코세사우루스
Cosesaurus| 코세사우루스 시간 범위:중기 트라이아스기, 프리 D K N | |
|---|---|
 | |
| 코사우루스 이두의 생활 재구성 | |
| 과학적 분류 | |
| 왕국: | 애니멀리아 |
| 문: | 챠다타 |
| 클래스: | 파충류 |
| 패밀리: | †타니스트로페과 |
| 속: | †코세사우루스 엘렌버거 & 드 빌랄타 1974 |
| 종류: | § C. 방두근 |
| 이항명 | |
| †코세사우루스이두근 엘렌버거 & 드 빌랄타 1974 | |
코세사우루스는 타니스트로피과에 속하는 것으로 보이는 대룡류 파충류이다.[1]그것은 스페인의 몬트랄과 알커버 시 근처의 무셸칼크 아웃크롭에서 발견된 MGB V1이라는 작은 골격의 화석 흔적으로 알려져 있다.이 발굴물들은 약 2억4천2백만년에서 2억3천7백만년 전 트라이아스기 중기의 라디니아 시대로 거슬러 올라간다.이 표본은 마르토렐 박물관(일명 바르셀로나 박물관)에 보관되어 있으며, 현재는 바르셀로나 박물관의 일부이다.표본의 보존 상태가 좋지 않고 어린 시절일 가능성이 높기 때문에 코세사우루스의 해부학이 여러 다른 출처에 의해 잘못 식별되었다.예를 들어, 폴 엘렌버거는 1970년대에 그것이 새의 조상이라고 주장한 반면, 데이비드 피터스는 2000년에 익룡 조상이라고 주장했다.이 두 주장 모두 두 그룹의 기원에 대한 주류 과학 이론과 대조되며, 이 표본을 연구하는 다른 고생물학자들은 엘렌버거나 피터스가 [2][1]존재한다고 보고한 특징을 찾을 수 없다.따라서 엘렌버거와 피터스의 가설은 재현성이 낮기 때문에 과학적 건전성이 의심스러운 비주류 이론으로 간주된다.코세사우루스의 관계에 대한 주류 가설은 일반적으로 이것이 "프로토로사우루스", 특히 마크로네무스, 타니트라첼로스, 타니스트로피우스 또는 랑고바르디사우루스 [3][1][4]같은 긴 목을 가진 파충류와 밀접한 관련이 있는 타니스트로피이드라는 것에 동의한다.
역사
입자가 많은 퇴적암에서 보존 상태가 좋지 않기 때문에, 이 화석은 과거에 많은 고생물학자들에 의해 잘못 해석되었다.표본의 첫 번째 기술자인 폴 엘렌버거는 새와 같은 특징을 잘못 관찰했는데, 이는 이후의 연구에서는 확인할 수 없었다.그는 1974년, 1977년,[5][6] 1993년에 표본에 대한 몇 가지 분석을 발표했다.그는 그것이 새의 조상이라고 가설을 세웠지만, 이 가설은 곧 다른 고생물학자들에 의해 버려졌다.
예를 들어, 1979년 폴 올슨은 작은 타니스트로페이트 타니트라첼로스를 설명하는 동안 표본에 대해 간략히 논의했다.그는 엘렌버거의 사진을 사용하여 표본을 진단하는 데 어려움을 겪었지만,[3] 새보다는 타니트라첼로스나 타니스트로피우스 같은 타니스트로피드들과 많은 유사점을 공유한다는 것을 알아챘다.
엘렌버거의 가설은 20세기 후반에 실루로사우루스 [7]공룡들 사이에서 조류의 기원에 대한 증거가 늘어남에 따라 받아들여지지 않게 되었다.이 기간 동안, 1984년 호세 루이스 산즈와 니에베스 로페스 마르티네즈가 수행한 엘렌버거 이후 코세사우루스에 대한 가장 포괄적인 연구입니다.그들은 엘렌버거가 코세사우루스를 새와 연결시키는 데 사용한 특징들 중 거의 모든 것이 타당하지 않다는 것을 발견했다.그들은 또한 그것의 비율을 연구했고 그것이 어린 표본일 가능성이 높다는 결론을 내렸다.이 가설을 바탕으로, 그들은 성인 코세사우루스가 어떻게 생겼는지 보여주는 골격도를 구성했고, 파충류의 비율이 발달하는 동안 어떻게 변하는지 고려했다.산츠와 로페스 마르티네스는 코사우루스의 가장 가까운 친척이 마크로크네무스와 [2]같은 프롤라케르티드였을 것이라는 것을 알아냈다.
