피라미드 세포
Pyramidal cell| 피라미드 세포 | |
|---|---|
 | |
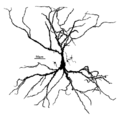
 피라미드형 세포의 재건. 소마와 덴드라이트는 붉은색으로, 액손 식물은 푸른색으로 표시한다. (1) 소마, (2) 바살 덴드라이트, (3) 아피칼 덴드라이트, (4) 액손, (5) 담보 액손. | |
| 세부 사항 | |
| 위치 | 대뇌피질 esp. 층 III 및 V |
| 모양 | 다극성 피라미드 |
| 함수 | 흥분 투영 뉴런 |
| 신경전달물질 | 글루탐산염, GABA |
| 식별자 | |
| 메슈 | D017966 |
| NeuroEx ID | 사오862606388 |
| TH | H1.00.01.0.00044 |
| FMA | 84105 |
| 신경조영술의 해부학적 용어 | |
피라미드 세포, 즉 피라미드 뉴런은 대뇌피질, 해마, 편도체를 포함한 뇌의 영역에서 발견되는 다극 뉴런의 일종이다. 피라미드 뉴런은 포유류 전두엽 피질 및 피질공막의 1차 흥분 단위다. 피라미드 뉴런은 또한 사후에 광견병 감염에서 특징적인 표식인 네그리 신체가 발견되는 두 가지 세포 유형 중 하나이다.[1] 피라미드 뉴런은 산티아고 라몬 이 카잘에 의해 처음 발견되고 연구되었다.[2][3] 그 이후로, 피라미드 뉴런에 대한 연구는 신경 플라스틱성에서 인지까지 이르는 주제에 초점을 맞추었다.
구조

녹색 형광 단백질로 시각화된 피라미드형 뉴런(gfp)
해마 피라미드 세포


피라미드형 뉴런의 주요 구조적 특징 중 하나는 원뿔 모양의 소마(soma) 즉 세포체인데, 그 뒤에 뉴런의 이름이 붙는다. 피라미드 셀의 다른 주요 구조적인 특징으로는 단일 축, 대형 아피칼 덴드라이트, 다중 기저 덴드라이트, 덴드리트리틱 가시의 존재 등이 있다.[4]
아피칼 덴드라이트
이 아피컬 덴드라이트는 피라미드 세포의 소마의 정점에서 솟아 있다. 아피칼 덴드라이트는 소마와의 거리가 증가하여 피질 표면을 향해 확장될수록 몇 배나 갈릴 수 있는 길고 두꺼운 덴드라이트다.[4]
바살 덴드라이트
염기성 덴드라이트는 소마 밑바닥에서 생긴다. 기초 덴드리트 나무는 3-5개의 기본 덴드라이트로 구성되어 있다. 소마로부터 거리가 늘어나면, 기초 덴드라이트가 많이 나온다.[4]
피라미드 세포는 뇌에서 가장 큰 뉴런 중 하나이다. 사람과 설치류 모두에서 피라미드 세포체(소마)는 평균 길이가 20μm 정도 된다. 피라미드 덴드라이트는 일반적으로 지름이 0.5마이크로미터에서 몇 마이크로미터까지 다양하다. 덴드라이트 한 개의 길이는 보통 수백 마이크로미터다. 분지로 인해, 피라미드 세포의 총 덴드리틱 길이는 몇 센티미터에 이를 수 있다. 피라미드 세포의 액손은 종종 더 길고 광범위하게 갈라져 총 길이가 수 센티미터에 이른다.
