꽃망울
Inflorescence| 무료 사전인 Wiktionary에서 꽃무늬를 찾아 보십시오. |

꽃차례는 줄기에 배열된 꽃차례로 주가지나 가지들의 복잡한 배열로 이루어져 있다.[1] 형태학적으로 꽃이 형성되는 종자식물 촬영의 변형된 부분이다.수정은 내막과 필로트축의 길이와 성질, 그리고 주축과 보조축의 비율, 압축, 팽창, 어음, 결절 및 축소의 변화를 수반할 수 있다.또한 꽃 군락을 특정 패턴으로 품는 식물의 생식 부분이라고 정의할 수 있다.
전체 염기를 잡고 있는 줄기를 족발이라고 한다.꽃이나 이차 가지가 있는 발판 위의 주축(주줄기로 잘못 칭함)을 라치스라고 한다.꽃차례의 꽃줄기를 꽃차례의 꽃줄기로 페디셀이라고 한다.꽃의 일부가 아닌 꽃을 고독한 꽃이라 하고 그 줄기는 족발이라고도 한다.특히 개별 꽃들이 특히 작고 pseudanthium과 같이 빽빽한 군집 안에서 자라는 경우, 꽃의 꽃들은 플로레트라고 불릴 수 있다.꽃의 결실을 맺는 단계는 과격으로 알려져 있다.염증은 단순(단일) 또는 복잡(패니클)일 수 있다.라치스는 단일, 복합, 탯줄, 스파이크 또는 레이스미(레이스미)를 포함한 여러 가지 유형 중 하나일 수 있다.
일반적 특성
꽃병은 꽃들이 어떻게 발판 위에 배열되어 있는지, 꽃들의 개화 순서, 그리고 꽃들의 다른 군집들이 어떻게 그 안에 그룹화되는지 등 많은 다른 특징들로 설명된다.이 용어는 자연 속의 식물이 여러 종류의 조합을 가질 수 있기 때문에 일반적인 표현이다.이러한 구조 유형은 주로 자연 선택에 기초한다.[2]
브랙츠
꽃병은 보통 식물의 식물성 부분과는 다른 잎을 변형시켰다.이 용어의 가장 넓은 의미를 고려할 때, 꽃가루와 관련된 잎은 브랙트라고 불린다.Bract는 보통 식물의 라치스와 결합되는 염기의 주 줄기가 형성되는 노드에 위치하지만, 다른 염기 그 자체 내에 다른 염기가 존재할 수 있다.그들은 꽃가루 매개자들을 끌어들이고 어린 꽃들을 보호하는 것을 포함한 다양한 기능들을 제공한다.거래의 유무와 그 특성에 따라 우리는 다음을 구별할 수 있다.
- 감산염 인플레 현상:꽃가루에 제약을 넣지 않는다.
- Bracteate 염증:염기에 있는 bract는 매우 전문화되어 있으며, 때로는 작은 규모로 축소되어 분할되거나 해부된다.
- 잎이 많은 꽃가루:크기도 많이 줄었지만, 브랙트는 특수하지 않고 식물의 전형적인 잎처럼 보이기 때문에 꽃 줄기가 부풀어 오르는 대신 일반적으로 사용된다.이러한 사용은 기술적으로 정확하지 않다. 왜냐하면, '정상적인' 외관에도 불구하고, 사실상 이러한 잎은 브랙티스로 간주되기 때문에 '잎 부풀리기'가 더 바람직하다.
- 잎이 많은 염증:브락테이트와 잎이 무성한 꽃 사이의 중간.
만약 아스테르과에서처럼 많은 약물이 존재하고 줄기에 엄격히 연결되어 있다면, 약물은 집단적으로 비자발적 약자라고 불릴 수 있다.만약 염증이 두 번째 단위가 줄기 위로 더 올라가면, 그것들은 비자발적이라 불릴지도 모른다.

감산염색소.

위스테리아 시넨시스 에블랙테이트 인플레시

브락테이트가 부풀어 오른다.

페디컬리스 verticillata의 Bracteate inflorex.

잎이 무성한 꽃가루.

리난투스 앵거스티폴리오스의 잎이 돋은 꽃가루.

잎이 무성한 꽃.