2000년에 연구원 데이브 [8]피터스가 엘렌버거의 가설의 변형을 발표했다.피터스는 엘렌버거가 코사우루스와 새의 연관성을 주장한 것과 비슷한 특징을 표본에서 발견했다고 주장했다.하지만, 피터스는 이러한 추정적 특징들이 코사우루스가 새보다는 익룡의 조상임을 보여준다고 한다.그러나 이 표본을 조사한 고생물학자들은 피터스의 주장을 뒷받침할 증거를 찾을 수 없었고 코세사우루스와 익룡 사이의 연관성이 형편없이 구성된 계통학 [9]분석에 의존했다는 점에 주목했다.네스빗(2011년)[10]과 에즈쿠라(2016년)[11] 등의 연구에서 설명되고 주창된 바와 같이 익룡은 코세사우루스와 같은 '원생룡'과 밀접한 관련이 없고 공룡과 밀접한 관계가 있는 발달된 대룡이라는 것이 공통된 견해다.2009년, Peters는 디지털 추적 알고리즘을 사용하여 프리큐브 뼈와 프릴 같은 피부 플랩과 같은 특징에 대한 그의 주장을 뒷받침하는 2000년 연구에 대한 후속 연구를 발표했습니다.하지만, 이 방법은 개인적인 관찰이나 CT 스캔 [12][9]증거보다는 문헌에 있는 사진에 의존하기 때문에 많은 비판을 받아왔다.
