린초케팔리아
Rhynchocephalia| 린코케팔리아스 | |
|---|---|
 | |
| 살아있는 유일한 린코세팔리아류인 투아타라(Sphenodon punctatus) | |
.jpg) | |
| 독일 쥐라기 후기의 운조류인 바다사우루스의 화석 | |
| 과학적 분류 | |
| 도메인: | 진핵생물 |
| 왕국: | 애니멀리아 |
| 문: | 초르다타 |
| 클래스: | 파충류 |
| 초순: | 레피도사우루스아과 |
| 순서: | 린초케팔리아 1867년 귄터 |
| 종류 | |
| 스페노돈펑터투스 그레이, 1842 | |
| 부분군 | |
| 본문을 보다 | |
린초케팔리아(/ˌ ɪŋ코 ʊ ɪˈ페 ɪ리 ə/; '비크헤드')는 뉴질랜드의 투아타라(Sphenodon punctatus) 단 하나의 살아있는 종을 포함하는 도마뱀과 같은 파충류 목입니다. 현재의 다양성 부족에도 불구하고, 중생대의 린초케팔리안(rynchocephalians)은 형태학적, 생태학적 다양성이 높은 종군이었습니다. 이 그룹의 가장 오래된 기록은 2억 3,800만년에서 2억 4,000만년 전의 트라이아스기 중기로 거슬러 올라가며, 그들은 쥐라기 초기에 전세계적인 분포를 이루었습니다.[1] 대부분의 린초케팔루스류는 스페노돈티아("wedge-tee") 그룹에 속합니다. 그들의 가장 가까운 살아있는 동족은 도마뱀과 뱀이며, 두 목은 레피도사우루스아목에 함께 묶였습니다.
오늘날 도마뱀이 차지하고 있는 많은 틈새시장은 트라이아스기와 쥐라기에 스페노돈티안이 차지하고 있었습니다. 린코케팔루스류는 백악기 동안 큰 쇠퇴를 겪었고, 신생대 초기에는 거의 완전히 사라졌습니다. 현대의 투아타라는 주로 육식성이지만, 잡식성(Opisthias), 초식성(Eilenodontinae), 십이지식성(sapheosaur) 생활양식을 가진 스페노돈티안(spenodontians)도 있었습니다. 심지어 플루로사우루스와 같은 수생 스페노돈티안의 성공적인 그룹도 있었습니다.[2]
역사
투아타라는 1831년 존 에드워드 그레이에 의해 처음으로 기술되었을 때 원래는 아마이드 도마뱀으로 분류되었습니다. 그들은 1867년 대영 박물관의 앨버트 귄터가 새, 거북이, 악어와 비슷한 특징을 언급할 때까지 잘못된 분류로 남아있었습니다. 그는 투아타라와 그 화석 친척들을 위해 린초케팔리아 목("beak head"라는 뜻)을 제안했습니다.[3] 1925년 새뮤얼 웬델 윌리스턴(Samuel Wendell Williston)은 스페노돈티아(Spenodontia)에 투아타라(tuatara)와 그들의 가장 가까운 화석 동족만을 포함시킬 것을 제안했습니다.[4] 스페노돈은 그리스 σφήν의 '웨지'와 ὀδούς의 '이빨'에서 유래했습니다. 이후에 린초세팔리아(Rynchocephalia)에 별도로 관련된 많은 종들이 추가되었으며 분류학자들은 이를 "쓰레기통 분류군"이라고 부릅니다. 이것들은 트라이아스기에 살았던 표면적으로 비슷하지만 (모양과 이름 모두에서) 관련이 없는 린코사우루스를 포함합니다.[4] 1970년대와 1980년대의 연구는 린코사우루스가 관련이 없다는 것을 보여주었고, 1980년대에 컴퓨터를 기반으로 한 분류학적 분석은 투아타라와 화석 린코세팔루스 사이의 밀접한 관계를 보여주었습니다.[8]
해부학

린초세팔리아와 그들의 자매 그룹인 스콰마타(도마뱀, 뱀, 양서류를 포함)는 레피도사우로모르파 내에서 유일하게 생존하는 분류군인 레피도사우루스아과에 속합니다.
