우위(유전자)
Dominance (genetics)
유전학에서 우위는 유전자의 한 변종(알레)이 염색체를 가리는 현상 또는 [1][2]염색체의 다른 복제에 대한 동일한 유전자의 다른 변종의 영향을 덮어쓰는 현상이다.첫 번째 변종은 우성이고 두 번째 변종은 열성이다.각 염색체에 동일한 유전자의 두 가지 다른 변형을 갖는 이 상태는 원래 새로운 유전자(de novo) 또는 유전 유전자 중 하나의 돌연변이에 의해 야기된다.상염색체 우성 또는 상염색체 열성이라는 용어는 비성염색체(autosomes)와 그 관련 특징에 대한 유전자 변형을 설명하기 위해 사용되는 반면, 성염색체(allosomes)는 X-연계 우성, X-연계 열성 또는 Y-연계 우성으로 불린다. 이것들은 부모의 성별에 따라 달라지는 유전과 발현 패턴을 가지고 있다.및 아이(성관계 참조).Y염색체의 복사본이 하나밖에 없기 때문에 Y염색체는 지배적이거나 열성적일 수 없다.또한, 유전자 변이가 양쪽 염색체에 존재할 때와 비교하여 부분적인 영향을 미치는 불완전한 우성, 그리고 각 염색체의 다른 변종들이 둘 다 연관된 특성을 보이는 공동 우성과 같은 다른 형태의 우성들이 있다.
지배력은 멘델의 유전과 고전 유전학의 핵심 개념이다.문자와 푸넷 사각형은 교수법에서 지배의 원리를 보여주기 위해 사용되며, 지배적 대립 유전자에 대문자, 열성 대립 유전자에 소문자를 사용하는 것은 널리 사용되는 관례이다.지배력의 전형적인 예는 완두콩의 종자 모양 유전이다.완두콩은 동그랗고 대립 유전자 R과 관련되거나 주름지고 대립 유전자 R과 관련될 수 있다.이 경우 RR, Rr 및 rr의 세 가지 대립 유전자 조합(유전자형) 조합이 가능합니다.RR(호모 접합) 개체는 둥근 완두콩을 가지고 있고, RR(호모 접합) 개체는 주름진 완두콩을 가지고 있습니다.Rr(헤테로 접합) 개체에서 R 대립 유전자는 r 대립 유전자의 존재를 가리기 때문에 이러한 개체는 둥근 완두콩도 가지고 있다.따라서 대립 유전자 R이 대립 유전자 R보다 우세하고 대립 유전자 R이 대립 유전자 R에 대해 열성적이다.
지배력은 대립 유전자나 그 특성(표현형)에 내재하지 않는다.그것은 어떤 기능에서든 주어진 유전자의 두 대립 유전자 사이에 엄밀하게 상대적인 영향이다; 하나의 대립 유전자는 같은 유전자의 두 번째 대립 유전자에 대해 지배적일 수 있고, 세 번째 대립 유전자에 대해서는 열성적일 수 있고, 네 번째 대립 유전자에 대해서는 공동 지배적일 수 있다.또한 하나의 대립 유전자가 하나의 특징에 지배적일 수 있지만 다른 특징에는 지배적이지 않을 수 있습니다.
우성은 한 유전자의 대립 유전자가 다른 [3]유전자의 대립 유전자의 효과를 가리는 현상인 인식론과는 다르다.
배경
_(14582377398).jpg)
지배의 개념은 그레고르 요한 멘델에 의해 도입되었다."유전학의 아버지"인 멘델이 1860년대에 이 용어를 처음 사용했지만, 20세기 초까지 널리 알려지지 않았다.멘델은 원예 완두콩의 다양한 특징들이 씨앗, 종자 꼬투리, 그리고 식물의 생김새와 관련이 있다는 것을 관찰했는데, 둥근 종자 대 주름종자, 노란색 대 녹색 종자, 붉은 꽃 대 흰 꽃 또는 키가 큰 식물 대 키가 작은 식물과 같은 두 개의 분리된 표현형이 있었다.따로따로 사육하면, 그 식물들은 대대로 같은 표현형을 만들어냈다.그러나 서로 다른 표현형을 가진 선이 교배(교배)되면 부모 표현형 중 하나만이 자손에게 나타났다(녹색, 원형, 적색, 키).그러나, 이러한 잡종 식물이 교배되었을 때, 자손 식물은 특징적인 3:1 비율로 두 개의 원래 표현형을 보여주었고, 더 일반적인 표현형은 부모 잡종 식물의 표현형이다.멘델은 첫 번째 십자가의 각 부모가 서로 다른 대립 유전자(한 부모 AA와 다른 부모 aa)에 대한 호모 접합체이며, 그 결과 이 모든 잡종이 헤테로 접합체(Aa)였고, 하이브리드 교차 발현에서 두 대립 유전자 중 하나가 다른 대립 유전자를 지배했다고 추론했다.두 heterozygotes(Aa XAa)는 마지막 부분에 십자를 긋다 2대 1로:2교시는(A)에 표현형을 보이는 것으로 나타나 1유전자형 비율에서, 그리고 마지막으로(를)에 표현형이 3표현형 비율을 보여 주는 AA, Aa, 괴상 용암 자손을 생산할 것이다.
멘델은 나중에 소개된 유전자, 대립 유전자, 표현형, 유전자형, 호모 접합체, 헤테로 접합체라는 용어를 사용하지 않았다.그는 지배적 대립 유전자와 열성 대립 유전자에 대문자 표기법과 소문자 표기법을 도입했는데, 각각 오늘날에도 여전히 사용되고 있다.
1928년, 영국의 인구 유전학자 로널드 피셔는 지배가 수식어 유전자의 기여를 통해 자연 도태에 기초해 작용한다고 제안했다.1929년, 미국의 유전학자 Sewall Wright는 지배력은 단순히 신진대사 경로와 관련된 유전자의 상대적 필요성의 생리적 결과라고 말함으로써 응답했다.라이트의 설명은 유전학에서 확립된 사실이 되었고, 논쟁은 대부분 끝났다.그러나 [4][5][6]일부 특성은 진화 메커니즘에 의해 지배력이 영향을 받을 수 있습니다.