아리스톨로키아 구개염의 잎이 돋는 것.
.svg)

.svg)

.svg)

.svg)

말단꽃
식물 기관은 두 가지 다른 계략에 따라 자랄 수 있다. 단면체 또는 경주체, 동정체 또는 시모스.염증에서는 이 두 가지 서로 다른 성장 패턴을 각각 미확정과 결정이라고 하며, 말기 꽃이 형성되는지 여부와 염증 내에서 개화가 시작되는 지점을 나타낸다.
- 불확실한 팽창:단면성(레이스모스) 성장.말단 꽃봉오리가 계속 자라 가로 꽃을 피운다.말단꽃은 결코 맺어지지 않는다.
- 팽창 결정:동정적(사이모스) 성장.말단봉오리는 말단꽃을 형성한 뒤 소멸한다.그리고 나서 다른 꽃들은 측면의 꽃봉오리에서 자란다.
불확정성 및 결정성 염증을 각각 개방성 및 폐쇄성 염증이라고 부르기도 한다.꽃의 불확실한 무늬는 결정되는 꽃에서 유래한다.미확정 꽃은 말기 꽃 성장을 막는 공통의 메커니즘을 가지고 있다고 제안한다.계통생리학적 분석에 기초하여, 이 메커니즘은 다른 종에서 독립적으로 여러 번 발생하였다.[3]
불확실한 꽃망울에는 진정한 말단 꽃이 없고 줄기는 보통 초보적인 끝을 가지고 있다.말단 꽃봉오리에 의해 형성된 마지막 참꽃(말단꽃)이 곧게 펴지면서 말단꽃으로 보이는 경우가 많다.종종 말단 봉오리의 흔적은 줄기에서 더 높게 발견될 수 있다.

완벽한 아크로페탈 성숙과 함께 불규칙적인 팽창.

아크로페탈 성숙과 측면 꽃봉오리로 불규칙적인 꽃망울을 만들어라.

단자 1을 시뮬레이션하기 위해 단자 이하의 꽃으로 불규칙한 꽃(베스티지가 있음)
염증을 결정함에 있어 말기 꽃은 보통 가장 먼저 성숙하는 (전발 발달) 반면, 다른 꽃들은 줄기의 밑부분부터 성숙하는 경향이 있다.이 패턴을 아크로페탈 성숙이라고 한다.꽃이 줄기의 꼭대기에서 성숙하기 시작할 때 성숙은 기저 태아인 반면, 중앙이 먼저 성숙하면 성숙은 다양하다.

아크로페탈 성숙을 통한 팽창성 결정

기저귀 성숙을 통한 염기성

다양한 성숙도로 팽창성 결정
필로타축
잎과 마찬가지로 꽃은 줄기에 여러 가지 무늬에 따라 배열할 수 있다.자세한 설명은 'Phyllotaxis'를 참조하십시오.
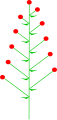
대체꽃

맞은편 꽃

.svg)
마찬가지로 잎의 싹 배열을 Ptyxis라고 한다.
단일 또는 한 무리의 꽃 군집이 브라트의 축에 위치할 때 꽃을 고정하는 줄기에 대한 브라트의 위치는 다른 용어의 사용으로 표시되며 유용한 진단 표시기가 될 수 있다.
일반적인 bract의 위치에는 다음이 포함된다.
- 어떤 식물들은 꽃들이 갈기 줄기에 있는 꽃의 꽃들을 미분하는 브랙트를 가지고 있다; 브랙트는 꽃을 들고 있는 줄기에 연결되지 않고, 장식되거나 주 줄기에 붙어 있다. (어드나이트는 서로 관련이 없는 다른 부분들의 혼합을 묘사한다.)서로 융합된 부품들이 동일할 때, 그들은 의도적으로 결합된다.)
- 다른 식물들은 단일 꽃의 발판이나 발판을 보조하는 역할을 한다.
교정조치의 전이적 위치에는 다음이 포함된다.
- 꽃(페디컬 또는 발톱)을 들고 있는 줄기에 브랙트를 붙이면 다시 발현된다고 하며, 때로는 이러한 브랙트나 브랙테롤이 크게 변형되어 꽃칼릭스의 덧장식으로 보인다.재결핵은 줄기가 꽃봉오리나 꽃봉오리를 잡고 있는 미분엽을 융합한 것으로, 따라서 잎이나 브랙트는 꽃봉오리에 장식된다.[4]
- 봉오리의 형성이 미분 잎 위로 뚜렷이 줄기 위로 이동될 때, 그것은 결석이라고 묘사된다.
.svg)