스쿼메이트와 린코세팔리안은 꼬리 척추 내의 골절면을 포함하여 많은 공유된 특성(동형)을 가지고 있습니다. 꼬리 척추 내의 골절면은 꼬리 자가절개(위협을 받았을 때 꼬리의 손실), 가로 배설강 슬릿, 갑상선 페네스트라로 알려진 골반의 개구, 사지 뼈 에피피에 추가 골화 센터의 존재, 대퇴골의 측면 오목부가 섬유의 관절을 가능하게 하는 무릎 관절, 신장의 성 분절의 발달, 그리고 새로운 관절을 생성하는 융합된 아스트랄라고-calc은 및 확대된 제4원위부 족골, 그리고 후크된 제5중족골을 포함하는 발 뼈의 다양한 특성.
일부 스쿼마이트들과 마찬가지로 투아타라는 머리 위쪽에 두정안(솔방울눈 또는 제3의 눈이라고도 함)을 가지고 있으며, 두개골 지붕에 동반된 구멍이 있는데, 이 구멍은 "솔방울구멍"이라고도 하며, 이는 화석 린코세팔리아류에도 존재합니다. 두정안은 주간 및 계절 주기를 모니터링하는 빛을 감지하여 다른 기능 중에서도 일주기 리듬을 조절하는 데 도움을 줍니다.[10][11][12][13] 솔방울눈은 초기 파충류를 포함한 초기 척추동물들 사이에 널리 퍼져 있었지만 대부분의 살아있는 그룹들 사이에서는 사라졌습니다.[12] 린코케팔류는 가스트랄리아(몸의 배에 존재하는 갈비뼈와 같은 뼈, 조상적으로 사지동물에 존재하며 살아있는 악어에도 존재함)의 보유를 포함한 많은 특성에 의해 스쿼메이트와 구별됩니다.[14] 대부분의 새들과 마찬가지로, 스쿼메이트와 달리 투아타라에는 음경이 없습니다. 이것은 2차적인 손실인데, 아마도 음경이나 편평과 같은 헤미펜이 린코케팔리안과 편평의 마지막 공통 조상에 존재했을 것이기 때문입니다.[15]
역사적으로 종종 초기 파충류로부터 유지된 원시적인 특징이라고 주장되는 투아타라의 완전한 하부 시간 막대(jugal 및 quadratate/quadratojugal bone의 융합으로 인해 발생함)는 실제로 스페노돈트류 사이에서 파생된 특징이며, 원시적인 레피도사우로모프와 가장 원시적인 것들을 포함한 많은 린코케팔류와 함께 시간 막대 없이 개방된 하부 시간 울타리를 가지고 있습니다.[16][17]
.jpg)
홍조류의 몸 크기는 매우 다양합니다. 투아타라는 암컷과 수컷의 평균 총 길이가 각각 34.8 센티미터와 42.7 센티미터입니다.[18] 클레보사우루스 종파충류는 총 길이가 12센티미터(4.7인치)인 반면,[19] 가장 큰 육상 스페노돈트류인 프리오스페노돈타벨라시는 총 길이가 100센티미터(39인치)를 조금 넘습니다.[20] 수생 흉막류는 길이가 최대 150cm(59인치)에 달했습니다.[21]
.jpg)
투아타라를 포함한 대부분의 운모충류의 치열은 치아가 턱뼈의 볏에 붙어 있는 상태와 관련된 crodont로 묘사되며, 치아 대체가 부족하고 치아와 턱을 결합하여 광범위한 뼈 성장을 하여 치아와 뼈의 경계를 식별하기가 어렵습니다. 이것은 대부분의 도마뱀(acrodontan 제외)에서 발견되는 상태와 다릅니다. 이 도마뱀은 턱의 안쪽을 향하는 쪽의 선반에 부착되어 있고, 평생 동안 교체됩니다. 투아타라의 이빨은 뿌리가 없지만, 다른 일부 린코케팔류의 이빨은 뿌리를 가지고 있습니다.[22] 아크로덴트 치열은 더 원시적인 레피도사우로모프에서 발견되지 않는 린코케팔리안의 파생된 특성으로 보입니다.[23] 가장 원시적인 윤조류는 흉막치 또는 흉막치 앞니와 악막치 뒷니를 모두 가지고 있습니다.[22][24] 일부 린코세팔리안은 이러한 상태와 다르며, 안킬로스페노돈은 턱 뼈 깊숙이 계속되는 표면적으로 톱니가 있고 소켓(안킬로스테코돈)의 기저부에 있는 뼈와 융합되어 있습니다.[22] 대부분의 척추동물에서는 위턱 앞쪽에 있는 상악 전치가 큰 끌과 같은 구조로 합쳐져 있습니다.[25]