염색체, 유전자 및 대립 유전자
대부분의 동물들과 몇몇 식물들은 쌍으로 된 염색체를 가지고 있고, 이배체로 묘사된다.그들은 각 염색체의 두 가지 버전을 가지고 있는데, 하나는 엄마의 난자에 의해 기여되고 다른 하나는 생식체로 알려져 있고, 반수체로 묘사되며 감수분열을 통해 만들어진다.그리고 나서 이 생식체들은 수정하는 동안 성생식을 하는 동안 여러 번 분열하는 새로운 단일 세포 접합체로 융합되어 부모 세포와 같은 수의 염색체를 가진 새로운 유기체가 된다.
일치하는 (상동) 쌍의 각 염색체는 구조적으로 서로 유사하며 매우 유사한 DNA 염기서열(위치, 단일한 궤적)을 가지고 있다.각 염색체의 DNA는 다양한 특징에 영향을 미치는 일련의 이산 유전자로 기능합니다.따라서, 각각의 유전자는 또한 대응하는 상동성을 가지고 있으며, 대립 유전자로 불리는 다른 형태로 존재할 수 있다.두 개의 상동 염색체 상의 같은 위치에 있는 대립 유전자는 동일하거나 다를 수 있다.
인간의 혈액형은 염색체 9번(9q34.2)[7]의 긴 팔이나 q 팔에 위치한 A, B, AB 또는 O 혈액형을 만드는 효소의 변형을 코드하는 ABO 유전자에 의해 결정된다.이 궤적에 존재할 수 있는 세 가지 다른 대립 유전자가 있지만, 어느 개인에게나 존재할 수 있는 것은 어머니로부터 물려받은 것과 [8]아버지로부터 물려받은 두 가지뿐입니다.
주어진 유전자의 두 개의 대립 유전자가 동일하면, 유기체는 호모 접합체라고 불리며, 그 유전자에 관해 호모 접합체라고 불리며, 대신 두 개의 대립 유전자가 다르면, 유기체는 헤테로 접합체이며 헤테로 접합체이다.단일 궤적이나 모든 유전자를 통틀어 유기체의 유전자 구성은 유전자형이라고 불린다.유기체의 유전자형은 직간접적으로 분자, 물리적, 그리고 개별적으로 또는 집합적으로 표현형이라고 불리는 다른 특징에 영향을 미친다.헤테로 접합 유전자 자리에서 두 개의 대립 유전자가 상호작용하여 [citation needed]표현형을 생성한다.
우위의 종류
완전한 우위
완전 우성에서는 헤테로 접합 유전자형에서 하나의 대립 유전자의 효과는 다른 유전자의 효과를 완전히 가린다.마스크가 다른 대립 유전자에 지배적인 것으로 간주되고 마스크된 대립 유전자는 [9]열성 유전자로 간주됩니다.헤테로 접합체의 표현형에서 완전한 우위는 지배적인 호모 접합체의 표현형과 구별할 수 없다.
완전한 우위의 전형적인 예는 완두콩의 종자 모양(pea 모양)의 유전이다.완두콩은 둥글거나(알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알이 경우, RR, Rr, Rr. RR 및 rr는 호모 접합, Rr은 헤테로 접합의 세 가지 대립 유전자 조합이 가능하다.RR 개인은 둥근 완두콩을 가지고 있고 RR 개인은 주름진 완두콩을 가지고 있다.Rr 개체에서 R 대립 유전자는 r 대립 유전자의 존재를 가리기 때문에, 이러한 개체는 둥근 완두콩도 가지고 있다.따라서 대립 유전자 R은 대립 유전자 r에 대해 완전히 우세하고 대립 유전자 r은 대립 유전자 R에 대해 열성적이다.
불완전한 우위

불완전 우위(부분 우위, 반우위 또는 중간 유전이라고도 함)는 헤테로 접합 유전자형의 표현형이 호모 접합 유전자형의 표현형과 구별되고 종종 중간일 때 발생한다.표현형 결과는 종종 헤테로 접합 상태의 혼합된 형태의 특성으로 나타난다.예를 들어 스냅드래곤 플라워 색상은 빨강 또는 하양 중 어느 쪽이든 호모 접합입니다.붉은 호모 접합성 꽃이 흰 호모 접합성 꽃과 짝을 이루면 분홍색 스냅드래곤 꽃이 만들어집니다.핑크 스냅드래곤은 불완전한 지배력의 결과이다.흰 꽃과 붉은 꽃의 부모 품종이 교배했을 때 분홍색이 생기는 4시 식물에서도 비슷한 불완전 우성이 발견된다.표현형이 측정되어 수치적으로 처리되는 정량유전학에서 헤테로 접합체의 표현형이 정확히 두 호모 접합체의 표현형 사이에 있으면 표현형은 전혀 우세를 보이지 않는다고 한다. 즉, 헤테로 접합체의 표현형 측정치가 다른 호모 접합체보다 한 호모 접합체에 가까울 때만 우위가 존재한다.
F세대의1 식물이 자가 수분될 때 F세대의2 표현형 및 유전자형 비율은 1:2:1(빨간색:핑크색:흰색).[10]
공동 지배



공동 우위는 두 대립 유전자의 기여가 표현형에서 보이고 두 대립 유전자가 다른 대립 유전자를 마스크하지 않을 때 발생합니다.
예를 들어 ABO 혈액형 시스템에서 혈구 표면의 당단백질(H항원)에 대한 화학적 변형이 3개의 대립 유전자에 의해 제어되며, 그 중 2개는 서로 공우성(IA, IB)이며 ABO 궤적에서의 열성 i보다 우세하다.I와B I의 대립 유전자는 다른A 변화를 일으킨다.I에 의해A 코드화된 효소는 막 결합 H 항원에 N-아세틸갈락토사민을 첨가한다.IB 효소는 갈락토스를 첨가한다.i 대립 유전자는 수정되지 않습니다.따라서 I와B I의 대립 유전자는 각각A I에 우세하지만AA(II와A Ii의 개인은 모두 A형, II와BBB Ii의 개인은 모두 B형), II의AB 개인은 양쪽 혈구를 모두 수정하고 AB형이기 때문에 I와B I의 대립A 유전자는 공지배적이라고 한다.
또 다른 예는 헤모글로빈의 베타글로빈 성분에 대한 궤적에서 발생하며, HbA/HbA, HbS/Hb, HbSS/Hb의A 세 가지 분자 표현형은 모두 단백질 전기영동으로 구별할 수 있다.(헤테로 접합 유전자형에 의해 생성되는 의학적 상태를 겸상세포 특성이라고 하며 겸상세포 빈혈과는 구별할 수 있는 경미한 상태이므로 빈혈에 관해 대립 유전자가 불완전한 우위를 보인다).분자 수준에서 대부분의 유전자 자리의 경우, 두 대립 유전자가 RNA로 전사되기 때문에 두 대립 유전자가 공존적으로 발현된다.
