양성자 껍질
Protist shell
| 관련 시리즈의 일부 |
| 생물유민화 |
|---|
 |
많은 양성자들은 보통 실리카(유리)나 탄산칼슘(찰크)으로 만들어진 보호 껍질이나 시험을 가지고 있다.[2] 양성자들은 대부분 단세포적이고 현미경적이다. 그들의 껍질은 종종 질기고 광물화된 형태로 분해에 저항하며, 마이크로포실로서 원생의 죽음에서 살아남을 수 있다. 비록 양성자들이 전형적으로 매우 작지만, 그것들은 어디에나 있다. 그들의 수는 그들의 껍질이 해양 퇴적물의 형성과 원소와 영양소의 세계적인 순환에 큰 역할을 할 정도로 많다.
개요
세포 생명은 단세포 원핵생물(현대 박테리아와 고고학 포함)에서 유래한 것으로 보이며, 이후 더욱 복잡한 진핵생물로 진화했다. 진핵생물에는 식물, 동물, 곰팡이, "양생자"와 같은 유기체가 포함된다. 양성자들은 보통 단세포적이고 현미경적이다.
양성자라는 용어는 생물학적으로 유사한 유기체 집단을 지칭하기 위해 역사적으로 사용되었지만, 현대의 연구는 그것이 공통 조상의 모든 후손들을 포함하고 있지 않은 편협한 집단이라는 것을 보여주었다. 그러므로 그것은 하나의 판을 구성하지 않으며 현재 공식적인 과학적 용도에 있지 않다. 그럼에도 불구하고, 이 용어는 식물, 곰팡이, 동물로 분류될 수 없는 진핵생물을 지칭하기 위해 비공식적으로 계속 사용되고 있다.
대부분의 양성자들은 육안으로는 볼 수 없을 정도로 작다. 그것들은 현재 18필라로 조직된 매우 다양한 유기체들이지만 분류하기가 쉽지 않다.[3][4] 연구 결과, 해양, 심해용제, 하천 퇴적물 등에 높은 양성자 다양성이 존재한다는 사실이 밝혀졌으며, 이는 많은 수의 진핵 미생물 집단이 아직 발견되지 않았음을 시사한다.[5][6] 진핵생물로서, 원자는 미토콘드리아나 골기 몸체와 같은 오르가넬뿐만 아니라 적어도 하나의 핵을 세포 내에 가지고 있다. 많은 양성자들은 무성생식이지만 유사분열이나 분열을 통해 빠르게 번식할 수 있다. 다른 양성자들은 성적으로나 무성으로 생식할 수 있다.[7]
박테리아와 고대의 세포와는 대조적으로, 양성자와 다른 진핵생물의 세포는 고도로 조직화된다. 식물, 동물, 곰팡이는 대개 다세포로 되어 있고 전형적으로 거시적이다. 대부분의 양성자는 단세포적이고 현미경적이지만 예외도 있고, 일부 해양 양성자는 단세포도 미역처럼 현미경도 아니다.
실리콘 기반