린코케팔리안은 구개(입천장의 뼈에 존재하는 치아)를 가지고 있습니다. 구개 치아는 조상적으로 사지동물에 존재하지만 많은 그룹에서 손실되었습니다.[26] 대부분의 윤조류는 익상골에 존재하는 치아는 소실되지만, 구개골에 존재하는 측치열은 확대되어 상악골의 치아와 평행하게 배향됩니다. 무는 동안 상악과 구개열 사이의 아래턱 슬롯에 있는 틀니의 치아. 양막 중에서도 독특한 이 배열은 음식물의 3점 굽힘을 가능하게 하며,[27] 프로팔라 동작(하악의 뒤와 앞으로 움직이는 동작)과 결합하여 전단 물림을 가능하게 합니다.[26][28]

분류
Rynchocephalia의 그룹화는 잘 뒷받침되지만 많은 분류군의 서로에 대한 관계는 불확실하여 연구마다 크게 다릅니다.[29] 현대의 분류학에서, 스페노돈티아 계통은 제피로사우루스와 다른 제피로사우루스류뿐만 아니라 위르템베르기아 이외의 모든 린코케팔루스류를 포함합니다. 게피로사우루스류는 일부 분석에서 스쿼메이트와 더 밀접한 관련이 있는 것으로 나타났습니다.[30][24] 2018년, Spenodontia 내의 두 개의 주요 분기군이 정의되었는데, Eusphenodontia는 Polysphenodon, Clevosaurus hudsoni 및 Spenodon을 포함하는 가장 덜 포괄적인 분기군에 의해 정의되며, 이는 세 개의 시냅스의 존재에 의해 뒷받침됩니다. 치아 또는 상악의 치아에 명확하게 보이는 마모면이 존재하는 것을 포함하여, 상악 전치는 끌과 같은 구조로 합쳐지고, 구개는 하나의 치아 열로 감소되고, 추가적인 고립된 치아가 존재하는 것을 특징으로 하는 치아. 순위가 매겨진 네오스페노돈티아(Neosphenodontia)는 스페노돈(Spenodon)을 포함하지만 클레보사우루스 허드슨(Clevosaurus hudsoni)은 포함하지 않는 가장 포괄적인 분기군으로 정의되며, 이는 두개골의 안와(antorbital) 부분의 상대적 길이 증가를 포함한 6개의 시냅스의 존재로 뒷받침됩니다. 전체 두개골 길이의 1/4~1/3에 이르며, 두정골의 뒤쪽(hind) 가장자리는 안쪽으로 약간만 휘어져 있고, 두정골 구멍은 상완골 회문의 앞쪽 경계와 같은 높이 또는 앞쪽에서 발견됩니다(두골의 입구), 구개 치아는 경막내 치아 상태에서 하나의 측면 치아열로 더 감소되고, 경막치열의 수는 1개 또는 1개로 감소되며, 좌골의 후방 경계는 특징적인 과정을 갖는 것을 특징으로 하는 치골.[25] 2021년에 Acrosphenodontia 분기군이 정의되었으며, 이는 Spenodontia보다 덜 포괄적이고 Eusphenodontia보다 더 포괄적이며 기저 부분적으로 acrodont spenodontian을 제외한 완전한 acrodontention을 가진 모든 spenodontian을 포함합니다.[31] 2022년에 멸종된 클레이드 렙토린키아(Leptorhynchia)가 정의되었습니다. 여기에는 다양한 신식각류가 포함되며, 적어도 일부는 수생적으로 적응되었으며, 네 번째 메타카팔(metacarpal)의 신장, 좌골(ischium)에 후방 과정이 존재하는 것이 특징입니다. 그리고 두개골의 안와 부분은 전체 두개골 길이의 3분의 1에서 4분의 1 사이입니다.[32] 오피스토돈티아 계통은 스페노돈보다는 프리오스페노돈(Eilenodontinae)과 더 밀접한 관련이 있는 모든 스페노돈류의 그룹화에 사용되었습니다.[33] 모든 연구에서 이 계통군을 사용하는 것은 아닙니다. 일부 연구에서 계통군의 범위가 Eilenodontinae와 동일한 것으로 밝혀졌기 때문입니다.[32]
스페노돈티과는 투아타라와 린초세팔리아 내의 가장 가까운 동족을 포함하는 데 사용되었습니다. 그러나 그룹화에는 공식적인 정의가 부족하여 포함된 분류군이 분석마다 크게 다릅니다.[30] 투아타라의 가장 가까운 동족은 완전히 닫힌 시간 막대를 특징으로 하는 스페노돈티에(Sphenodontinae) 분기군에 배치됩니다.[17]
다음은 DeMar et al. 2022 이후의 린초케팔리아의 클라도그램입니다.[32]