표현형에서 대립 유전자 생성물이 공존하는 공동 우위는 대립 유전자 생성물의 양적 상호작용이 중간 표현형을 생성하는 불완전한 우성과 다르다.예를 들어 공도에서는 적색 호모 접합성 꽃과 백색 호모 접합성 꽃은 적색과 백색 반점을 가진 자손을 낳는다.F1 세대의 식물이 자가 수분될 때 F2 세대의 표현형 및 유전자형 비율은 1:2:1(빨간색: 반점:흰색). 이 비율은 불완전한 우위에 대한 비율과 동일합니다.다시 한 번 말하지만, 이 고전적인 용어는 부적절합니다.실제로는 그러한 용어가 지배력을 나타낸다고는 할 수 없습니다.
일반적인 오해 해소
이 섹션의 톤 또는 스타일은 Wikipedia에서 사용되는 백과사전 톤을 반영하지 않을 수 있습니다.(2022년 5월 (이를을 학습합니다) |
지배력은 유전자의 두 버전 사이의 관계에 관련된다.지배적 특성은 일반적으로 Punnett Square에서 볼 수 있는 상속 패턴과 일치합니다.만약 한 개인이 두 가지 버전의 유전자를 가지고 있다면, 다음 세대에서 자주 관찰되는 유전자는 "지배적"[11]으로 간주됩니다.
유전학에는 꽤 흔한 오해들이 몇 가지 있다.지배적인 특성은 "강함"이고 "강함"은 열성 특성이라고 생각됩니다.지배적인 특성은 또한 집단에서 더 널리 퍼질 뿐만 아니라 유전될 가능성이 더 높다고 추정됩니다.지배적인 특성이 남성적이거나 남성적이라는 생각은 또 다른 흔한 오해이다.이러한 다른 생각의 출현은 유전자가 아닌 환경에서의 지배의 다양한 개념에 기인한다.예를 들어, 강하고 강력하며 지배적이다.이것은 지배의 유전적 개념과는 다르다.[12]
지배력은 대립 유전자가 유해한지, 중성인지, 유리한지를 결정하지 않는다.그러나, 선택은 표현형을 통해 간접적으로 유전자에 작용해야 하며 지배력은 표현형의 대립 유전자의 노출에 영향을 미치므로 선택 중인 대립 유전자의 빈도가 변화한다.유해 열성 대립 유전자는 저주파수의 집단에서 유지될 수 있으며, 대부분의 복사는 이형 접합체로 운반되며, 그러한 개인에게는 비용이 들지 않습니다.이러한 희귀한 열성 질환은 많은 유전성 질환의 기초가 된다.
명명법
유전학에서 기호는 대수적 자리 표시자로 시작되었다.하나의 대립 유전자가 다른 대립 유전자에 지배적인 경우, 가장 오래된 관습은 지배적인 대립 유전자를 대문자로 상징합니다.열성 대립 유전자는 소문자로 동일한 문자가 할당됩니다.완두콩의 예에서는 두 대립 유전자의 우위 관계가 판명되면 대문자 기호 R로 둥근 형상을 생성하는 우위 대립 유전자와 소문자 기호 r로 주름 형상을 생성하는 열성 대립 유전자를 지정할 수 있다.이어서 호모 접합 우성 유전자형, 헤테로 접합 유전자형 및 호모 접합 열성 유전자형은 각각 RR, Rr 및 rr로 표기된다.또한 두 가지 대립 유전자는 W와 W, 그리고 세 가지 유전자형 WW, Ww, ww로 지정할 수 있는데, 그 중 첫 번째 두 가지는 둥근 완두콩과 세 번째 주름이 잡힌 완두콩을 생산한다.지배적 대립 유전자에 대한 기호로 "R" 또는 "W"를 선택하는 것은 호모 접합이 지배적일 때 "둥근" 또는 "주름" 표현형을 유발하는 대립 유전자가 사전 판단하지 않습니다.
유전자는 여러 개의 대립 유전자를 가질 수 있다.각 대립 유전자는 궤적 기호 뒤에 고유한 위첨자가 이어집니다.많은 종에서, 야생 개체군에서 가장 흔한 대립 유전자는 야생형 대립 유전자로 지정됩니다.위첨자로 + 문자와 함께 표시됩니다.다른 대립 유전자들은 야생형 대립 유전자에 지배적이거나 열성적이다.열성 대립 유전자의 경우 궤적 기호는 소문자로 표시됩니다.야생형 대립 유전자에 대해 어느 정도의 우위성을 가지는 대립 유전자의 경우, 궤적 기호의 첫 글자는 대문자로 한다.예를 들어, 여기 실험용 쥐의 궤적에 있는 대립 유전자들 중 일부가 있습니다y: A, 지배적인 노란색+, a, 야생형, 그리고bt a, 검정색, 황갈색.대립bt 유전자는 야생형 대립 유전자에 대해 열성이고, A 대립 유전자는y 야생형 대립 유전자에 대해 공분자이다.또한 A 대립 유전자는y 대립bt 유전자와 대동소이하지만, 관계가 생쥐 유전자 명명법의 한계를 벗어난다는 것을 보여준다.
유전학이 복잡해지면서 유전학 명명법칙이 발전해 왔다.위원회는 일부 종에 대한 규칙을 표준화했지만 모든 종에 대한 규칙은 아니다.한 종에 대한 규칙은 다른 [13][14]종에 대한 규칙과 다소 다를 수 있습니다.
다른 유전자 개념과의 관계
다중 대립 유전자
비록 어떤 이배체 유기체의 개인이 하나의 궤적에서 최대 두 개의 다른 대립 유전자를 가지고 있지만, 대부분의 유전자는 전체적으로 다수의 대립 유전자 버전으로 존재한다.만약 대립 유전자들이 표현형에 다른 영향을 미친다면, 때때로 그들의 지배 관계는 연속적으로 묘사될 수 있다.
예를 들어, 길들여진 고양이의 털 색깔은 티로시나아제 효소를 코드하는 TYR 유전자의 일련의 대립 유전자에 의해 영향을 받습니다.대립 유전자 C, cb, cs, ca(풀컬러, 버마, 샴, 알비노)는 각각 다른 수준의 색소를 생성하며, 따라서 다른 수준의 색소 희석을 생성한다.C 대립 유전자(풀컬러)는 마지막 3가지에 대해 완전히 우세하며, C 대립 유전자(알비노)는a 처음 [15][16][17]3가지에 대해 완전히 열성입니다.