2016년 프랑코 살러가 발표한 박사학위 논문도 코세사우루스의 근골격계를 주로 다루고 있다.이탈리아어로 작성된 이 논문은 엘렌버거나 피터스가 주장하는 표본의 특정 특징, 예를 들어 선골에 흉골이나 두 개 이상의 척추뼈가 있다는 것을 밝혀냈다.그것은 또한 이 [1]속 구성원의 해부학과 생물학에 추가적인 의미를 부여했다.
묘사
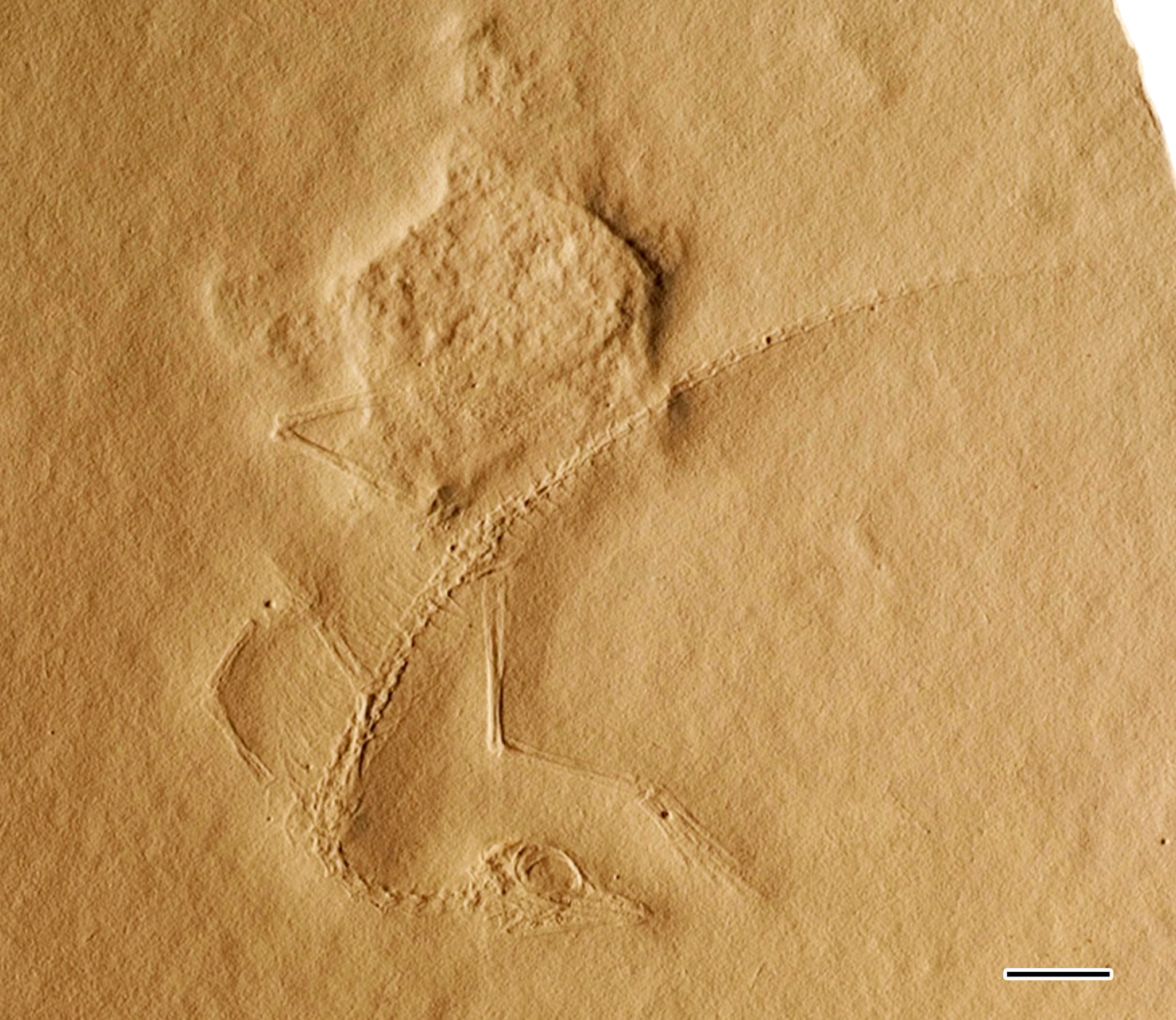
표본의 길이는 14cm(5.5인치)로 추정되지만 꼬리 끝의 자국은 희미하기 때문에 이 길이는 약간 더 길었을 수 있다.시료의 보존 상태가 좋지 않기 때문에 명확한 특징을 찾기 어렵다.엘렌버거는 뼈 [6]흔적 사이에 물갈퀴가 있는 발과 깃털 자국이 있다고 보고했고, 피터스(2000)는 엘렌버거가 대신 파타지움이나 액틴 섬유와 같은 피부 잔해를 관찰했다고 주장했다.그러나 이 두 가지 연조직에 대한 주장은 모두 의심스럽다. 왜냐하면 표본을 분석한 다른 고생물학자들은 뼈 근처의 암석 표면의 질감을 다른 암석 [2][1]슬래브와 구별할 수 없다고 설명한다.