규소는 규산염의 형태로 쉽게 구할 수 있지만, 직접 사용하는 유기체는 극히 적다. 규조류, 방사성 물질, 규조 해면류는 그들의 골격의 구조 재료로 생물 발생 실리카를 사용한다. 더 발전된 식물에서 실리카 식물석(오피탈 식물석)은 세포에서 발생하는 단단한 미세한 몸이다; 쌀을 포함한 몇몇 식물들은 성장을 위해 실리카를 필요로 한다.[8][9][10] 실리카는 일부 발전소에서 식물 세포벽 강도와 구조적 무결성을 향상시키는 것으로 나타났다.[11]
디아톰스
디아톰은 주로 단세포 조류로 알려진 약 10만 종을 포함하는 망울을 형성한다. 디아톰은 매년 지구에서 생산되는 산소의 약 20%를 생산하고,[12] 그들이 살고 있는 바다로부터 매년 67억 미터톤의 실리콘을 흡수하며,[13] 바다에서 발견되는 유기 물질의 거의 절반을 기여한다.
규조류는 좌절이라고 불리는 보호 실리카(유리) 껍질에 둘러싸여 있다. 이 좌절들 중 많은 것들이 아름답게 공들여지고 복잡한 구조는 종종 "바다의 보석"이라고 불릴 정도로 그렇게 되어 있다.[14] 각각의 좌절은 작은 구멍으로 덮인 두 개의 서로 맞물리는 부분으로 만들어지며, 이 두 부분으로 규조 원자가 영양분과 노폐물을 교환한다.[15] 죽은 규조류의 좌절은 수백만년에 걸쳐 반 마일이나 깊이를 쌓을 수 있는 해저로 떠내려간다.[16]
디아톰은 실리콘을 생물학적 실리카(BSIO2) 형태로 사용하며,[17] 실리콘 운반 단백질은 주로 이러한 보호 세포벽 구조를 구성하는 데 사용된다.[18] 실리콘은 규산이나 규산염과 같은 용해된 형태로 바다로 들어간다.[19] 규조류는 이러한 형태의 실리콘의 주요 사용자들 중 하나이기 때문에, 그것들은 바다 전체에 실리콘의 농도에 크게 기여한다. 실리콘은 얕은 깊이의 규조류 생산성으로 인해 바다에서 영양분 같은 프로파일을 형성하는데, 이는 상해의 실리콘 농도가 적고 심해에 실리콘 농도가 높다는 것을 의미한다.[19]
상해의 이원자 생산성은 저해상으로 수출되는 실리콘의 양에 기여한다.[20] 이원자 세포가 상층 대양에서 라이스될 때, 철, 아연, 실리콘과 같은 그들의 영양소는 해양 눈이라고 불리는 과정을 통해 하층 대양으로 보내진다. 해양의 눈은 용해된 유기물의 수직적 혼합에 의해 미립자 유기물이 아래로 전달되는 것을 포함한다.[21] 규소의 가용성은 규소 생산성에 있어 매우 중요한 것으로 보이며 규산이 규산이 이용 가능한 한, 규소는 심해에 있는 다른 중요한 영양소 농도를 제공한다.[22]
해안 지역에서, 규조류는 주요 식물성 플랑크톤 유기체로 작용하며, 생물 유발 실리카 생산에 크게 기여한다. 그러나, 외해에서, 규조류는 전세계 연간 규소 생산에서 감소된 역할을 한다. 북대서양과 북태평양 아열대성 광산의 다이아톰은 전 세계 연간 해양 실리카 생산량의 약 6%에 불과한 반면, 남양은 전 세계 해양 생물 유발 실리카의 약 3분의 1을 생산한다.[23] 남해는 극미량의 실리콘만 이 지역에서 운반되기 때문에 "생물 지질 화학적 분열"을 가지고 있다고 한다.[24]



디아톰은 식물성 플랑크톤의 가장 흔한 종류 중 하나이다.

그들의 보호 껍질(퍼즐)은 실리콘으로 만들어졌다.


그것들은 많은 모양과 크기로 나온다.
_Various_diatoms.jpg)



.jpg)
.jpg)

두 개의 반쪽이 겹치는 펜네이트 규산염의 규산

이원자 화석

해양 1차 생산량의 50%를 차지하는 10만 종 이상의 규조류가 있다.








디아톰 좌절은 1억년 이상 축적되어 전 세계에 나노와 마이크로 구조화된 실리콘 산화물의 풍부한 퇴적물을 이아톰 지구의 형태로 남겨두었다. 광합성 조류에 의한 나노와 미세구조화된 실리카의 발생에 대한 진화적 원인은 아직 명확하지 않다. 다만 2018년 나노구조 실리카가 자외선을 반사하면 녹조세포 내 DNA가 보호되는 것으로 나타났으며, 이는 유리 우리 형성의 진화적 원인일 수 있다.[28][29]