.svg)

.svg)

조직
다른 꽃가루를 정의하는데 일반적인 동의는 없다.다음은 Focko Weberling의 Morphologie der Blüten und der Blütensternde (Stuttgart, 1981년)에 근거한 것이다.염증의 주요 집단은 가지에 의해 구별된다.이러한 그룹 내에서 가장 중요한 특성은 축의 교차점 및 모델의 다른 변형이다.그들은 많은 꽃들을 포함할 수 있다.염증은 단순하거나 복합적일 수 있다.
단순염색증

미확정 또는 레이스모스
불확실한 단순 염증은 일반적으로 레이스모스/ˈræsɪmoʊs/라고 불린다.레이스모스 꽃의 주요 종류는 레이스미(/ˈræsiːm/, 고전 라틴 레이스무스로부터, 포도의 군집)이다.[5]다른 종류의 레이스모스 인플레 현상은 모두 다른 축의 팽창, 압축, 팽창 또는 감소에 의해 이것으로부터 파생될 수 있다.명백한 것 사이의 일부 통로의 양식은 일반적으로 인정된다.
- 경주마는 축을 따라 페디셀레이트(짧은 꽃 줄기를 가지고 있는) 꽃이 핀, 갈지 않고 불규칙한 꽃이다.
- 스파이크는 페디셀이 없는 꽃이 있는 경주마의 일종이다.
- 레이스모스 코리움은 내부보다 점차적으로 긴 바깥쪽 페디셀 때문에 평평하게 펴지거나 볼록하게 부풀어 오른다.
- 탯줄은 짧은 축과 길이가 같은 다수의 꽃무늬 페디셀을 가진 경주미의 일종으로 공통점에서 발생하는 것으로 보인다.그것은 움벨리퍼아이의 특징이다.
- 첨탑은 그 주위에 빽빽하게 배열된 꽃의 스파이크를 말하며, 주걱이라고 불리는 매우 전문화된 브라크를 감싸거나 동반한다.아리아과의 특색이다.
- 꽃 머리나 모피툴럼은 하나의 세실 꽃이 공유하는 매우 수축된 종족이다.딥사카과의 특징이다.
- 캣킨이나 아멘트는 비늘이 있고 일반적으로 축 처진 스파이크나 경주용 스파이크다.피상적으로 비슷한 사이모스나 다른 복잡한 염증도 일반적으로 이렇게 부른다.

스파이크

플랜타고 매체(스파이크)

레이스모스 코리움

이베리스 움벨라타(racemose corymb)

아스트란티아 마이너(우멜)

아룸 마쿨라툼(스파딕스)

머리(원형)

딥사쿠스 풀오넘(머리)

캣킨(레이스모스 또는 스프레이)

알누스 잉카나 (증정)
.svg)


.svg)




.svg)

.svg)

결정 또는 시모스
결정적으로 단순한 염증을 일으키는 것을 일반적으로 사이모스라고 부른다.시모스 팽창의 주요 종류는 시메('saim', 라틴 사이마에서 '배추새싹'이라는 의미로, 그리스 쿠마 '아무거나 부은 것'[6][7]으로 발음)이다.Cymes는 이 계획에 따라 더욱 세분된다.
- 단 하나의 보조 축: 모노카슘
- 이차 축 2개: 이차 축
- 2차축은 여전히 이분법: 이분법(Caryophyllus의 특성)
- 2차 축 단색성: 이중 전갈자리 cyme 또는 이중 헬리코이드 cyme
- 2차축 2개 이상: 플레오차슘
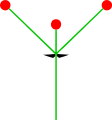
모노카슘

쌍싸이메
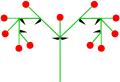
쌍싸이메
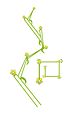
보스트리x(측면 및 상단 보기)

Hypericum 천공(보스트리x)
드레파늄(측면 및 상단)