해골
코는 적당한 길이로, 강골투석 같은 초기 수각류 공룡의 상태만큼 길지는 않지만, 초기 [2]양막류만큼 짧지는 않다.두개골에 대한 자세한 내용은 거의 확실하게 확인할 수 없습니다.두개골과 특히 눈은 비교적 컸지만, 이것들은 어린 시절의 특징일 가능성이 높다.돔처럼 생긴 두개골 지붕은 뒤통수를 향해 넓어지기 전에 눈 위에서 가장 얇았다.입에는 길고 뾰족한 상악전부에 약 6-7개 정도의 작은 이빨이 있었다.입 뒤쪽에 있는 몇 개의 치아는 턱의 나머지 얇은 [1]치아에 비해 크고, 삼각형이며, 칼날처럼 생겼다.두개골의 인상은 찌그러져 있지만, 눈 앞의 작은 움푹 패인 곳이 눈에 띈다.엘렌버거(1977)는 이 움푹 패인 곳이 코에 뚫려 안와 전회전을 나타낸다고 주장한 반면, 피터스(2000)는 이것이 일련의 회전을 나타낸다고 주장했다.그러나 살러(2016년)의 정밀 조사 결과 우울증의 깊이와 그것을 [1]둘러싼 뼈는 알 수 없었다.턱 관절은 코세사우루스가 아레오셀리디아와 바이겔티사우루스 같은 원시적인 암니오테 집단 대신 도마뱀, 공룡, 악어 같은 현대의 파충류와 비슷한 사우리아인이었음을 보여주는 몇 가지 특징을 가지고 있다.이러한 특징에는 하악골의 후관절 과정과 [13]두개골 외부에서 볼 수 있고 뒤에서 절단된 4중 뼈의 소유가 포함됩니다.그러나 4중 속도(따라서 두개골 키네시스 가능성)의 유연성은 불분명하다.[2]
척추뼈
목은 적당히 길며, 두개골의 길이와 거의 같고 몸 길이의 절반 이상이 된다.그것은 초기 공룡, 프로라세르타, 마크로크네무스와 같은 다양한 다른 대룡류와 마찬가지로 7~8개의 길쭉한 척추뼈로 형성된다.다른 "프로토사우루스"와 같지만, 새나 익룡과는 달리, 경추(목 척추)는 매우 길고 얇은 경추 늑골에 붙어 있다.다른 타니스트로피드에 비해 목이 짧지만, 산츠 & 로페스 마르티네스(1984년)와 살러(2016년)는 타니스트로피드의 목이 나이가 들면서 비례적으로 길어지는 경향이 있다고 지적한다.따라서 코세사우루스의 짧은 목은 (타니스트로페이트 기준으로는) 어린 시절의 [2][1]특징일 가능성이 높다.
인체의 척추뼈는 경추보다 짧았고, 키가 거의 같은 길이였다.등뼈는 짧고 가늘었다.엘렌버거(1977년)는 코세사우루스가 세 개의 융합된 천골(엉덩이) 척추를 가지고 있다는 것을 발견한 반면 피터스(2000년)는 네 개가 존재한다고 주장했다.그러나 이 두 주장 모두 살러(2016년)에 의해 밝혀졌다.그는 코세사우루스의 엉덩이 부분에 세 개의 천골 척추를 둘 수 있는 공간이 있다는 것을 지적했지만, 그는 또한 두 쌍의 천골 갈비뼈만이 존재했고, 보존의 질이 너무 떨어져 엘렌버거나 피터스의 주장을 뒷받침할 천골 내의 봉합사를 알아볼 수 없다는 것을 보여주었다.따라서 코세사우루스는 대부분의 [1]파충류와 비슷한 두 개의 선골 척추만을 가지고 있었을 것이다.
꼬리는 40개 이상의 길쭉한 척추뼈로 이루어져 있다.처음 몇 마리는 얇은 갈비뼈를 가지고 있었는데,[1] 이 갈비뼈의 길이는 점차 줄어들었다.엘렌버거는 척추뼈가 막대기 모양의 피그고스타일에 융합되어 혈가시가 없다고 주장했지만, 나중에 산츠 & 로페스 마르티네즈(1984년)[2]와 살러(2016년)에 의해 둘 다 밝혀졌다.
가슴 거들 및 앞다리
가슴(어깨) 거들은 코세사우루스 표본에서 가장 일반적으로 잘못 식별된 영역 중 하나입니다.엘렌버거(1977년)는 새와 같은 흉골(가슴판)과 퍼큘라(위시본)가 존재한다고 주장했지만 산츠 & 로페스 마르티네즈(1984년)[2]는 이러한 특징을 발견하지 못했다.살러(2016)는 엘렌버거의 오인식을 상세히 설명했다.그는 "furcula"는 다른 파충류처럼 사용되지 않은 쇄골의 한 쌍일 뿐이고 "sternum"은 단순히 코라코이드 뼈의 둥근 아랫부분이라고 설명했다.피터스(2000년)도 엘렌버거의 흉골(sternum)을 코사우루스(Cosaurus)[1]에서 뼈의 존재에 대한 증거를 찾을 수 없었기 때문에 흉골(sternum)이 존재한다고 추정하는 것은 틀렸다.Olson(1979)은 타니스트로피드의 [3]어깨 거들과의 연결을 그렸다.