상염색체 대 성관계 우위
이 섹션은 어떠한 출처도 인용하지 않습니다.(2020년 1월 (이 및 에 대해 ) |
인간과 다른 포유동물 종에서 성은 X염색체와 Y염색체라고 불리는 두 개의 성염색체에 의해 결정된다.인간 여성은 XX, 남성은 XY입니다.나머지 염색체 쌍은 양성에서 발견되고 자동 염색체라고 불립니다; 이 염색체들의 위치와 관련된 유전적 특성은 상염색체로 묘사되며, 우세하거나 열성적일 수 있습니다.X염색체와 Y염색체의 유전적 특성은 성염색체와 연결되어 있기 때문에 성연관이라고 불리며, 성별이나 성별의 특성 때문이 아닙니다.실제로, 이 용어는 거의 항상 X-연계 특성을 가리키며, 그러한 특성(예: 적-녹색 시력 결핍)은 성별의 영향을 받지 않는다.암컷은 X염색체에서 발견되는 모든 유전자 자리의 두 개의 복사본을 가지고 있습니다. 자동 염색체와 마찬가지로, 동일한 지배 관계가 적용됩니다.그러나 수컷은 각각의 X염색체 유전자 자리의 복사본을 하나만 가지고 있으며, 이러한 유전자에 대해 반수지로 묘사된다.Y 염색체는 X 염색체보다 훨씬 작고, 고환 결정 인자에 대한 SRY 유전자와 같이 '악성'에 영향을 미치는 유전자를 포함하지만 이에 한정되지 않는 훨씬 작은 유전자 세트를 포함합니다.성별 관련 유전자 위치에 대한 지배력 규칙은 암컷에서의 그들의 행동에 의해 결정된다: 수컷은 오직 하나의 대립 유전자를 가지고 있기 때문에(특정 유형의 Y 염색체 부배수의 경우 제외), 그 대립 유전자는 항상 지배적인지 아니면 열성적인지에 관계없이 발현된다.새들은 이성 염색체를 가지고 있다: 수컷 새들은 ZZ, 암컷 새들은 ZW 염색체를 가지고 있다.하지만, 특징의 유전은 그렇지 않으면 XY계를 상기시킨다; 수컷 얼룩말 핀치는 그들의 두 개의 Z 염색체 중 하나에 흰색 착색 유전자를 가지고 있을 수 있지만, 암컷은 항상 흰색 착색 유전자를 가지고 있다.메뚜기는 XO 시스템을 가지고 있다.여성은 XX를 가지고 있지만, 남성은 X만 가지고 있습니다.Y염색체는 전혀 없어요.
서신
에피타시스["epi + stasis = to on top on"]는 단일 특성에 영향을 미치는 두 개의 다른 유전자 궤적에서 대립 유전자 사이의 상호작용이며, 때때로 동일한 궤적에서 두 개의 다른 대립 유전자 사이의 지배적 상호작용과 유사할 수 있다.에피타시스는 두 개의 비 에피스타틱 유전자에 대해 예상되는 특성 9:3:3:1 비율을 수정한다.두 개의 궤적에 대해, 14개의 종류의 서신 상호작용이 인식된다.열성 서신의 예로서 한 유전자 궤적은 꽃 색소가 노란색(AA 또는 Aa)인지 녹색(aa)인지를 판정할 수 있으며, 다른 유전자 궤적은 색소가 생성되는지(BB 또는 Bb) 여부를 판정할 수 있다(bb).bb 식물에서는 AA, Aa, aa와 같은 다른 궤적의 유전자형에 관계없이 꽃이 흰색입니다.bb 조합은 A 대립 유전자에 지배적이지 않다. 오히려, B 유전자는 A 유전자에 대한 열성 서간시스를 나타낸다. 왜냐하면 열성 대립 유전자에 대한 호모 접합 시 B 궤적이 A 궤적의 표현형 발현을 억제하기 때문이다.두 개의 AaBb 식물 사이의 교배에서, 이것은 특징적인 9:3:4의 비율을 생성하며, 노란색: 녹색: 흰색 꽃의 경우이다.
우성 서간증에서는 앞의 예와 같이 하나의 유전자 궤적이 황색 또는 녹색 색소를 결정할 수 있다. 즉, AA와 Aa는 황색, aa는 녹색이다.제2의 궤적은 안료 전구체가 생성되는지 여부(dd)를 판정한다(DD 또는 Dd).여기서 DD 또는 Dd 식물에서는 지배적인 D 대립 유전자의 서한 효과 때문에 A자리의 유전자형에 관계없이 꽃이 무색이다.따라서 두 AaDd 식물 사이의 교배에서 식물의 3/4은 무색이며 노란색과 녹색 표현형은 dd 식물에서만 발현된다.이는 백색: 황색: 녹색 식물의 특징적인 12:3:1 비율을 생성한다.
보조편지는 두 개의 위치가 동일한 표현형에 영향을 미칠 때 발생한다.예를 들어 색소색이 CC 또는 Cc에 의해 생성되지만 Cc에 의해 생성되지 않으며 DD 또는 Dd에 의해 생성되지 않은 경우 색소는 cc 또는 dd 중 하나의 유전자형 조합으로 생성되지 않는다.즉, 표현형을 생성하기 위해서는 두 위치 모두 적어도 하나의 지배적 대립 유전자가 있어야 한다.이것은 착색된 식물과 비착색된 식물의 특징적인 9:7 비율을 생성합니다.이와는 대조적으로 상보적 인식은 유전자형이 cc 및 dd일 경우에만 비색소식물을 생성하며, 색소식물과 비색소식물의 [18]특징비는 15:1이다.
고전 유전학에서는 한 번에 두 유전자 사이의 서신적 상호작용을 고려했다.분자 유전학에서 이제 모든 유전자 자리는 많은 다른 유전자와의 복잡한 상호작용에 관여하고 있고 (예를 들어, 대사 경로는 수십 개의 유전자를 포함할 수 있다), 이것이 전형적인 두 개의 위치 모델보다 훨씬 더 복잡한 서신적 상호작용을 만든다는 것이 명백하다.
하디-바인버그 원리(반송파 주파수 추정)
이 섹션은 어떠한 출처도 인용하지 않습니다.(2020년 1월 (이 및 에 대해 ) |
Hardy-Weinberg 공식( 2 + 2 p + 2 {\2}++q}=1}을 사용하여 헤테로 접합 상태(열성 특성의 운반체 상태)의 빈도를 추정할 수 있다.