글라디올루스 임브리카투스 (드레파늄)

신시누스(측면 및 상단)

심피툼 오피시날레(신시누스)

Rhipidium(측면 및 상단 보기)

캔나 sp. (리피듐)

디카시움
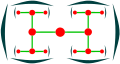
디카시움, 맨 위 보기

실렌다이오이카(디차슘)
.svg)
.svg)
.svg)
.jpg)

.jpg)



.jpg)

.svg)
_(inflorescence).svg)

cyme는 또한 탯줄처럼 보일 정도로 압축될 수 있다.엄밀히 말하면 이런 종류의 염증은 보통 단순히 'umbel'이라고 부르지만 탯줄형 cyme이라고 할 수 있다.
또 다른 종류의 확실한 단순 팽창은 종말꽃을 가진 종말의 종말의 종말의 종말의 종말의 종말의 종말의 종말의 종말의 종말의 종말의 종말의 종말의 종말의 종말의 종말로서 보통 부적절하게 '종말의 종말'이라고 불린다.
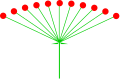
움벨리폼사이메

펠라르고늄 조날레(우멜형 종양)

보트료이드

베르베리스 버나에(보트리오이드)

.svg)

브랙트의 도끼 속에서 자라는 축소된 종족이나 종족(cyme)을 파시클이라고 한다.verticillaster는 디카시움의 구조를 가진 페시클로서, 라미아과에서 흔히 볼 수 있다.가시가 줄어든 많은 진균자들은 보통 스파이크라고 불리는 뾰루지(스파이크 유사) 염증을 형성할 수 있다.



복합염색증
단순 염증은 복합 염증이나 동시 염증의 기초가 된다.단화꽃은 단순한 꽃으로 대체되며, 그것은 경주용이나 시모스가 될 수 있다.복합성 염증은 갈린 줄기로 구성되며 본점까지 추적하기 어려운 복잡한 배치를 수반할 수 있다.
복합형 인플레스의 한 종류는 이중 인플레로, 단일 플로레트 대신 기본 구조가 반복된다.예를 들어, 더블 레이스미(double raceme)는 하나의 꽃이 다른 단순한 레이스미(classme)로 대체되는 레이스미(classme)이다. 같은 구조를 반복하여 트리플 또는 더 복잡한 구조를 형성할 수 있다.
복합 레이스미 인플레 현상은 최종 레이스미(호모티컬)로 끝날 수도 있고, 그렇지 않을 수도 있다.복합 경주마는 흔히 패니클이라고 불린다.이 정의는 베벌링이 제공한 정의와 매우 다르다는 점에 유의하십시오.
복합 탯줄은 탯줄이라 불리는 많은 작은 탯줄로 대체되는 탯줄이다.옆쪽 탯줄을 주줄기에 붙이는 줄기를 광선이라고 한다.

동심 화합물 레이스미

멜릴로투스 주례(동음이의 화합물 레이스미)
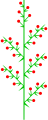
이질화합물 레이스미

헤베 알비칸 (히테로틱 화합물 경주)

컴파운드 스파이크

로륨 테만티움(복합 스파이크)

화합물 카피툴럼

에치놉스 리트로 (복합 카피툴룸)

복합(이중) 탯줄

레이저피튬라티폴륨(이중 탯줄)
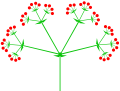
복합(트리플) 탯줄
.svg)




.svg)



.svg)
가장 흔한 종류의 확실한 복합적 팽창은 패니클(Webeling, 또는 '패니클 같은 사이메')이다.패니클(panicle)은 위에서 아래로 점점 더 강하게 불규칙하게 갈라지고, 가지마다 말단 꽃이 피는 확실한 꽃이다.
이른바 사이모스 코롬은 레이스모스 코롬과 비슷하지만 패니클 같은 구조를 갖고 있다.또 다른 형태의 패니클로는 안델라다.안테라는 중앙보다 측면 꽃이 높은 사이모스 코리움이다.