| 외부 영상 | |
|---|---|
라디올리언


방사선은 주로 실리카로 만들어지고 구멍으로 뚫린 정교한 구상 껍질로 둘러싸인 단세포 포식자 원자다. 그들의 이름은 라틴어로 "라디우스"라는 뜻에서 왔다. 그들은 구멍을 통해 몸의 일부를 확장하여 먹이를 잡는다. 규조류의 실리카 좌절과 마찬가지로 방사선의 껍질은 방사선이 죽으면 해저로 가라앉아 해양 침전물의 일부로 보존될 수 있다. 이 잔해들은 미세화석으로서 과거의 해양 상태에 대한 귀중한 정보를 제공한다.[32]

다이어텀처럼, 방사선 전문의는 여러 가지 형태로 나타난다.

또한 규조류와 마찬가지로, 방사선의 껍질은 보통 규산염으로 만들어진다.

.jpg)
.jpg)


구형 방사선 쉘의 절단 도식도

클로도코쿠스아비에티누스




.jpg)
| 외부 영상 | |
|---|---|
칼슘 기반
코콜리토포스
코콜리토포레스는 단세포 광합성을 위한 두 개의 플라겔로 이루어진 미세한 양성자들이다. 그들 대부분은 cocc스피어라고 불리는 껍질에 의해 보호된다. 코코스피어는 화려한 원형 판이나 코코리스라고 불리는 비늘로 덮여있다. 코코리츠는 탄산칼슘으로 만들어진다. coccolithophore라는 용어는 그리스어에서 돌을 나르는 씨앗으로, 그 작은 크기와 그들이 가지고 있는 coccolith 돌들을 가리킨다. 적절한 조건 하에서 그들은 다른 식물성 플랑크톤처럼 꽃을 피우며, 바다를 흰색으로 바꿀 수 있다.[36]


.png)


보호 포탄을 운반하는 양성자들에게는 이득이 있다. 아래 왼쪽에 있는 도표는 coccolithophore가 coccolith를 운반함으로써 얻는 몇 가지 이점을 보여준다. 도표에서 (A)는 탄소집중 메커니즘(CCM)을 포함한 가속 광합성 및 심층수양종의 희귀 광자 산란을 통한 광 흡수 강화 광합성을 나타낸다. (B)는 자외선(UV) 및 광합성 활성 방사선(PAR)으로부터 자외선 차단과 에너지 방해를 포함한 광자극으로부터의 보호를 나타낸다.(C)는 바이러스/박테리아 감염으로부터 보호되고 선택적 및 비선택적 방목자에 의한 방목과 방목을 포함하는 갑옷 보호 기능을 나타낸다.[38]


또한 보호 포탄을 운반하는 양성자의 비용도 있다. 오른쪽의 도표는 코코리스를 운반할 때 발생하는 에너지적인 비용의 일부를 보여준다. 도표에서 에너지 비용은 총 광합성 예산의 비율로 보고된다. (A) 운송 프로세스는 1차 석회화 기판인 Ca2+와 HCO-3(검은색 화살표)의 주변 바닷물로부터 세포로 운송하는 것과 최종 제품 H+를 세포에서 제거하는 것을 포함한다(회색 화살표). 세포질(coccolith vesicle, CV)을 통한 Ca2+의 운반은 석회화와 관련된 지배적인 비용이다. (B)는 CaCO3 결정의 핵화와 기하학을 조절하는 골기 복합체(흰색 직사각형)에 의한 코콜리트 관련 다당체(CAP - 회색 직사각형)의 합성을 대사 과정을 나타낸다. 완성된 coccolith(회색판)는 복잡하게 배열된 CAP와 CaCO3 결정의 복잡한 구조물이다. (C) 기계적 및 구조적 공정은 완성된 coccolith가 핵에 인접한 원래 위치에서 세포 주변으로 운반되어 c의 표면으로 전달되는 것으로서 분비를 설명한다.타원형의[38]
포아미니페란스