이 공식은 정확히 두 개의 대립 유전자가 있는 유전자에 적용되며 많은 집단에서 이러한 대립 유전자의 빈도와 그 집단에서 세 개의 유전자형의 빈도를 관련짓습니다.
예를 들어 p가 대립 유전자 A의 빈도, q가 대립 유전자 a의 빈도라면 p, 2pq2, q는2 각각 유전자형 AA, Aa의 빈도이다.유전자는 단지 두 개의 대립 유전자를 가지고 있기 때문에, 모든 대립 유전자는 A 또는 a와 p + q = 1 중 하나여야 한다. 이제, A가 A에 대해 완전히 지배적인 경우, 운반 유전자형 Aa의 빈도는 직접적으로 관찰될 수 없다(호모 접합 유전자형 AA와 동일하기 때문에). 하지만, 모집단 내 열성 형질의 빈도로 추정할 수 있다.이는 호모 접합 유전자형 aa와 동일하기 때문에 개별 대립 유전자 빈도를 추정할 수 있다:f q = δ(aa), p = 1 - q, 이들로부터 운반 유전자형 빈도를 도출할 수 있다. (Aa) = 2pq
이 공식은 많은 가정과 열성 특성의 빈도에 대한 정확한 추정에 의존합니다.일반적으로 실제 상황은 이러한 가정으로부터 어느 정도 벗어나서 그에 상응하는 부정확성을 추정치에 도입한다.열성 특성이 드문 경우 매우 큰 샘플 크기가 필요하기 때문에 주파수를 정확하게 추정하기가 어렵습니다.
우위 대 우위
이 섹션은 어떠한 출처도 인용하지 않습니다.(2020년 1월 (이 및 에 대해 ) |
"지배적"의 속성은 때때로 유리하다는 개념과 혼동되고 "역행적"의 속성은 때때로 유해하다는 개념과 혼동되지만, 현상은 구별된다.우성은 서로 다른 표현형이 유익하거나 유해할 수 있는 정도에 관계없이 호모 접합자의 표현형에 관한 헤테로 접합자의 표현형을 기술한다.많은 유전병 대립 유전자가 열성이고 우세라는 단어가 긍정적인 의미를 내포하고 있기 때문에 지배적인 표현형이 적합성에 대해 우월하다는 가정이 종종 만들어진다.그러나 이것은 확실치 않다; 아래에 논의된 것처럼 대부분의 유전병 대립 유전자가 유해하고 열성인 반면, 모든 유전병이 열성인 것은 아니다.
그럼에도 불구하고, 이러한 혼란은 유전학의 역사를 통해 퍼져왔고 오늘날까지 지속되고 있다.이 혼란을 해결하는 것이 하디-바인버그 원칙을 출판하는 주된 동기 중 하나였다.
분자 메커니즘
지배력의 분자적 기초는 멘델에게 알려지지 않았다.유전자 궤적은 염색체의 특정 지점에 디옥시리보핵산(DNA)의 긴 염기 시리즈 또는 뉴클레오티드를 포함하고 있는 것으로 현재 이해되고 있다.분자생물학의 중심 교의는 "DNA가 RNA를 만들어 단백질을 만든다"고 말한다. 즉, DNA는 RNA 복사를 만들기 위해 전사되고 RNA는 단백질을 만들기 위해 번역된다.이 과정에서 궤적에서 다른 대립 유전자가 전사될 수도 있고 전사될 수도 있으며, 전사될 경우 동일한 단백질의 약간 다른 버전(isoforms라고 함)으로 번역될 수도 있다.단백질은 종종 직간접적으로 표현형을 생성하는 세포에서 화학 반응을 촉매하는 효소의 역할을 한다.게놈 내의 돌연변이는 촉매 활성을 바꿀 수 있고,[19] 따라서 지배력에 영향을 미칠 수 있다.어떤 이배체 유기체에서도 어떤 유전자 궤적에 존재하는 2개의 대립 유전자의 DNA 배열을 동일(호모 접합) 또는 다른(헤테로 접합)으로 할 수 있다.유전자 궤적이 DNA 배열 수준에서 헤테로 접합되어 있어도 각 대립 유전자에 의해 만들어지는 단백질은 동일할 수 있다.단백질 생성물 사이에 어떠한 차이도 없는 경우, 어느 대립 유전자도 우세하다고 말할 수 없다(위의 공우성 참조).두 단백질 생성물이 약간 다르더라도(알로자임), 효소 작용에 관해 동일한 표현형을 생성하기 쉬우며, 다시 말해 두 가지 대립 유전자가 지배적이라고 말할 수 없다.
접합성
역사적으로, 멘델의 독립적 분류의 법칙은 대립 유전자가 독립적으로 분류될 것이며, 하나의 대립 유전자가 "지배적"이라고 가정했다.유기체 대립 유전자의 유사성 정도인 접합성은 지배력에 영향을 미칠 수 있다.이배체 생물 내에서 이것들은 대립 유전자의 반수형 상호작용에 의해 정의될 것이다.유전자 반수체는 단일 기능성 대립 유전자가 호모[20] 접합체와 동일한 표현형을 생성하기에 충분한 단백질을 만드는 결과를 초래할 수 있다.일반적으로 다음 세 가지 유형의 하플로타입 상호작용이 가능합니다.
- 반만족.이배체에서는 반흡수 유전자의 기능적 대립 유전자가 우성으로 간주되는 반면 비기능적 대립 유전자는 열성 [21]유전자로 간주됩니다.예를 들어, 기능성 호모 접합체에서 생성되는 효소의 표준 양이 100%이며, 두 기능성 대립 유전자가 각각 50%를 기여한다고 가정합니다.헤테로 접합체의 단일 기능적 대립 유전자는 표준 효소의 50%를 생성하며, 이는 표준 표현형을 생성하기에 충분하다.헤테로 접합체와 기능알레 호모 접합체가 동일한 표현형을 가지면 기능성 대립 유전자가 비기능성 대립 유전자에 우세하다.이것은 알비노 유전자 궤적에서 발생한다: 헤테로 접합체는 색소 전구체를 멜라닌으로 전환하기에 충분한 효소를 생성하며, 개인은 표준 색소를 가지고 있다.예를 들어 인간과 다른 유기체에서 알비노 표현형의 색소화되지 않은 피부는 개인이 피부 색소 [22]멜라닌을 생성하는데 필요한 비기능적 버전의 효소를 코드하는 대립 유전자와 동질체일 때 발생한다.