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Simoes et al: 2022년 이후의 Cladogram:[17]
| Sphenodontia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
분기군과 속
- †Wirtembergia
- †Gephyrosauridae?
- Sphenodontia 1925년 윌리스턴
- †디프히돈토사우루스
- †Micromenodon
- †팔롤라노사우루스?
- †Pelecymala
- †레바나사우루스
- †Sphenocondor
- Acrosphenodontia Chambi-Trowell et al., 2021
- †고다바리사우루스
- †플라노케팔사우루스
- †Theretairus
- Eusphenodontia Herrera-Flores et al. 2018
- †Opisthiamimus
- †Brachyrhinodon
- †클레보사우루스
- †콜롭스?
- †Lanceirosphenodon
- †Polysphenodon
- †시그말라
- 네오스페노돈티아 Herrera-Flores et al. 2018
- †라마르케사우루스
- †Pamizinsaurus
- †팅기타나
- †Whitakeraurus?
- †Ankylosphenodon
- †Opisthodontia
- 스페노돈티과
- †Leptorhynchia DeMar, Jones & Carrano, 2022
- †렙토사우루스
- †Kallimodon
- †호메오사우루스
- †바다사우루스
- †Sapheosauridae
- †Pleurosauridae
-
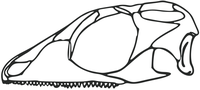
게피로사우루스의 두개골 재구성 가능한 기저 린코케팔리안
-

스페노돈티아의 기초적 구성원인 디프히돈토사우루스의 두개골 재건
-

Clevosaurus hudsoni (A)와 Clevosaurus cambrica (B)의 두개골 재건
-

기저 유스테노돈티안 오피스티아미무스의 두개골
-

-
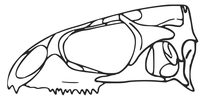
스페노돈티에(Spenodontinae)의 초기 구성원인 나바조스페노돈(Navajosphenodon)의 두개골 재건
-

-

네오스페노돈티안 바다사우루스의 두개골








생태학

린코세팔류의 화석 기록은 그들이 다양한 생태학적 틈새를 이용한 다양한 집단이었음을 보여줍니다.[3][27] 초기의 운조류는 피어싱을 위해 고안된 작은 난형 이빨을 가지고 있으며, 아마도 식충류였을 것입니다.[34] 현생 투아타라와 마찬가지로, 멸종된 스페노돈티에(Spenodontinae)의 구성원들은 육식성/곤충성 식단을 가진 일반론자였을 가능성이 높습니다.[35] 가장 뚜렷한 린코세팔루스 중 하나는 해양 생물에 적응한 유럽의 쥐라기 시대부터 알려진 플루로사우루스로, 팔다리가 줄어든 길쭉한 뱀처럼 생긴 몸을 가지고 있으며, 전문화된 후기 쥐라기 속 플루로사우루스는 다른 린코세팔루스류에서 고도로 변형된 길쭉한 삼각형 두개골을 가지고 있습니다. 흉각류는 물고기를 잡아먹는 육식동물이었던 것으로 생각됩니다.[21] 다른 여러 린코세팔리안 계통들은 반수생 습성을 가지고 있었던 것으로 제시되었습니다.[36] Eilenodontines는 식물 재료를 가공하는 데 두꺼운 법랑질을 가진 넓은 치아의 배터리와 함께 초식성이었던 것으로 생각됩니다.[37] 유럽 쥐라기 후기의 오에노사우루스와 사페오사우루스와 같은 사페오사우루스류는 네발동물 사이에서 독특한 넓은 이빨 판을 가지고 있으며 이빨 판은 딱딱한 껍질의 유기체를 으깨는 데 사용되는 것으로 생각됩니다.[38][30] 멕시코 쥐라기의 스페노비페라는 아래턱[39] 앞쪽에 두 개의 확대된 치아에 홈이 있는 것으로 보아 독이 있는 것으로 추정되지만 다른 저자들은 이 해석에 의문을 제기했습니다.[40]
진화사