앞다리는 뒷다리보다 훨씬 작지만 대부분의 파충류 기준으로 보면 여전히 적당한 크기이다.상완골(상완골)은 길고 곧은 편평한 관절 표면과 가운데가 뼈의 끝보다 약간 얇다.요골과 척골(하완골)은 구조가 비슷하다.그 손은 매우 길고, 다섯 개의 얇은 손가락이 있다.세 번째 손가락이 가장 길고, 손과 결합했을 때의 길이가 팔의 [1]길이를 초과합니다.
골반 거들 및 뒷다리
고관절 부위와 골반 거들이 잘 보존되어 있지 않습니다.아마도 골반에서 유일하게 보이는 장골은 [1]코세사우루스에서 비정상적으로 길쭉하다.올슨(1979)은 타니트라첼로스와 타니스트로피우스의 "헤테로토픽" 뼈와 유사한 크고 구부러진 뼈들이 꼬리 밑부분에 있었을 수 있다고 언급했다.이 뼈들은 생식 과정과 관련이 있을 수 있다.그러나 올슨은 코사우러스에 대한 그의 관찰이 엘렌버거의 원래의 거친 사진들로 촉진되었고, 그러한 방법을 사용하여 이루어진 새로운 관찰은 [3]의심스러울 수 있다고 설명했다.피터스는 코세사우루스에서 익룡과 유사한 것으로 추정되는 '프리푸빅 뼈'를 확인했지만 대런 나이쉬는 피터스의 [9]사진을 이용해 그러한 특징에 대한 명확한 증거를 찾을 수 없었고 살러(2016년)는 이 표본을 연구하는 [1]동안 피터스의 주장에 대해 언급하지 않았다.
뒷다리는 앞다리와 몸통 전체보다 길었고, 이는 동물의 생물학에 영향을 미쳤을 수도 있다.다리뼈는 길고 곧았으며, 팔뼈와 구조가 비슷했다.대퇴골의 근위부(가느다란 뼈)가 원위부보다 넓어 타니스트로피드의 경우와 유사했다.다섯 개의 중족골(주족골) 중 네 개는 길고 곧으며 촘촘하게 뭉쳐 있었다.그들은 끝이 뾰족한 긴 발가락과 연결되었고, 네 번째 발가락이 가장 길었다.다섯 번째(가장 바깥쪽) 중족골은 훨씬 짧고 넓어서 "후크" 모양을 갖게 됩니다.Olson(1979년)에 따르면, 발의 다섯 번째 발가락은 수축된 다섯 번째 [3]중족골에 대항하기 위해 긴 중족골과 같은 첫 번째 중족골(발가락 뼈)을 가지고 있었을 수 있다.이 기능은 Macrocnemus를 [11]제외하고 대부분의 타니스트로피드에 고유합니다.올슨의 의심은 살러(2016)의 표본 [1]연구에 의해 나중에 확인되었다.
분류
1977년 엘렌버거는 코세사우루스가 현대 조류들의 [6]조상이라고 제안했다.이 출판물은 새와 같은 수각류인 데이노니쿠스의 묘사를 따랐지만, 수각류 조상이 널리 받아들여지기 훨씬 전에 나타났다.이 출판물에서 엘렌버거는 화석의 특징들을 다음과 같이 해석했다: 끈 같은 견갑골, 털뼈, 용골 흉골, 부리 같은 턱, 역행하는 치골 그리고 꼬리 깃털.하지만 엘렌버거(1977) 이후 코세사우루스를 포함한 다른 모든 과학적 연구는 이러한 특성들 중 많은 것들이 잘못 확인되었고 새와 관련이 있을 것 같지 않다는 것을 발견했다.