- 불완전한 하플로우 기능.단일한 기능성 대립 유전자의 존재는 비기능성 호모 접합체보다 정상적이지 않지만 덜 심각한 표현형을 제공한다.이는 기능적 대립 유전자가 충분하지 않을 때 발생하며, 따라서 일반적으로 이러한 경우에 하플로 부족과 불완전한 우세라는 용어가 적용된다.중간 상호작용은 헤테로 접합 유전자형이 두 호모 접합체 사이의 중간 표현형을 생성하는 경우에 발생한다.헤테로 접합체 중 어느 것이 가장 닮았는지에 따라 하나의 대립 유전자가 다른 대립 유전자에 대해 불완전한 우위를 보이는 것으로 알려져 있다.예를 들어, 인간의 Hb 유전자 궤적은 혈액 색소 헤모글로빈을 구성하는 두 개의 글로빈 단백질 중 하나인 베타 사슬 단백질(HBB)을 담당합니다.많은 사람들이 Hb라고 불리는A 대립 유전자에 대해 동질감을 가지고 있다; 어떤 사람들은 Hb라고 불리는S 대체 대립 유전자를 가지고 있다, 호모 접합체 또는 헤테로 접합체.HbS/HbS 호모 접합체의 헤모글로빈 분자는 적혈구의 형태를 변형시키는 형태의 변화를 겪으며 겸상적혈구 빈혈이라고 불리는 심각하고 생명을 위협하는 형태의 빈혈을 일으킨다.이 대립 유전자에 대한 헤테로 접합 HbA/Hb는S 겸상세포 특성이라고 불리는 훨씬 덜 심각한 빈혈 형태를 가지고 있다.HbA/HbS 헤테로 접합자의 질병 표현형은 HbA/HbA 호모 접합체와 유사하지만 동일하지 않기 때문에 HbA 대립 유전자는 HbS 대립 [23]유전자에 불완전하게 우세한 것으로 알려져 있다.
- 완전한 하플로우 기능.헤테로 접합체 내의 단일 기능적 대립 유전자는 유전자의 어떤 기능에 대해서도 불충분한 유전자 생성물을 생성해 통상 기능적이지 않은 대립 유전자가 우세해지는 원인이 될 수 있다.그러면 표현형은 야생형 대신 비기능적 대립 유전자를 가진 호모 접합체와 유사할 것이다.비기능적 대립 유전자는 야생형 표현형의 기능적 대립 유전자에 지배적이라고 할 수 있다.이러한 상황은 비기능적 대립 유전자가 표준 대립 유전자에 의해 생성된 단백질의 적절한 기능을 방해하는 결함이 있는 단백질을 생성할 때 발생할 수 있습니다.결함이 있는 단백질의 존재는 표준 단백질을 "지배"하며, 헤테로 접합자의 질병 표현형은 두 개의 결함이 있는 대립 유전자에 대한 호모 접합자의 질병 표현형과 더 유사합니다."지배적"이라는 용어는 종종 동종 접합 표현형이 검사되지 않은 결함 대립 유전자에 잘못 적용되지만, 정상 대립 유전자와 헤테로 접합할 때 별개의 표현형을 야기한다.이 현상은 헌팅턴병을 예로 들 수 있는 많은 트리뉴클레오티드 반복 질환에서 발생한다.헌팅턴병에서는 완전한 하플로필이 돌연변이 단백질의 지배적인 효과를 일으킨다.보통 사람은 HTT 유전자에 약 20개의 C-A-G 뉴클레오티드 반복을 가지고 있지만 헌팅턴병을 앓고 있는 사람은 40개 이상의 C-A-G [24]반복을 가지고 있다.또 다른 예로는 피브릴린-1(FBN1) 유전자의 돌연변이에 의해 야기되는 유전성 결합조직 장애인 마르판 증후군이 있다.FBN1 유전자의 정상적인 복제는 한쪽 부모로부터 유전되며, 지배적인 비정상적인 FBN1 유전자는 다른 [25]한쪽 부모로부터 유전된다.
우성 음성 돌연변이
많은 단백질은 일반적으로 동일한 단백질의 여러 복사본의 집합체인 멀티머의 형태로 활성화되며, 호모몰리제릭 단백질 또는 호모올리제릭 단백질로 알려져 있다.사실, 브렌다 효소[26] 데이터베이스에 있는 9800개의 서로 다른 유기체의 83,000개의 서로 다른 효소들 중 대다수는 동질 [27]이성질체를 나타냅니다.돌연변이 버전과 함께 야생형 단백질이 존재할 경우 혼합 멀티머를 형성할 수 있다.멀티머 중 야생형 단백질의 활성을 교란시키는 돌연변이 단백질로 이어지는 돌연변이는 우성 음성 돌연변이이다.
인간 체세포에서 우성 음성 돌연변이가 일어나 돌연변이 세포에 증식상의 이점을 제공하여 복제세포의 팽창을 가져올 수 있다.예를 들어 DNA 손상에 대한 프로그램된 세포사망(Apoptosis)의 정상적인 과정에 필요한 유전자의 우성 음성 돌연변이는 세포를 아포토시스에 내성을 갖게 할 수 있다.이것은 과도한 DNA 손상이 있는 경우에도 클론의 증식을 가능하게 할 것이다.이러한 우성 음성 돌연변이는 종양 억제 유전자 p53에서 [28][29]발생한다.P53 야생형 단백질은 일반적으로 4단백질 멀티머(올리고테트라머)로 존재한다.우성 음성 p53 돌연변이는 다양한 유형의 암과 암 전 병변(예: 뇌종양, 유방암, 구강 전 암 병변 및 구강암)[28]에서 발생한다.
우성 음성 돌연변이는 다른 종양 억제 유전자에서도 발생한다.예를 들어 유방암에 [30]걸리기 쉬운 Ataxia telangioctasia mutiated(ATM) 유전자에서 2개의 우성 음성 생식선 돌연변이가 확인되었다.전사인자 C/EBPα의 지배적인 음성 돌연변이는 급성 골수성 [31]백혈병을 일으킬 수 있다.유전적인 우성 음성 돌연변이는 또한 암 이외의 질병의 위험을 증가시킬 수 있다.페르옥시좀 증식기 활성화 수용체 감마(PPAR))의 우성 음성 돌연변이는 심각한 인슐린 저항성, 당뇨병 및 [32]고혈압과 관련이 있다.