린초케팔리아가 언제 스콰마타에서 갈라졌을 것으로 추정되는지에 대해서는 논쟁의 여지가 있습니다. 오래된 추정에 따르면 페름기 중기와 트라이아스기 초기의 차이는 약 2억 7,200만 년 전에서 2억 5,200만 년 전으로 추정되며 [30]다른 저자들은 약 2억 4,200만 년 전의 더 젊은 날짜를 제시합니다.[1] 알려진 가장 오래된 린초케팔리안의 유적은 독일 남부 벨베르크 근처의 에르푸르트 층에서 알려진 비르템베르기아의 유적으로, 약 2억 3,800만-2억 4,000만 년 된 중기 트라이아스기의 라디니아 단계까지 거슬러 올라갑니다. 이는 알려진 가장 원시적인 린초케팔리안이기도 합니다.[24] 린코케팔류는 트라이아스기 후기에 상당히 다양화되었고 [3]트라이아스기 말에 판게아 전역에 걸쳐 전 세계적으로 분포하게 되었으며 트라이아스기 후기-초기 쥐라기 속 클레보사우루스는 아시아, 아프리카, 유럽, 북미 및 남아메리카 전역에 걸쳐 10종을 가지고 있습니다.[41] 초기의 운조류는 작은 동물이었지만 트라이아스기 후기에 이르러 이 그룹은 다양한 신체 크기로 진화했습니다.[42] 쥐라기 동안 린코세팔류는 전문적인 초식 및 수생 형태를 포함하여 형태학적 다양성의 정점에 도달했습니다.[3] 아시아에서 온 린초케팔루스에 대한 유일한 기록(중생대에는 아시아의 일부가 아니었던 인도 아대륙 제외)은 중국 윈난성의 초기 쥐라기(시네무리아)의 클레보사우루스의 불확실한 유해입니다. 린코세팔리안은 보존 조건이 양호함에도 불구하고 이 지역의 젊은 지역에는 눈에 띄게 부재합니다.[43]
린코케팔류는 백악기 전기 이후 북아메리카와 유럽에서 사라졌고,[44] 백악기 전기에는 북아프리카와[45] 남아메리카[46] 북부에서 없었습니다. 린초케팔리아의 감소 원인은 아직 명확하지 않지만, 종종 첨단 도마뱀 및 포유류와의 경쟁에서 기인하는 것으로 제시되어 왔습니다.[47] 이들은 백악기 후기에 남아메리카 남부 지역에 널리 분포한 것으로 보이며, 도마뱀은 희귀한 상태로 남아 있으며, 이 지역의 육상 도마뱀보다 그들의 유해가 200배나 더 많습니다.[45] 백악기 후기 남아메리카의 스페노돈티우스는 엘라이엔돈티에(Eliendontinae)와 스페노돈티에(Sphenodontinae)로 대표되며, 엘리엔돈티에에 대한 가장 어린 기록은 투로니안과(Turonian)이다.[48] 불확실한 린초케팔리안은 백악기 후기 또는 아마도 가장 초기의 인도 인섬의 고생대에서 알려져 있으며, 이는 기초적인 아크로스페돈티안으로 보입니다.[49] 뉴질랜드 이외의 지역에서 가장 젊은 의심의 여지가 없는 린초케팔리안의 유적은 백악기-팔레오네 멸종 사건 직후 파타고니아의 초기 고생대(다니안)에서 발견된 스페노돈티드 카와스페노돈 펠리그렌시스의 유적입니다.[50] 치아를 가진 불확정의 스페노돈틴 턱 조각은 중신세 초기(19-1600만년 전)의 뉴질랜드 세인트 배산스 동물군에서 알려져 있으며, 이는 살아있는 투아타라와 구별할 수 없습니다. 투아타라의 조상이 해양 분산을 통해 뉴질랜드에 도착했을 가능성은 거의 없으며, 8천만 년에서 6천 6백만 년 전 사이에 남극에서 분리되었을 때 이미 뉴질랜드에 존재했을 것으로 추정됩니다.[47]
참고문헌
- ^ a b Jones ME, Anderson CL, Hipsley CA, Müller J, Evans SE, Schoch RR (September 2013). "Integration of molecules and new fossils supports a Triassic origin for Lepidosauria (lizards, snakes, and tuatara)". BMC Evolutionary Biology. 13: 208. doi:10.1186/1471-2148-13-208. PMC 4016551. PMID 24063680.