덴드리트리틱 가시
Dendritic spins는 피라미드 세포에 들어가는 대부분의 흥분성 자극(EPSP)을 받는다. 덴드리트리트의 가시나무는 1888년 라몬 이 카잘이 골기의 방법을 사용하여 처음 주목한 것이다. 또한 라몬 y 카잘은 뉴런의 수용성 표면적을 증가시키는 생리학적 역할을 처음으로 제안한 사람이었다. 피라미드 세포의 표면적이 클수록 뉴런의 대량의 정보를 처리하고 통합하는 능력이 커진다. 소마에는 덴드리틱 가시가 없는 반면, 소마에서는 숫자가 증가한다.[3] 쥐의 전형적인 아피셜 덴드라이트는 적어도 3,000개의 덴드리틱 가시를 가지고 있다. 평균적인 인간 아피알 덴드라이트는 쥐의 길이보다 약 두 배 정도 길기 때문에, 인간 아피알 덴드라이트에 존재하는 덴드리틱 가시의 수는 6,000개까지 될 수 있다.[5]
성장과 발전
차별화
피라미드 사양은 대뇌의 초기 발달 중에 발생한다. 생식기 세포는 부항 증식 심실 영역(VZ)과 심실 영역(SVZ)에서 신경 혈통에 헌신한다. 미성숙 피라미드 세포는 피질 판을 차지하기 위해 이동을 겪으며, 거기서 그들은 더욱 다양해진다. 엔도카나비노이드(eCBs)는 피라미드 세포 발달과 축 경로 찾기를 지시하는 것으로 보여진 분자의 한 종류다.[6] ctip2와 삭스5와 같은 전사 요인은 피라미드형 뉴런이 축을 지시하는 방향에 기여하는 것으로 나타났다.[7]
산후 조기발육
쥐의 피라미드 세포는 초기 산후 생활 동안 많은 빠른 변화를 겪는 것으로 나타났다. 산후 3일에서 21일 사이에, 피라미드 세포는 소마의 크기가 두 배가 되고, 아피칼 덴드라이트의 길이가 다섯 배 증가하며, 기저 덴드라이트 길이가 13배 증가하는 것으로 나타났다. 다른 변화로는 막의 휴면전위 감소, 막저항 감소, 작용전위의 피크값 증가 등이 있다.[8]
신호
대부분의 다른 뉴런의 덴드라이트와 마찬가지로 덴드라이트는 일반적으로 뉴런의 입력 영역인 반면, 액손은 뉴런의 출력 영역이다. 액손과 덴드라이트는 모두 브랜딩이 높다. 분기의 양이 많으면 뉴런은 많은 다른 뉴런들과 신호를 주고 받을 수 있다.
피라미드형 뉴런은 다른 뉴런들과 마찬가지로 수많은 전압 게이트 이온 채널을 가지고 있다. 피라미드 세포에는 덴드라이트에는 Na+, Ca2+, K+ 채널이 풍부하고, 소마에는 일부 채널이 있다. 피라미드 셀 덴드라이트 내의 이온 채널은 피라미드 셀 소마 내의 동일한 이온 채널 유형과 다른 특성을 가지고 있다. 피라미드 셀 덴드라이트의 전압 게이트 Ca2+ 채널은 하위 계위 EPSP와 역방향 제안 작용 전위에 의해 활성화된다. 피라미드 덴드라이트 내 행동 전위의 후방 제안 범위는 K+ 채널에 따라 달라진다. 피라미드 세포 덴드라이트의 K+ 채널은 작용 전위의 진폭을 제어하는 메커니즘을 제공한다.[9]
정보를 통합하는 피라미드형 뉴런의 능력은 그들이 받는 시냅스 입력의 수와 분포에 달려 있다. 단일 피라미드형 셀은 약 30,000개의 흥분 입력과 1700개의 억제(IPSP) 입력을 받는다. 흥분성(EPSP) 입력은 덴드리틱 척추에서만 종료되는 반면 억제성(IPSP) 입력은 덴드리틱 샤프트, 소모 및 액손에서도 종료된다. 피라미드 뉴런은 신경전달물질 글루탐산염에 의해 흥분될 수 있고 신경전달물질 GABA에 의해 억제될 수 있다.[4][10][4]
발화분류
피라미드 뉴런은 400-1000밀리초의 전류 펄스에 대한 발화 반응에 기초하여 다른 하위 분류로 분류되었다. 이러한 분류는 RSad, RSNA, IB 뉴런이다.