엘렌버거(1977년) 이후의 연구는 코사우루스를 "프롤라케르티폼" 또는 "프로토사우루스"로 해석했다.이 용어들은 Prolacerta, Macrocnemus, Tanystropheus와 같이 목이 긴 파충류들의 특정 그룹을 가리킨다.이 그룹은 내부적으로도, 외부적으로도(파충류 가계도에 속하는) 많은 개정을 거쳤다.20세기 후반 이전, "프롤라케르티폼"은 렙틸리아 주변에서 섞였고, 일부 자료들은 그들을 도마뱀의 조상이라고 추정했다.이러한 출처에는 코세사우루스, 산츠 및 로페스 마르티네스의 재기술자(1984)[2]가 포함된다.하지만 1980년대 후반에는 "프롤라케르티폼"을 대룡류 동물로 배치하는 것에 대한 합의가 이루어졌는데, 이는 그들이 도마뱀이나 [14][13]뱀 같은 레피도사우루스보다는 악어나 공룡 같은 대룡류에 더 가깝다는 것을 의미한다.
그러나 1998년 데이비드 딜케스가 프롤라케르타가 실제로는 다른 프롤라케르타상과 가까운 친척이 아니라 더 왕관 쪽으로 [15]향하는 아치오사우루스라는 결론을 내리면서 프롤라케르타라는 용어는 무너졌다.이 발견으로 인해 "Prolacertiformes"라는 용어는 한때 Prolacerta와 가까운 것으로 여겨졌던 많은 파충류들을 포함하는 "Protorosauria"로 대체되었다.코사우러스는 원시룡류들 사이에서 보존되었지만, 코사우러스는 단편적인 특성 때문에 보통 계통학적 분석으로는 실험되지 않는다.올슨(1979)은 타니트라첼로스와 타니스트로페우스를 [3]포함한 아마도 반수생으로 보이는 원생 공룡 집단인 타니스트로피드 사이에 거짓말을 했다고 주장했다.이 해석은 2017년 [4]Ezcurra 등에 의해서도 뒷받침되었다.Sanz & Lopez-Martinez(1984)는 마크로크네무스와 프로라세르타와 같은 "프롤라세티드"로 식별했지만, 이 용어는 주로 "프롤라세티폼"과 유사한 이유로 폐기되었다.마크로크네무스는 또한 현재 타니스트로페이드로 분류된다.타니스트로피드는 때때로 [11]프로라세르타와 같은 정도는 아니지만 다른 "프로토사우루스"의 왕관을 쓴 것으로 발견되기도 한다.
Sanz & Lopez-Martinez(1984)는 그들의 비례 [2]분석을 근거로 코사우루스의 가장 가까운 친척이 마크로크네무스라고 주장했지만, Olson(1979년)과 Saller(2016년)는 각각 코사우루스를 타니스트로페과(Tanystropheidae)의 더 발전된 멤버로 분류했다.그들은 다섯 번째 발가락의 중족골과 같은 첫 번째 골격은 특히 [3][1]마크로크네무스와의 연결을 무효화한다고 주장했다.
살러(2016)의 논문은 "원생룡"의 관계를 연구하기 위해 이전 저자들과는 다른 두 가지 계통발생학적 분석을 사용했다.이러한 분석 중 첫 번째는 "Prolacertiformes"[15]의 단동학을 분해한 것으로 유명한 연구인 Dilkes(1998)에서 나왔다.그러나 더 많은 타니스트로피드를 포함하도록 분석을 수정하고 Saller에 의해 실행했을 때 결과는 매우 불확실했다.살러의 가장 인색한 나무들(MPT)은 마크로크네무스보다 더 진보된 타니스트로피드의 구조에 동의하지 않았다.MPT는 가장 적은 수의 "단계"를 포함하는 가계도이다(특성이 획득되고 상실되는 진화적 전환).Occam's 면도기의 지침에 따르면 MPT는 진화적 관계를 가장 정확하게 묘사합니다.그러나 살러의 6개의 MPT는 타니스트로피과의 구조에 대해 의견이 엇갈렸다.6개의 MPT에 의해 발견된 유일한 결과는 Macrocnemus가 그 [1]과의 가장 기초적인 멤버라는 것이었다.