- ^ Reynoso VH (2000). "An unusual aquatic sphenodontian (Reptilia: Diapsida) from the Tlayua Formation (Albian), central Mexico". Journal of Paleontology. 74 (1): 133–148. Bibcode:2000JPal...74..133R. doi:10.1017/s0022336000031310. S2CID 232346834.
- ^ a b c d Herrera-Flores JA, Stubbs TL, Benton MJ (2017). "Macroevolutionary patterns in Rhynchocephalia: is the tuatara (Sphenodon punctatus) a living fossil?". Palaeontology. 60 (3): 319–328. Bibcode:2017Palgy..60..319H. doi:10.1111/pala.12284. ISSN 1475-4983.
- ^ a b Fraser N, Sues HD, eds. (1994). "Phylogeny" In the Shadow of the Dinosaurs: Early Mesozoic Tetrapods. Cambridge University Press. ISBN 0-521-45242-2.
- ^ Evans SE, Prasad GV, Manhas BK (November 2001). "Rhynchocephalians (Diapsida: Lepidosauria) from the Jurassic Kota Formation of India". Zoological Journal of the Linnean Society. 133 (3): 309–34. doi:10.1111/j.1096-3642.2001.tb00629.x.
- ^ "Sphenodon". Dictionary.com Unabridged (v 1.1). Random House, Inc. Retrieved 8 January 2007.
- ^ Evans SE, Borsuk-Białynicka M (2009). "A small lepidosauromorph reptile from the Early Triassic of Poland" (PDF). Paleontologica Polonica. 65: 179–202.
- ^ 고티에, J., Estes, R. & De Quiroz, K. (1988) 레피도사우로모르파의 계통발생학적 분석. 도마뱀과의 계통발생학적 관계에서 (eds R). Estes and G. Pregill), pp. 15–98. 스탠퍼드 대학 출판부, 스탠퍼드
- ^ "Rhynchocephalians". University College London. Retrieved 2022-08-09.
- ^ Dendy A (1911). "VI. On the structure, development and morphological interpretation of the pineal organs and adjacent parts of the brain in the tuatara ( sphenodon punctatus )". Philosophical Transactions of the Royal Society B. 201 (274–281): 227–331. doi:10.1098/rstb.1911.0006. ISSN 0264-3960.
- ^ Smith, Krister T.; Bhullar, Bhart-Anjan S.; Köhler, Gunther; Habersetzer, Jörg (2 April 2018). "The only known jawed vertebrate with four eyes and the bauplan of the pineal complex". Current Biology. 28 (7): 1101–1107.e2. doi:10.1016/j.cub.2018.02.021. ISSN 0960-9822. PMID 29614279.