RSad
RSad pyramidal nerrones, 즉 규칙적으로 스파이크하는 뉴런을 적응시키는 것은 개별적인 행동 전위(AP)로 불을 지피고, 그 뒤에 극지방화 후 전위(potolarizing potential)가 뒤따른다. 애프터 전위는 뉴런에서 스파이크 주파수 적응(SFA)을 생성하는 지속시간이 증가한다.[11]
RSNA
RSNA 피라미드형 뉴런 또는 비접착형 정규 스파이크 뉴런은 맥박이 끝난 후 일련의 행동 전위를 발사한다. 이 뉴런들은 적응할 기미가 보이지 않는다.[11]
IB
IB 피라미드형 뉴런 또는 본질적으로 폭발하는 뉴런은 2-5개의 빠른 작용 전위가 폭발하면서 임계 펄스에 반응한다. IB 피라미드형 뉴런은 적응이 안 된다.[11]
분자분류
형태학적 및 전기적 피라미드적 세포 특성이 단일 세포 염기서열에 의해 측정된 유전자 발현으로부터 추론될 수 있다는 것을 보여주는 여러 연구가 있다.[12] 몇몇 연구들은 유전자 발현에 기초하여 쥐와[13][14] 인간의 뉴런에 단일 세포 분류를 제안하고 있다. 이러한 분류에서 뉴런 타입은 흥분성, 억제성, 그리고 그에 상응하는 수백 개의 서브바이트로 나뉜다. 예를 들어 인간 내 2-3층의 피라미드형 세포는 FREM3형으로[12] 분류되며 HCN 채널에서 발생하는 Ih 전류의[15] 양이 많은 경우가 많다.
함수
코티코스피탈관
피라미드 뉴런은 코르티코스피탈 트랙에서 1차 신경 세포 유형이다. 정상적인 모터 제어는 코티코스페이스와 척수 사이의 연결부의 발달에 따라 달라진다. 피라미드 세포 액손은 특정한 연결을 만들기 위해 성장 요인 같은 단서들을 따른다. 적절한 연결을 통해 피라미드형 셀은 시력 유도 모터 기능을 담당하는 회로에 참여한다.[16]
인식
전두엽 피질의 피라미드형 뉴런은 인지능력에 관련되어 있다. 포유류에서 피라미드 세포의 복잡성은 후뇌에서 전뇌 영역으로 증가한다. 피라미드 뉴런의 복잡성의 정도는 다른 인류학적 종의 인지 능력과 연관되어 있을 가능성이 높다. 전두엽 피질 내의 피라미드형 세포는 1차 청각 피질, 1차 섬광 피질, 1차 시각 피질로부터의 입력을 처리하는 데 책임이 있는 것으로 보이며, 이 모든 것은 감각 양식을[citation needed] 처리한다. 이 세포들은 또한 피질의 시각적 처리 영역 내에서 복잡한 물체 인식에 중요한 역할을 할 수 있다.[2]
참고 항목
참조
- ^ Sketchy Group, LLC. "2.3 rhabdovirus". SketchyMedical. Archived from the original on 2017-04-13.
- ^ a b Elston GN (November 2003). "Cortex, cognition and the cell: new insights into the pyramidal neuron and prefrontal function". Cereb. Cortex. 13 (11): 1124–38. doi:10.1093/cercor/bhg093. PMID 14576205.
- ^ a b García-López P, García-Marín V, Freire M (November 2006). "Three-dimensional reconstruction and quantitative study of a pyramidal cell of a Cajal histological preparation". J. Neurosci. 26 (44): 11249–52. doi:10.1523/JNEUROSCI.3543-06.2006. PMC 6674523. PMID 17079652.
- ^ a b c d e Megías M, Emri Z, Freund TF, Gulyás AI (2001). "Total number and distribution of inhibitory and excitatory synapses on hippocampal CA1 pyramidal cells". Neuroscience. 102 (3): 527–40. doi:10.1016/S0306-4522(00)00496-6. PMID 11226691. S2CID 16458290.
- ^ Laberge D, Kasevich R (November 2007). "The apical dendrite theory of consciousness". Neural Netw. 20 (9): 1004–20. doi:10.1016/j.neunet.2007.09.006. PMID 17920812.
- ^ Mulder J, Aguado T, Keimpema E, et al. (June 2008). "Endocannabinoid signaling controls pyramidal cell specification and long-range axon patterning". Proc. Natl. Acad. Sci. U.S.A. 105 (25): 8760–5. Bibcode:2008PNAS..105.8760M. doi:10.1073/pnas.0803545105. PMC 2438381. PMID 18562289.