좀 더 결정적인 결과를 얻기 위해 살러는 특이한 드레파노사우루스 메가란코사우루스를 분석에 추가했다.분석의 일부(예를 들어 아르코사우루스의 구조가 에즈쿠라[2016][11]에 의해 발견된 구조와 비슷함)가 더 결정적이긴 했지만, 프로토사우루시아와 타니스트로페이대의 내부 구조는 메가란코사우루스 없이 분석을 수행했을 때보다 더 결정적이지는 않았다.
이 문제는 살러가 두 번째 계통 분석을 사용하도록 자극했다.이 분석은 원래 프리처드 외 연구진(2015)에 의해 작성되었으며,[16] 특히 타니스트로피드에 초점을 맞췄다.이 분석에서 3개의 MPT가 발견되었습니다.Archosaurforma의 구조는 다시 한번 결정적이지 않았지만, 세 MPT 모두 Tanystropheidae의 구조와 일치했다.가장 흥미로운 결과 중 하나는 코사우루스가 두 발로 걷고 초식했을지도 모르는 특이한 타니스트로피이드인 랑고바르디사우루스의 자매 분류군으로 배치되었다는 것입니다.이러한 연관성은 세 가지 시나포모형(유래된 구별 특징)에 의해 알려졌습니다: 평평한 앞관절을 가진 경추와 등뼈, 낮은 신경 가시를 가진 등뼈, 그리고 잘 확장되지 않은 [1]대퇴골 원위부 결절.
다음은 살러(2016년)가 프리차드 등(2015년)을 이용한 분석 결과다.다음 분해도는 엄격한 합의 나무이며, 이는 특정 [1]MPT에 동의하지 않기 위해 경합 영역을 폴리토미(Polytomy)로 감소시키는 세 가지 MPT의 조합임을 의미한다.
| 대룡류 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
고생물학
이동
코세사우루스 표본의 뒷다리는 앞다리보다 길어서, 두 발로 걸었을 가능성이 있지만 확실하지는 않습니다.Sanz & Lopez-Martinez(1984)는 가능성을 조사하기 위해 "Galton 지수"로 알려진 비례 도구를 사용했다.피터 갈튼이 1976년 "프로사우로포드"에 대한 연구에서 시작한 이 지수는 뒷다리 전체의 길이를 동물의 몸 전체 길이와 비교합니다(목이나 꼬리를 포함하지 않음).코세사우루스 표본의 비율은 1.21로, 이는 4족보행 eolacertian(원시적 스쿼메이트 친척)보다는 크지만, 실로피시스와 같은 2족보행 초기 수각류보다는 작다.이것은 어느 정도 이족 보행의 움직임을 나타낼 수 있지만, 이 표본은 어린 개체로 추정되기도 한다.현대 악어들의 경우, 갈톤지수는 부화기와 성체 사이에서 1.06에서 0.79로 줄어든다.만약 코세사우루스 개체에서 그 비율이 같은 정도로 줄어든다면,[2] 그들은 성인이 되었을 때 이족보행을 허용하는 범위를 벗어났을 것이다.
레퍼런스
- ^ a b c d e f g h i j k l m n o p q r s Saller, Franco (15 April 2016). "Anatomia, paleobiologia e filogenesi di Macrocnemus bassanii Nopcsa 1930 (Reptilia, Protorosauria)" (PDF). Dissertation Thesis: 1–206. doi:10.6092/unibo/amsdottorato/7449.
- ^ a b c d e f g h i j k Sanz, J.L.; López-Martínez, N. (1984). "The prolacertid Lepidosaurian Cosesaurus aviceps Ellenberger & Villalta, a claimed "Protoavian" from the Middle Triassic of Spain". Geobios. 17 (6): 747–755. doi:10.1016/S0016-6995(84)80119-9.