방사선과와 마찬가지로 포라미페란(짧은 포름)은 단세포 포식자 원생이며, 또한 구멍이 있는 포탄으로 보호된다. 그들의 이름은 "구멍 뚫는 사람"을 뜻하는 라틴어에서 유래되었다. 종종 시험이라고 불리는 그들의 껍질은 단일 챔버로 되어 있거나 다중 챔버로 되어 있을 수 있다; 다중 챔버로 된 포람은 자라면서 더 많은 챔버를 추가한다. 이것들 중 가장 유명한 것은 석회암이지만 테스트는 아라곤이트, 아글루티드 침전물 입자, 치톤 또는 실리카 (rarely)로 만들어질 수도 있다. 대부분의 포람은 구부러져 있지만 약 40종의 살아있는 종은 플랑크어다.[39] 그것들은 과학자들이 과거의 환경과 기후에 대해 많은 것을 유추할 수 있도록 해주는 잘 확립된 화석 기록으로 널리 연구되고 있다.[32] 일부 포미니페라들은 전체적으로 검사가 부족하다.[40]


| 외상상상 | |
|---|---|


나선형 포램의 방을 보여주는 단면

먹이를 잡기 위한 세밀한 엑토플라즘을 스트리밍하는 살아있는 암모니아 테피다

플랑크토닉 포람 그룹





기타
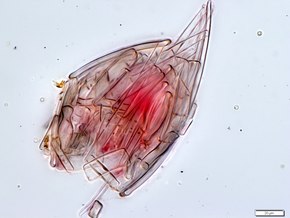
로리시페라(코르세레 또는 무장을 위한 라틴 로리카에서 유래)는 매우 작은 동물에서 미세한 해양 사이클론 퇴적물 처리 동물로 이루어진 망상이다. 이것의 크기는 약 100 μm에서 1 mm이다.[43][44] 그들은 로리카라고 불리는 보호용 외부 케이스에 의해 특징지어지고 그들의 서식지는 그들이 그들 자신을 붙이는 해양 자갈 사이의 공간에 있다. 갑옷과 같은 로리카는 보호용 외부 껍질 또는 포위 플리케 케이스로 구성된다.[45]
많은 choanoflagellate의 세포체는 구별되는 세포외 기질 또는 세포외 기질에 둘러싸여 있다. 이러한 세포 덮개는 구조와 구성이 크게 달라 분류학자들이 분류 목적으로 사용한다. 많은 choanoflagellates는 여러 실리카 조각들을 함께 접합한 것으로 로리카라 불리는 복잡한 바구니 모양의 "집"을 짓는다.[46] 경막의 기능적 중요성은 알려져 있지 않지만, sessile 유기체에서는 기질에 부착하는 것을 돕는다고 생각된다. 플랑크톤 유기체에서는 페리플라스트가 항력을 증가시켜 플라겔럼에 의해 생성되는 힘에 대항하고 먹이 효율을 증가시킨다는 추측이 있다.[46][47]
| 외상상상 | |
|---|---|