우성-음성 돌연변이는 인간 이외의 유기체에서도 설명되고 있다.실제로 혼합 멀티머에서 야생형 단백질의 정상 기능을 저해하는 돌연변이 단백질을 보고한 최초의 연구는 박테리오파지 T4 꼬리섬유 단백질 GP37이었다.[33]P53, ATM, C/EBPα 및 박테리오파지 T4 GP37의 연구에서 전장 돌연변이 단백질이 아닌 잘린 단백질을 생성하는 돌연변이는 지배-음성 효과가 가장 강한 것으로 보인다.
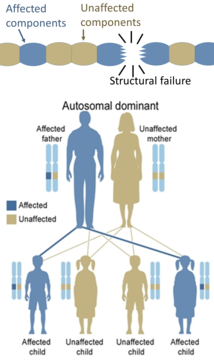
반면에, 구조 단백질의 유전적 결함(골형성 불완전증, 마르판 증후군, 엘러스-단로스 증후군 등)은 전체 구조를 기능하지 못하게 하기에 일부 성분이 결함이 있는 것으로 충분하기 때문에 일반적으로 상염색체 우성이다.이것은 변이된 유전자 생성물이 같은 세포 내에서 변이되지 않은 유전자 생성물에 악영향을 미치는 지배-음성 과정이다.


인간의 우성 및 열성 유전병
인간에게서 많은 유전적 특징이나 질병은 단순히 "지배적" 또는 "역행적"으로 분류된다.특히 소위 열성질환이라고 불리는 질환은 열성유전자의 한 요인이지만 기초분자기반을 지나치게 단순화시켜 우위의 본질에 대한 오해를 불러일으킬 수 있습니다.
예를 들어 열성유전병 페닐케톤뇨증(PKU)[34]은 효소 페닐알라닌수산화효소(PAH)[35]의 유전자 궤적에서 다수의 대립 유전자 중 하나(>60)에서 발생한다.이들 대립 유전자의 대부분은 PAH를 거의 생성하지 않으며, 그 결과 기질 페닐알라닌(Phe)과 그 대사 부산물이 중추신경계에 축적되어 치료되지 않으면 심각한 지적 장애를 일으킬 수 있다.
이러한 차이를 설명하기 위해, 세 가지 가설 PAH 대립 유전자 사이의 상호작용의 유전자형과 표현형 결과를 다음 [36]표에 나타냅니다.
| 유전자형 | PAH 활성 | [후] 콘센트 | PKU? |
|---|---|---|---|
| AA | 100% | 60μM | 아니요. |
| AB | 30% | 120μM | 아니요. |
| 참조 | 5% | 200 ~ 300 μM | 고페닐알라닌혈증 |
| BB | 0.3% | 600~2400μM | 네. |
비영향자는 표준기능알레알레아(AA)와 동형 접합하는 경우 PAH 활성은 표준(100%)이며, 혈중 페닐알라닌 농도는 약 60μM(=μmol/L)이다.1개의 PKU 대립 유전자(BB)에 대해 균질접합하는 치료되지 않은 사람에서는 PAH 활성은 0에 가깝고 [Phe]는 표준 10~40배이며 개인은 PKU를 나타낸다.
AB 헤테로 접합체는 PAH 활성도가 기준치의 30%(50%가 아님)에 불과하고, 혈액 [Phe]가 2배 상승하며, PKU가 나타나지 않는다.따라서, PKU에 관해서는 A 대립 유전자가 B 대립 유전자에 우세하지만, B 대립 유전자는 PAH 활성 수준(0.3% < 30% < < 100%)의 분자 효과와 관련하여 A 대립 유전자에 불완전하게 우세하다.마지막으로 A알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레알레열성 대립 유전자가 더 극단적인 [Phe] 표현형을 생성하는 것은 우성의 문제와 무관하다는 것을 다시 한 번 주목하십시오.
세 번째 대립 유전자 C의 경우, CC 호모 접합체는 매우 적은 양의 PAH 효소를 생성하며, 이는 혈중 [Phe]의 수치가 다소 높아지는데, 이는 고페닐알라닌혈증이라고 불리며 지적 장애를 초래하지 않는다.
즉, 두 대립 유전자의 지배 관계는 표현형의 어떤 측면이 고려되고 있는지에 따라 달라질 수 있다.유전자형과 관련된 대립 유전자 상호작용의 표현형 결과에 대해 이야기하는 것이 지배적이고 열성적인 범주로 강요하는 것보다 전형적으로 더 유용하다.
「 」를 참조해 주세요.
레퍼런스
- ^ "dominance". Oxford Dictionaries Online. Oxford University Press. Retrieved 14 May 2014.
- ^ "express". Oxford Dictionaries Online. Oxford University Press. Retrieved 14 May 2014.
- ^ Griffiths AJF; Gelbart WM; Miller JH; et al. (1999). "Gene Interaction Leads to Modified Dihybrid Ratios". Modern Genetic Analysis. New York: W. H. Freeman & Company. ISBN 978-0-7167-3118-4.
- ^ Mayo, O.와 Burger, R. 1997.우위의 진화: 시간이 흐른 이론?"생물학적 검토", 제72권, 제1호, 97-110페이지
- ^ Bourguet, D. 1999우성 유전의 진화, 제83, 제1권, 페이지 1~4
- ^ 바게리, H.C. 2006진화론의 해결되지 않은 경계와 유전체계가 어떻게 진화하느냐에 대한 질문: 지배력의 진화에 대한 75년간의 논쟁 "실험 동물학 저널 B: 분자와 발달 진화", 제306B권, 제4호, 페이지 329–359
- ^ "Human hg38 chr9:133,232,311-133,298,505 UCSC Genome Browser v433". genome.ucsc.edu. Retrieved 2022-07-10.
- ^ Ridley, Matt (1999). "Disease". Genome: The Autobiography of a Species in 23 Chapters. Harper Collins. pp. 136–146. ISBN 978-0-06-089408-5.
- ^ King, RC; et al. (2006). A Dictionary of Genetics (7th ed.). Oxford University Press. p. 129. ISBN 978-0-19-530761-0.
Dominance [refers] to alleles that fully manifest their phenotype when present in the heterozygous ... state.
- ^ Pennington, Sandra (1999). 11th Hour: Introduction to Genetics. Wiley. p. 43. ISBN 978-0-632-04438-2.