비티스비니페라(패니클)

삼부쿠스 니그라 (사이모스 코리움)

안델라

융커스 orolidus (안델라)


.svg)

.svg)

단 한 송이의 꽃이 cymes로 대체되는 종족(종족)을 (불확실한) 흉선이라고 한다.2차 cymes는 다른 형태의 dichasia와 monochasia일 수 있다.단 한 송이의 꽃이 cymes로 대체되는 보트리오이드(botryoid)는 확실한 흉선 또는 흉선이다.흉선은 종종 혼란스럽게 패니클이라고 불린다.

티르스

티르소이드

.svg)

다른 조합도 가능하다.예를 들어 머리나 탯줄은 corymb나 panicle로 배열할 수 있다.
_-_16.JPG)

기타
아스테라과 과는 기술적으로 칼라티드라고 불리는 매우 전문화된 두상으로 특징지어진다. (capitulum)' 또는 '머리'라고 부른다)에 의해 특징지어진다.포아과(Poikae) 계열은 보통 단순하고 부적절하게 스파이크와 패니클이라고 부르는 패니클이나 스파이크에 조직된 작은 스파이크(spikelet)의 독특한 팽창성을 가지고 있다.피쿠스속(Morae)은 시코늄이라고 불리는 염증을 가지고 있고, 유포르비아속에는 시아티아(sing. cyathium)가 있는데, 보통 탯줄로 조직되어 있다.

마트리카리아 카모밀라(칼라티드)

삼중수소 미학(복합 스파이크, "스파이크")

오리자 사티바(패니클의 스파이크, "패니클")

피쿠스 카리카 (시코늄)

유포르비아삼지타타(시아튬)

유포르비아시파리시(탯줄 속의 시아티아)

콜러스(거짓 스파이크)