마이크로포실

마이크로포셀은 일반적으로 0.001mm에서 1mm 사이인 화석으로,[48] 연구를 통해 빛이나 전자현미경을 사용해야 한다. 손 렌즈와 같이 육안으로 연구할 수 있는 화석이나 저전력 확대 등을 매크로포실이라고 한다.
예를 들어, 브라이조아(특히 제일모스토마타)와 같은 일부 식민지 유기체는 비교적 큰 군락을 가지고 있지만, 군락의 소개인의 미세한 골격 세부사항으로 분류된다.
또 다른 예로, 원자인 포라미니페라의 많은 화석생물은 누물속과 같이 동전만큼 큰 껍질("시험"이라고 한다)에서 알려져 있다.
마이크로포셀은 프리캄브리아에서 홀로세까지 지질학적 기록의 공통적인 특징이다. 그것들은 해양 환경의 퇴적물에서 가장 흔하지만, 또한 고사리 물, 민물, 육지 퇴적물에서도 발생한다. 모든 생명의 왕국이 마이크로포실 기록에 나타나 있지만, 가장 풍부한 형태는 크리소피타, 피러호피타, 사코디나, 아세리타치, 치티노조아스의 원생 해골이나 낭종이며, 혈관 식물에서 나온 꽃가루와 포자와 함께 나타난다.
2017년 캐나다 퀘벡 누브와기투크 벨트(Nuvvuagittuq Belt)에서 화석화된 미생물, 즉 마이크로포실(microfossil)이 발견됐다고 발표돼 지구상 생명체의 가장 오래된 기록인 42억8000만년 전(지질적 시간적 의미로는) 이후 '생물의 거의 순간 출현'을 시사했다.ean 형성은 44억 1천만 년 전, 그리고 45억 4천만 년 전 지구의 형성이 있은 지 얼마 되지 않았다.[49][50][51][52] 그럼에도 불구하고, 일부 연구자들의 주장대로, 생명은 거의 45억년 전에 훨씬 더 일찍 시작되었을지도 모른다.[53][54]
참고 항목
참조
- ^ 뉴스의 가파르, 에어, B.D.와 K.G.의 슐츠(Schulz)는 "2019년 해양 식물성 플랑크톤 석회화에서 빛과 탄산염 화학의 변화에 대한 종별 민감도의 비교"라고 말했다. 자연: 과학 보고서, 9(1): 1–12. doi:10.1038/s41598-019-38661-0.
 자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다.
자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다. - ^ "Groups of Protists Boundless Biology". courses.lumenlearning.com. Retrieved 16 February 2021.
- ^ Cavalier-Smith T (December 1993). "Kingdom protozoa and its 18 phyla". Microbiological Reviews. 57 (4): 953–94. doi:10.1128/mmbr.57.4.953-994.1993. PMC 372943. PMID 8302218.
- ^ Corliss JO (1992). "Should there be a separate code of nomenclature for the protists?". BioSystems. 28 (1–3): 1–14. doi:10.1016/0303-2647(92)90003-H. PMID 1292654.
- ^ Slapeta J, Moreira D, López-García P (2005). "The extent of protist diversity: insights from molecular ecology of freshwater eukaryotes". Proceedings of the Royal Society B: Biological Sciences. 272 (1576): 2073–81. doi:10.1098/rspb.2005.3195. PMC 1559898. PMID 16191619.
- ^ Moreira D, López-García P (2002). "The molecular ecology of microbial eukaryotes unveils a hidden world" (PDF). Trends in Microbiology. 10 (1): 31–8. doi:10.1016/S0966-842X(01)02257-0. PMID 11755083.
- ^ Foraminifera : notes for a short course organized by M.A. Buzas and B.K. Sen Gupta : prepared for the short course on foraminifera sponsored by the Paleontological Society, held at New Orleans, Louisiana, October 17, 1982. Thomas W. Broadhead, Paleontological Society. [Knoxville, Tenn.]: University of Tennessee, Dept. of Geological Sciences. 1982. ISBN 0-910249-05-9. OCLC 9276403.
{{cite book}}: CS1 maint : 기타(링크) - ^ Rahman, Atta-ur- (2008). "Silicon". Studies in Natural Products Chemistry. Vol. 35. p. 856. ISBN 978-0-444-53181-0.
- ^ Exley, C. (1998). "Silicon in life:A bioinorganic solution to bioorganic essentiality". Journal of Inorganic Biochemistry. 69 (3): 139–144. doi:10.1016/S0162-0134(97)10010-1.