- ^ 우세하다.Genome.gov (n.d.)2022년 3월 29일 https://www.genome.gov/genetics-glossary/Dominant에서 취득
- ^ Allchin, D. (2005)지배의 딜레마.생물학 및 철학, 20(2-3), 427-451.doi:http://dx.doi.org/10.1007/s10539-005-2561-z
- ^ [1], 온라인 '쥐와 쥐의 유전자, 유전자 마커, 대립 유전자, 돌연변이 명명 가이드라인'
- ^ [2], 온라인 '옥수수 유전자 명명 표준'
- ^ "Cat Coat Color". Veterinary Genetics Laboratory, University of California. Archived from the original on 2012-05-05. Retrieved 2011-11-02.
- ^ Imes, D. L.; Geary, L. A.; Grahn, R. A.; Lyons, L. A. (April 2006). "Albinism in the domestic cat (Felis catus) is associated with a tyrosinase (TYR) mutation". Animal Genetics. 37 (2): 175–8. doi:10.1111/j.1365-2052.2005.01409.x. PMC 1464423. PMID 16573534.
- ^ Schmidt-Küntzel, A.; Eizirik, E.; O'Brien, S. J.; Menotti-Raymond, M. (April 2005). "Tyrosinase and tyrosinase related protein 1 alleles specify domestic cat coat color phenotypes of the albino and brown loci". Journal of Heredity. 96 (4): 289–301. doi:10.1093/jhered/esi066. PMID 15858157.
- ^ Carr, Steven M. "Extensions to Mendelian Analysis". Memorial University of Newfoundland.
- ^ Kacser, Henrik; Burns, James A (1981-03-01). "The Molecular Basis of Dominance". Genetics. 97 (3–4): 639–666. doi:10.1093/genetics/97.3-4.639. ISSN 1943-2631. PMC 1214416. PMID 7297851.
- ^ Encyclopedia of cancer. M. Schwab (2nd ed.). Berlin: Springer. 2009. ISBN 978-3-540-47648-1. OCLC 656366443.
{{cite book}}: CS1 유지보수: 기타 (링크) - ^ www.cancer.gov. 2012-07-20 https://www.cancer.gov/publications/dictionaries/genetics-dictionary/def/haploinsufficiency. Retrieved 2022-04-28.
{{cite web}}:누락 또는 비어 있음title=(도움말) - ^ Federico, Justin R.; Krishnamurthy, Karthik (April 19, 2022). "Albinism". StatPearls. StatPearls Publishing. PMID 30085560 – via PubMed.
- ^ Steinberg, M. H., & Sebastiani, P. (2012).겸상적혈구 질환의 유전적 수식어.미국 혈액학 저널, 87(8), 795~803.https://doi.org/10.1002/ajh.23232
- ^ 신, J. W., 김, K.-H., 차오, M. J., 앗월, R. S., 길리스, T., 맥도날드, M. E., 구셀라, J. F. & Lee, J.-M. (2016년 9월 15일).개인화된 대립 유전자 특이적 CRISPR/Cas9에 의한 헌팅턴병 돌연변이의 영구적 비활성화.OUP Academic.2022년 3월 4일 https://academic.oup.com/hmg/article/25/20/4566/2525895에서 취득
- ^ Nancy Garrick, Deputy Director (2017-04-11). "Marfan Syndrome". National Institute of Arthritis and Musculoskeletal and Skin Diseases. Retrieved 2022-03-18.
- ^ Schomburg I; Chang A; Ebeling C; et al. (January 2004). "BRENDA, the enzyme database: updates and major new developments". Nucleic Acids Res. 32 (Database issue): D431–3. doi:10.1093/nar/gkh081. PMC 308815. PMID 14681450.
- ^ Hashimoto K; Nishi H; Bryant S; Panchenko AR (June 2011). "Caught in self-interaction: evolutionary and functional mechanisms of protein homooligomerization". Phys Biol. 8 (3): 035007. Bibcode:2011PhBio...8c5007H. doi:10.1088/1478-3975/8/3/035007. PMC 3148176. PMID 21572178.
- ^ a b Marutani M; Tonoki H; Tada M; et al. (October 1999). "Dominant-negative mutations of the tumor suppressor p53 relating to early onset of glioblastoma multiforme". Cancer Res. 59 (19): 4765–9. PMID 10519380.
- ^ Goh AM; Coffill CR; Lane DP (January 2011). "The role of mutant p53 in human cancer". J. Pathol. 223 (2): 116–26. doi:10.1002/path.2784. PMID 21125670. S2CID 23998813.
- ^ Chenevix-Trench G; Spurdle AB; Gatei M; et al. (February 2002). "Dominant negative ATM mutations in breast cancer families". J. Natl. Cancer Inst. 94 (3): 205–15. doi:10.1093/jnci/94.3.205. PMID 11830610.
- ^ Pabst T; Mueller BU; Zhang P; et al. (March 2001). "Dominant-negative mutations of CEBPA, encoding CCAAT/enhancer binding protein-alpha (C/EBPalpha), in acute myeloid leukemia". Nat. Genet. 27 (3): 263–70. doi:10.1038/85820. PMID 11242107. S2CID 33788907.
- ^ Barroso I; Gurnell M; Crowley VE; et al. (1999). "Dominant negative mutations in human PPARgamma associated with severe insulin resistance, diabetes mellitus and hypertension". Nature. 402 (6764): 880–3. Bibcode:1999Natur.402..880B. doi:10.1038/47254. PMID 10622252. S2CID 4423555.
- ^ Bernstein H; Fisher KM (March 1968). "Dominance in bacteriophage T4D". Genetics. 58 (3): 307–18. doi:10.1093/genetics/58.3.307. PMC 1211863. PMID 5662621.
- ^ 온라인 Mendelian In Man (OMIM) : 고페닐알라닌혈증, 비PKU 마일드 - 261600
- ^ 온라인 멘델리아 인간유전: 페닐알라닌 히드록실화효소; PAH - 612349
- ^ Carr, Steven M. "One Gene, One Enzyme". Memorial University of Newfoundland.
- "생물학 2250 – 유전학의 원리 온라인 노트"뉴펀들랜드 메모리얼 대학교
- 온라인 Mendelian In Man (OMIM) : 헤모글로빈 - 베타궤적; HBB - 141900 - 겸상세포 빈혈
- 온라인 멘델리안 인간유전(OMIM) : ABO 글리코실전달효소 - 110300 - ABO 혈액형
외부 링크
- 온라인 멘델리안 인 맨 상속(OMIM)
- "헌팅턴병의 자가 염색체 우세"스탠포드 교육을 위한 헌팅턴병 지원 프로젝트