일부 종은 복합화 또는 pseudania로 인플레 현상이 감소하는데, 이 경우 인플레와 단일 꽃을 구별하기 어렵다.[8]
개발 및 패터닝
개발
유전적 근거
염기 발달을 형성하는 유전자는 아라비독시스(Abraidopsis)에서 상당히 오랫동안 연구되어 왔다.LIPY(LFY)는 아라비독시스(Arabidopsis)에서 꽃무늬 메리스템 정체성을 촉진하는 유전자로, 꽃무늬의 발달을 조절한다.[9]LFY 표현의 타이밍을 변경하면 발전소에서 다른 염증이 발생할 수 있다.[10]LFY와 기능이 유사한 유전자는 APETALA1(AP1)이다.LFY, AP1 및 이와 유사한 촉진 유전자의 돌연변이는 꽃을 새싹으로 변환시키는 원인이 될 수 있다.[9]LEFY와는 대조적으로, 단자꽃(TFL)과 같은 유전자는 꽃들이 인플레시펙스(꽃 프리스트미엄 시작) 위에서 자라는 것을 방지하는 억제제의 활동을 지원하여 인플레시즘 메리스템 정체성을 유지한다.[11]두 종류의 유전자는 모두 꽃의 발달의 ABC 모델에 따라 꽃의 발달을 형성하는데 도움을 준다.최근 다른 꽃종에서 이러한 유전자의 호몰로로그에 대한 연구가 진행 중이거나 진행 중이다.
환경 영향
꽃가루를 먹는 곤충 초식동물은 평생의 체력(꽃 피는 정도), 꽃가루에 의한 종자 생산, 식물 밀도 등을 줄여 꽃가루를 형성한다.[12]이 초식물이 없을 때, 꽃들은 보통 더 많은 꽃 머리와 씨앗을 생산한다.[12]온도는 또한 다양한 형태의 염기 발달을 형성할 수 있다.고온은 특정 종에서 꽃봉오리의 적절한 발달을 저해하거나 꽃봉오리의 발달을 지연시킬 수 있는 반면, 다른 종에서는 기온의 상승이 꽃봉오리의 발달을 앞당길 수 있다.[13][14][15]
메리스템과 인플레센스 아키텍처
식물체에서 꽃의 생식기로의 전환은 꽃의 메리스템을 생성하는 꽃무늬 메리스템의 개발을 포함한다.[16]식물성 꽃 건축은 어떤 메리템이 꽃이 되고 어떤 것이 새싹이 되는가에 달려 있다.[17]결과적으로, 꽃무늬 메리스템 정체성을 조절하는 유전자는 꽃의 표현 영역이 식물의 꽃이 형성되는 위치를 지시하기 때문에 꽃무늬를 결정하는 데 중요한 역할을 한다.[16]
더 큰 규모로 볼 때, 팽창 구조는 자기 계발과 교차에서 오는 자손의 질과 양에 영향을 미친다. 그 구조는 수분 성공에 영향을 미칠 수 있기 때문이다.예를 들어, 아스클레피아스 꽃병은 같은 식물의 꽃 사이 또는 같은 꽃 사이의 교차점에 의한 자가 오염 수준에 의해 형성되는 상한 크기를 가지고 있는 것으로 나타났다.[18]애스쿨루스 실바티카에서는 가장 흔한 꽃가루 크기가 가장 높은 과일 생산과 상관관계가 있는 것으로 나타났다.[19]
참조
- ^ Guertin, P, Barnett, L, Denny, E.G, Schaffer, S.N. 2015.미국 국립 페놀로지 네트워크 식물학 프라이머.USA-NPN 교육 및 참여 시리즈 2015-001. www.usanpn.org
- ^ Kirchoff, Bruce K.; Claßen-Bockhoff, Regine (2013). "Inflorescences: concepts, function, development and evolution". Annals of Botany. 112 (8): 1471–6. doi:10.1093/aob/mct267. PMC 3828949. PMID 24383103.
- ^ Bradley, Desmond; Ratcliffe, Oliver; Vincent, Coral; Carpenter, Rosemary; Coen, Enrico (1997-01-03). "Inflorescence Commitment and Architecture in Arabidopsis". Science. 275 (5296): 80–83. doi:10.1126/science.275.5296.80. ISSN 0036-8075. PMID 8974397. S2CID 20301629.
- ^ 쿠비츠키, 클라우스, 클레멘스 바이어. 2002년.꽃이 피는 식물, 디코틸돈: 말발레, 카파랄레스, 그리고 비베탈레인 카리오필라목이다. 혈관 식물의 가족 및 일반, 5. 베를린: 스프링거. 페이지 77
- ^ 옥스퍼드 영어 사전경주미 2. 봇.꽃이 하나의 긴 축을 따라 같은 거리에서 짧고 거의 동일한 측면 페디셀에 배열되는 꽃의 일종
- ^ 콜린스 영어사전.2006년에 처음 출판된 제8판
- ^ 옥스퍼드 영어 사전사이메(1) 봇.1차 축에 먼저 발달하는 단자화(단자화)가 있는 팽창의 한 종류로, 2차 및 상위 순서의 축에 의해 시스템이 지속되며, 유사한 방식으로 연속적으로 발달한다. 원심 또는 확실한 팽창: 레이스미와는 반대되는 것이다.다소 평평한 헤드를 형성하는 이러한 유형의 복합 인플레에 esp.를 적용.
- ^ Tucker, Shirley C.; Grimes, James (1999-10-01). "The inflorescence: Introduction". The Botanical Review. 65 (4): 303–316. doi:10.1007/BF02857752. ISSN 0006-8101. S2CID 29599096.
- ^ a b Shannon, S.; Meeks-Wagner, D. R. (1993-06-01). "Genetic Interactions That Regulate Inflorescence Development in Arabidopsis". The Plant Cell. 5 (6): 639–655. doi:10.1105/tpc.5.6.639. ISSN 1040-4651. PMC 160302. PMID 12271079.
- ^ Schultz, E. A.; Haughn, G. W. (1991-08-01). "LEAFY, a Homeotic Gene That Regulates Inflorescence Development in Arabidopsis". The Plant Cell. 3 (8): 771–781. doi:10.1105/tpc.3.8.771. ISSN 1040-4651. PMC 160044. PMID 12324613.