- ^ Epstein, Emanuel (1999). "SILICON". Annual Review of Plant Physiology and Plant Molecular Biology. 50: 641–664. doi:10.1146/annurev.arplant.50.1.641. PMID 15012222.
- ^ Kim, Sang Gyu; Kim, Ki Woo; Park, Eun Woo; Choi, Doil (2002). "Silicon-Induced Cell Wall Fortification of Rice Leaves: A Possible Cellular Mechanism of Enhanced Host Resistance to Blast". Phytopathology. 92 (10): 1095–103. doi:10.1094/PHYTO.2002.92.10.1095. PMID 18944220.
- ^ 숨쉬는 공기? 그것을 만든 디아톰
- ^ Treguer, P.; Nelson, D. M.; Van Bennekom, A. J.; Demaster, D. J.; Leynaert, A.; Queguiner, B. (1995). "The Silica Balance in the World Ocean: A Reestimate". Science. 268 (5209): 375–9. Bibcode:1995Sci...268..375T. doi:10.1126/science.268.5209.375. PMID 17746543. S2CID 5672525.
- ^ 아일랜드, T, "조류 공학" 생물학자, 63(5): 10.
- ^ 와실리프, 매기(2006) "플랑크톤 - 식물 플랑크톤", 티아라 - 뉴질랜드 백과사전 접속: 2019년 11월 2일.
- ^ "King's College London - Lake Megachad". www.kcl.ac.uk. Retrieved 5 May 2018.
- ^ Bidle, Kay D.; Manganelli, Maura; Azam, Farooq (6 December 2002). "Regulation of Oceanic Silicon and Carbon Preservation by Temperature Control on Bacteria". Science. 298 (5600): 1980–1984. Bibcode:2002Sci...298.1980B. doi:10.1126/science.1076076. ISSN 0036-8075. PMID 12471255. S2CID 216994.
- ^ Durkin, Colleen A.; Koester, Julie A.; Bender, Sara J.; Armbrust, E. Virginia (2016). "The evolution of silicon transporters in diatoms". Journal of Phycology. 52 (5): 716–731. doi:10.1111/jpy.12441. ISSN 1529-8817. PMC 5129515. PMID 27335204.
- ^ a b Dugdale, R. C.; Wilkerson, F. P. (30 December 2001). "Sources and fates of silicon in the ocean: the role of diatoms in the climate and glacial cycles". Scientia Marina. 65 (S2): 141–152. doi:10.3989/scimar.2001.65s2141. ISSN 1886-8134.
- ^ Baines, Stephen B.; Twining, Benjamin S.; Brzezinski, Mark A.; Krause, Jeffrey W.; Vogt, Stefan; Assael, Dylan; McDaniel, Hannah (December 2012). "Significant silicon accumulation by marine picocyanobacteria". Nature Geoscience. 5 (12): 886–891. Bibcode:2012NatGe...5..886B. doi:10.1038/ngeo1641. ISSN 1752-0908.
- ^ Turner, Jefferson T. (January 2015). "Zooplankton fecal pellets, marine snow, phytodetritus and the ocean's biological pump". Progress in Oceanography. 130: 205–248. Bibcode:2015PrOce.130..205T. doi:10.1016/j.pocean.2014.08.005. ISSN 0079-6611.
- ^ Yool, Andrew; Tyrrell, Toby (2003). "Role of diatoms in regulating the ocean's silicon cycle". Global Biogeochemical Cycles. 17 (4): n/a. Bibcode:2003GBioC..17.1103Y. doi:10.1029/2002GB002018. ISSN 1944-9224.
- ^ Tréguer, Paul J.; De La Rocha, Christina L. (3 January 2013). "The World Ocean Silica Cycle". Annual Review of Marine Science. 5 (1): 477–501. doi:10.1146/annurev-marine-121211-172346. PMID 22809182.
- ^ Marinov, I.; Gnanadesikan, A.; Toggweiler, J. R.; Sarmiento, J. L. (June 2006). "The Southern Ocean biogeochemical divide". Nature. 441 (7096): 964–967. Bibcode:2006Natur.441..964M. doi:10.1038/nature04883. PMID 16791191. S2CID 4428683.