- ^ Fishell G, Hanashima C (February 2008). "Pyramidal neurons grow up and change their mind". Neuron. 57 (3): 333–8. doi:10.1016/j.neuron.2008.01.018. PMID 18255026. S2CID 15095100.
- ^ Zhang ZW (March 2004). "Maturation of layer V pyramidal neurons in the rat prefrontal cortex: intrinsic properties and synaptic function". J. Neurophysiol. 91 (3): 1171–82. doi:10.1152/jn.00855.2003. PMID 14602839.
- ^ Magee J, Hoffman D, Colbert C, Johnston D (1998). "Electrical and calcium signaling in dendrites of hippocampal pyramidal neurons". Annu. Rev. Physiol. 60 (1): 327–46. doi:10.1146/annurev.physiol.60.1.327. PMID 9558467.
- ^ Wong, R. K. S.; Traub, R. D. (2009-01-01), "NETWORKS Cellular Properties and Synaptic Connectivity of CA3 Pyramidal Cells: Mechanisms for Epileptic Synchronization and Epileptogenesis", in Schwartzkroin, Philip A. (ed.), Encyclopedia of Basic Epilepsy Research, Oxford: Academic Press, pp. 815–819, doi:10.1016/b978-012373961-2.00215-0, ISBN 978-0-12-373961-2, retrieved 2020-11-18
- ^ a b c Franceschetti S, Sancini G, Panzica F, Radici C, Avanzini G (April 1998). "Postnatal differentiation of firing properties and morphological characteristics in layer V pyramidal neurons of the sensorimotor cortex". Neuroscience. 83 (4): 1013–24. doi:10.1016/S0306-4522(97)00463-6. PMID 9502243. S2CID 6986307.
- ^ a b Berg, Jim; Sorensen, Staci A.; Ting, Jonathan T.; Miller, Jeremy A.; Chartrand, Thomas; Buchin, Anatoly; Bakken, Trygve E.; Budzillo, Agata; Dee, Nick; Ding, Song-Lin; Gouwens, Nathan W. (October 2021). "Human neocortical expansion involves glutamatergic neuron diversification". Nature. 598 (7879): 151–158. doi:10.1038/s41586-021-03813-8. ISSN 1476-4687. PMID 34616067.
- ^ Gouwens, Nathan W.; Sorensen, Staci A.; Berg, Jim; Lee, Changkyu; Jarsky, Tim; Ting, Jonathan; Sunkin, Susan M.; Feng, David; Anastassiou, Costas A.; Barkan, Eliza; Bickley, Kris (July 2019). "Classification of electrophysiological and morphological neuron types in the mouse visual cortex". Nature Neuroscience. 22 (7): 1182–1195. doi:10.1038/s41593-019-0417-0. ISSN 1546-1726. PMC 8078853.
- ^ Bakken, Trygve E.; Jorstad, Nikolas L.; Hu, Qiwen; Lake, Blue B.; Tian, Wei; Kalmbach, Brian E.; Crow, Megan; Hodge, Rebecca D.; Krienen, Fenna M.; Sorensen, Staci A.; Eggermont, Jeroen (October 2021). "Comparative cellular analysis of motor cortex in human, marmoset and mouse". Nature. 598 (7879): 111–119. doi:10.1038/s41586-021-03465-8. ISSN 1476-4687.
- ^ Kalmbach, Brian E.; Buchin, Anatoly; Long, Brian; Close, Jennie; Nandi, Anirban; Miller, Jeremy A.; Bakken, Trygve E.; Hodge, Rebecca D.; Chong, Peter; de Frates, Rebecca; Dai, Kael (2018-12-05). "h-Channels Contribute to Divergent Intrinsic Membrane Properties of Supragranular Pyramidal Neurons in Human versus Mouse Cerebral Cortex". Neuron. 100 (5): 1194–1208.e5. doi:10.1016/j.neuron.2018.10.012. ISSN 0896-6273.
- ^ Salimi I, Friel KM, Martin JH (July 2008). "Pyramidal tract stimulation restores normal corticospinal tract connections and visuomotor skill after early postnatal motor cortex activity blockade". J. Neurosci. 28 (29): 7426–34. doi:10.1523/JNEUROSCI.1078-08.2008. PMC 2567132. PMID 18632946.



{kind=link}
{kind=link}