필로피타 위치: 기관지학 위치: 태생 식물 위치: 스트렙토피티나 위치: 스트렙토피타 위치: 비리디플랜테아과 위치: 진핵생물 위치: 분류법 위치: 유니프로트
- 오크나과 위치: 말피히알레스 위치: 패비드 위치: 장미 위치: 펜타페탈레 위치: 군네루스과 위치: 우디코티레돈 위치: 메산지오스페르마에 위치: 목련식물(꽃식물) · · · 입력 : · · · · 진핵생물 위치: 택사노미 브라우저 위치: 분류 데이터베이스 위치: 분류법 위치: 국립생명공학정보센터

{kind=link}