- ^ Arsenieff, L.; Simon, N.; Rigaut-Jalabert, F.; Le Gall, F.; Chaffron, S.; Corre, E.; Com, E.; Bigeard, E.; Baudoux, A.C. (2018). "First Viruses Infecting the Marine Diatom Guinardia delicatula". Frontiers in Microbiology. 9: 3235. doi:10.3389/fmicb.2018.03235. PMC 6334475. PMID 30687251.
- ^ Zhang, D.; Wang, Y.; Cai, J.; Pan, J.; Jiang, X.; Jiang, Y. (2012). "Bio-manufacturing technology based on diatom micro- and nanostructure". Chinese Science Bulletin. 57 (30): 3836–3849. Bibcode:2012ChSBu..57.3836Z. doi:10.1007/s11434-012-5410-x.
- ^ 생분해성 반짝이와 공해를 먹는 미세조류: 자연 Horizon, 2020년 5월 28일 영감을 받은 신소재.
- ^ a b 아귀레, L.E., 오우양, L., 엘프윙, A, 헤드블럼, M, 울프, A., O. (2018) "다이아톰 좌절은 자외선으로부터 DNA를 보호한다. 과학 보고서, 8(1): 1–6. doi:10.1038/s41598-018-21810-2. 자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다.
- ^ De Tomasi, E, Bulfri, R, Dardano, P, De Luca, A.C, Managa, S, Rea, I., De Stefano, M. (2018) "중심적 규조 나노파트에 의한 UV 차폐 및 파장 변환" 자연: 과학 보고서, 8(1): 1–14. doi:10.1038/s41598-018-34651-w. 자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다.
- ^ B.C.의 부스와 H.J.의 Marchant(1987) "파르말레스, 해양 번데기의 새로운 순서, 3개의 새로운 제네라와 7개의 새로운 종에 대한 설명이 곁들여져 있다." 식물학 저널, 23:245–260. doi:10.111/j.1529-8817.1987.tb04132.x.
- ^ 쿠와타, A, 야마다, K, 이치노미야, M, 요시카와, S, 트라긴, M, 바울롯, D., 로페스 도스 산토스, A. (2018) "보리도피콜롬의 자매 피코플랑크톤 그룹인 볼리도피체 - 검토" 해양 과학 분야의 프론티어, 5: 370. doi:10.3389/fmars.2018.00370. 자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다.
- ^ a b 와실리프, 매기(2006) "플랑크톤 - 동물 플랑크톤", 티아라 - 뉴질랜드 백과사전 접속: 2019년 11월 2일.
- ^ 카초비치, 사라 (2018) "방법에 대한 마인드: 마이크로포셀과 텍토닉스를 연결" 유럽지질과학연합의 구조지질학부 블로그
- ^ 카초비치, S, 현, J.C., 에이치슨, 2019. 초기 방사선 진화 연구에 새로운 차원을 추가한다. 과학 보고서, 9(1), 페이지.1-10. doi:10.1038/s41598-019-42771-0. 자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다.
- ^ Varea, C.; Aragon, J.L.; Barrio, R.A. (1999). "Turing patterns on a sphere". Physical Review E. 60 (4): 4588–92. Bibcode:1999PhRvE..60.4588V. doi:10.1103/PhysRevE.60.4588. PMID 11970318.
- ^ Washsilieff, Maggy(2006) "A coccolithophore", Te Ara - 뉴질랜드 백과사전 접속: 2019년 11월 2일.
- ^ 로스트, B, 리에베셀, U.S. (2004) "코콜리토포레스와 생물 펌프: 환경 변화에 대한 대응" 인: 코콜리토포레스: 분자 프로세스에서 글로벌 임팩트에 이르기까지, 99-125페이지의 스프링어. ISBN 9783662062784.
- ^ a b c d Monteiro, F.M., Bach, L.T., Brownlee, C., Bown, P., Rickaby, R.E., Poulton, A.J., Tyrrell, T., Beaufort, L., Dutkiewicz, S., Gibbs, S. and Gutowska, M.A. (2016) "Why marine phytoplankton calcify". Science Advance, 2(7): e1501822. doi:10.1126/sciadv.1501822. 자료는 이 출처에서 복사되었으며, Creative Commons Accountation 4.0 International License에 따라 이용할 수 있다.
- ^ Hemleben, C.; Anderson, O.R.; Spindler, M. (1989). Modern Planktonic Foraminifera. Springer-Verlag. ISBN 978-3-540-96815-3.