- ^ a b c d e f g Olson, Paul E. (3 May 1979). "A new aquatic eosuchian from the Newark Supergroup (Late Triassic – Early Jurassic) of North Carolina and Virginia" (PDF). Postilla. 176: 1–13.
- ^ a b Ezcurra, Martin; Fortuny, Josep; Mujal, Eudald; Bolet, Arnau (2017). "First direct archosauromorph remains from the Early-Middle Triassic transition of the Iberian Peninsula" (PDF). Palaeontologia Electronica. 20 (3): 1–10. doi:10.26879/686.
- ^ Ellenberger, Paul; de Villalta, José F. (1974). "Sur la présence d'un ancètre probable des Oiseaux dans le Muschelkalk supérieur de Catalogne (Espagne). Note préliminaire". Acta Geológica Hispánica. 9 (5): 162–168.
- ^ a b c Ellenberger, P.-P. (1977). "Quelques precisions sur l'anatomie et la place systematique tres speciale de Cosesaurus aviceps (Ladinien superieur de Montral, Catalogne)". Caud. Geologica Iberica. 4: 169–188.
- ^ Padian, K.; Chiappe, L.M. (1998). "The origin and early evolution of birds". Biological Reviews. 73 (1): 1–42. doi:10.1111/j.1469-185X.1997.tb00024.x. S2CID 86007060.
- ^ Peters, David (December 2000). "A reexamination of four prolacertiforms with implications for pterosaur phylogenesis". Rivista Italiana di Paleontologia e Stratigrafia (Research in Paleontology and Stratigraphy). 106 (3): 293–336. ISSN 2039-4942.
- ^ a b c Naish, Darren (3 July 2012). "Why the world has to ignore ReptileEvolution.com". Scientific American Blog Network. Archived from the original on 7 February 2017.
- ^ Nesbitt, S.J. (2011). "The early evolution of archosaurs: relationships and the origin of major clades". Bulletin of the American Museum of Natural History. 352: 1–292. doi:10.1206/352.1. hdl:2246/6112. S2CID 83493714.
- ^ a b c d Ezcurra, Martín D. (2016-04-28). "The phylogenetic relationships of basal archosauromorphs, with an emphasis on the systematics of proterosuchian archosauriforms". PeerJ. 4: e1778. doi:10.7717/peerj.1778. ISSN 2167-8359. PMC 4860341. PMID 27162705.
- ^ Bennett, S. C. (2005). Pterosaur science or pterosaur fantasy? (PDF). Prehistoric Times. Vol. 70. pp. 21–23, 40.
- ^ a b Evans, Susan E. (1988). "The early history and relationships of the Diapsida". In Benton, Michael J. (ed.). The Phylogeny and Classification of the Tetrapods. Oxford: Clarendon Press. pp. 221–260.
- ^ Benton, Michael J. (June 1985). "Classification and phylogeny of the diapsid reptiles". Zoological Journal of the Linnean Society. 84 (2): 97–164. doi:10.1111/j.1096-3642.1985.tb01796.x.
- ^ a b David M. Dilkes (1998). "The Early Triassic rhynchosaur Mesosuchus browni and the interrelationships of basal archosauromorph reptiles". Philosophical Transactions of the Royal Society of London, Series B. 353 (1368): 501–541. doi:10.1098/rstb.1998.0225. PMC 1692244.
- ^ Pritchard, Adam C.; Turner, Alan H.; Nesbitt, Sterling J.; Irmis, Randall B.; Smith, Nathan D. (2015-03-04). "Late Triassic tanystropheids (Reptilia, Archosauromorpha) from northern New Mexico (Petrified Forest Member, Chinle Formation) and the biogeography, functional morphology, and evolution of Tanystropheidae". Journal of Vertebrate Paleontology. 35 (2): e911186. doi:10.1080/02724634.2014.911186. ISSN 0272-4634. S2CID 130089407.
외부 링크
| |||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||








{kind=link}
{kind=link}