- ^ Alvarez, John; Guli, Catherine L.; Yu, Xiang-Hua; Smyth, David R. (1992-01-01). "terminal flower: a gene affecting inflorescence development in Arabidopsis thaliana". The Plant Journal. 2 (1): 103–116. doi:10.1111/j.1365-313X.1992.00103.x. ISSN 1365-313X.
- ^ a b Louda, Svata M.; Potvin, Martha A. (1995-01-01). "Effect of Inflorescence-Feeding Insects on the Demography and Lifetime of a Native Plant". Ecology. 76 (1): 229–245. doi:10.2307/1940645. ISSN 1939-9170. JSTOR 1940645.
- ^ Moss, G (27 November 2015). "Influence of Temperature and Photoperiod on Flower Induction and Inflorescence Development in Sweet Orange (Citrus Sinensis L. Osbeck)". Journal of Horticultural Sciences. 44 (4): 311–320. doi:10.1080/00221589.1969.11514314.
- ^ Bjorkman, T.; Pearson, K. J. (1998-01-01). "High temperature arrest of inflorescence development in broccoli (Brassica oleracea var. italica L.)". Journal of Experimental Botany. 49 (318): 101–106. doi:10.1093/jxb/49.318.101. ISSN 0022-0957.
- ^ BREWSTER, J. L. (1983-04-01). "Effects of Photoperiod, Nitrogen Nutrition and Temperature on Inflorescence Initiation and Development in Onion (Allium cepa L.)". Annals of Botany. 51 (4): 429–440. doi:10.1093/oxfordjournals.aob.a086487. ISSN 0305-7364.
- ^ a b Souer, E.; Krol, A. van der; Kloos, D.; Spelt, C.; Bliek, M.; Mol, J.; Koes, R. (1998-02-15). "Genetic control of branching pattern and floral identity during Petunia inflorescence development". Development. 125 (4): 733–742. doi:10.1242/dev.125.4.733. ISSN 0950-1991. PMID 9435293.
- ^ Benlloch, R.; Berbel, A.; Serrano-Mislata, A.; Madueno, F. (2007-09-01). "Floral Initiation and Inflorescence Architecture: A Comparative View". Annals of Botany. 100 (3): 659–676. doi:10.1093/aob/mcm146. ISSN 0305-7364. PMC 2759223. PMID 17679690.
- ^ WYATT, ROBERT (1980-05-01). "The Reproductive Biology of Asclepias Tuberosa: I. Flower Number, Arrangement, and Fruit-Set". New Phytologist. 85 (1): 119–131. doi:10.1111/j.1469-8137.1980.tb04453.x. ISSN 1469-8137.
- ^ Wyatt, Robert (1982-04-01). "Inflorescence Architecture: How Flower Number, Arrangement, and Phenology Affect Pollination and Fruit-Set". American Journal of Botany. 69 (4): 585–594. doi:10.1002/j.1537-2197.1982.tb13295.x. ISSN 1537-2197. JSTOR 2443068.
참고 문헌 목록
- Focko Weberling: Morphologie der Blüten und der Blütensternde; Zweiter Teil.Verlag Eugen Ulmer, Stuttgart 1981
- 빌헬름 트롤: 디 인플로레젠; 에스터 밴드.구스타프 피셔 베를라크, 슈투트가르트 1964
- 빌헬름 트롤: 디 인플로레스젠; 즈웨이터 밴드, 에르스터 틸.구스타프 피셔 베를라크, 슈투트가르트 1969
- 빌헬름 트롤: 프라크티스체 에인퓌룽 다이 프란젠모폴로니.구스타프 피셔 베를라크, 제나 1957
- 베른하르트 카우스만: 프란제나나토미.구스타프 피셔 베를라크, 제나 1963
- 월터 S. 저드, 크리스토퍼 S. 캠벨, 엘리자베스 A. 켈로그, 피터 F.스티븐스, Michael J. Donoghue: 식물 체계론: 시나우어 어소시에이츠 주식회사의 계발적 접근법
- 스티븐스, P.F.(2001년 이후)Angiosperm Phylogeny 웹사이트[1].버전 7, 2006년 5월 [그 후 또는 그 이하가 지속적으로 업데이트됨]
- 스트라스버거, 눌, 스헨크, 딤퍼: 르르부흐 데르 보타닉 퓌르 홉슈천엔. 4.아우플라주, 구스타프 피셔, 제나 1900, 페이지 459
- R J 페리.염증 및 그 이름.맥알렌 국제 난초 협회 저널.제12권(6), 페이지 4-11 2011년 6월
외부 링크
 위키미디어 커먼스의 인플레시젼 관련 매체
위키미디어 커먼스의 인플레시젼 관련 매체