- ^ Pawlowski, Jan; Bolivar, Ignacio; Fahrni, Jose F.; Vargas, Colomban De; Bowser, Samuel S. (November 1999). "Molecular Evidence That Reticulomyxa Filosa Is A Freshwater Naked Foraminifer". The Journal of Eukaryotic Microbiology. 46 (6): 612–617. doi:10.1111/j.1550-7408.1999.tb05137.x. ISSN 1066-5234.
- ^ Favulina 육각형 유럽 지질학 연합, 2020년 11월 9일.
- ^ 포라미니페라: 런던 대학의 공부 역사. 검색됨: 2019년 11월 18일.
- ^ Gad, Gunnar (17 June 2005). "Successive reduction of the last instar larva of Loricifera, as evidenced by two new species of Pliciloricus from the Great Meteor Seamount (Atlantic Ocean)". Zoologischer Anzeiger. 243 (4): 239–71. doi:10.1016/j.jcz.2004.09.001.
- ^ Heiner, Iben. "Preliminary account of the Loriciferan fauna of the Faroe Bank (NE Atlantic)". Annales Societatis Scientiatum Færoensis Supplementum. 41: 213–9. Archived from the original on 27 December 2016. Retrieved 26 December 2016.
- ^ Heiner, Iben; Sorensen, Martin Vinther; Kristensen, Reinhardt Mobjerg (2004). Loricifera (Girdle Wearers) in Grzimek's Animal Life Encyclopedia Volume 1. New York, NY: Van Nostrand Reinhold Company. pp. 343–350.
- ^ a b Leadbeater BS, Thomsen H (2000). "Order Choanoflagellida". An Illustrated Guide to the Protozoa, Second Edition. Lawrence : Society of Protozoologists. 451: 14–38.
- ^ Leadbeater BS, Kelly M (2001). "Evolution of animals choanoflagellates and sponges". Water and Atmosphere Online. 9 (2): 9–11.
- ^ Drewes, Charlie. "Discovering Devonian Microfossils". Iowa State University. Retrieved 4 March 2017.
- ^ Dodd, Matthew S.; Papineau, Dominic; Grenne, Tor; slack, John F.; Rittner, Martin; Pirajno, Franco; O'Neil, Jonathan; Little, Crispin T. S. (2 March 2017). "Evidence for early life in Earth's oldest hydrothermal vent precipitates" (PDF). Nature. 543 (7643): 60–64. Bibcode:2017Natur.543...60D. doi:10.1038/nature21377. PMID 28252057.
- ^ Zimmer, Carl (1 March 2017). "Scientists Say Canadian Bacteria Fossils May Be Earth's Oldest". The New York Times. Retrieved 2 March 2017.
- ^ Ghosh, Pallab (1 March 2017). "Earliest evidence of life on Earth 'found". BBC News. Retrieved 2 March 2017.
- ^ Dunham, Will (1 March 2017). "Canadian bacteria-like fossils called oldest evidence of life". Reuters. Retrieved 1 March 2017.
- ^ Staff (20 August 2018). "A timescale for the origin and evolution of all of life on Earth". Phys.org. Retrieved 20 August 2018.
- ^ Betts, Holly C.; Putick, Mark N.; Clark, James W.; Williams, Tom A.; Donoghue, Philip C.J.; Pisani, Davide (20 August 2018). "Integrated genomic and fossil evidence illuminates life's early evolution and eukaryote origin". Nature. 2 (10): 1556–1562. doi:10.1038/s41559-018-0644-x. PMC 6152910. PMID 30127539.
추가 참조사항
- 쉬, 케이, 허친스, 디, 가오(2018년)는 "코콜리토포레 에밀리아 헉슬리에서는 코콜리스 배열이 오일리아 수학을 따른다"고 말했다. PeerJ, 6: e4608. doi:10.1126/science.aa7378.
- 프로토스탄 스켈레톤: 진화와 제약의 지질학적 역사