- ^ a b Benoit, Julien; Abdala, Fernando; Manger, Paul; Rubidge, Bruce (2016). "The sixth sense in mammalians forerunners: variability of the parietal foramen and the evolution of the pineal eye in South African Permo-Triassic eutheriodont therapsids". Acta Palaeontologica Polonica. doi:10.4202/app.00219.2015.
- ^ Paulina-Carabajal, Ariana; Jiménez-Huidobro, Paulina; Triviño, Laura Natalia; Stanley, Edward L.; Zaher, Hussam; Daza, Juan D. (2023), Dozo, María Teresa; Paulina-Carabajal, Ariana; Macrini, Thomas E.; Walsh, Stig (eds.), "A Look in to the Neurocranium of Living and Extinct Lepidosauria", Paleoneurology of Amniotes, Cham: Springer International Publishing, pp. 123–177, doi:10.1007/978-3-031-13983-3_5, ISBN 978-3-031-13982-6, retrieved 2023-12-08
- ^ Vitt LJ, Caldwell JP (2014). "Chapter 20: Rhynchocephalians (Sphenodontids)". Herpetology. Elsevier. pp. 553–554. doi:10.1016/b978-0-12-386919-7.00020-4. ISBN 978-0-12-386919-7.
- ^ Sanger TJ, Gredler ML, Cohn MJ (October 2015). "Resurrecting embryos of the tuatara, Sphenodon punctatus, to resolve vertebrate phallus evolution". Biology Letters. 11 (10): 20150694. doi:10.1098/rsbl.2015.0694. PMC 4650183. PMID 26510679.
- ^ Evans SE, Jones ME (2010). "The Origin, Early History and Diversification of Lepidosauromorph Reptiles". New Aspects of Mesozoic Biodiversity. Lecture Notes in Earth Sciences. Vol. 132. Berlin, Heidelberg: Springer Berlin Heidelberg. pp. 27–44. Bibcode:2010LNES..132...27E. doi:10.1007/978-3-642-10311-7_2. ISBN 978-3-642-10310-0.
- ^ a b c Simões TR, Kinney-Broderick G, Pierce SE (March 2022). "An exceptionally preserved Sphenodon-like sphenodontian reveals deep time conservation of the tuatara skeleton and ontogeny". Communications Biology. 5 (1): 195. doi:10.1038/s42003-022-03144-y. PMC 8894340. PMID 35241764.
- ^ Herrel, Anthony; Moore, Jennifer A.; Bredeweg, Evan M.; Nelson, Nicola J. (2010-05-19). "Sexual dimorphism, body size, bite force and male mating success in tuatara: SEXUAL DIMORPHISM IN TUATARA". Biological Journal of the Linnean Society. 100 (2): 287–292. doi:10.1111/j.1095-8312.2010.01433.x.
- ^ Keeble, Emily; Whiteside, David I.; Benton, Michael J. (April 2018). "The terrestrial fauna of the Late Triassic Pant-y-ffynnon Quarry fissures, South Wales, UK and a new species of Clevosaurus (Lepidosauria: Rhynchocephalia)". Proceedings of the Geologists' Association. 129 (2): 99–119. doi:10.1016/j.pgeola.2017.11.001.
- ^ Apesteguía, Sebastián; Novas, Fernando E. (9 October 2003). "Large Cretaceous sphenodontian from Patagonia provides insight into lepidosaur evolution in Gondwana". Nature. 425 (6958): 609–612. Bibcode:2003Natur.425..609A. doi:10.1038/nature01995. PMID 14534584. S2CID 4425130.
- ^ a b Klein N, Scheyer TM (February 2017). "Microanatomy and life history in Palaeopleurosaurus (Rhynchocephalia: Pleurosauridae) from the Early Jurassic of Germany". Die Naturwissenschaften. 104 (1–2): 4. Bibcode:2017SciNa.104....4K. doi:10.1007/s00114-016-1427-3. PMID 28005148. S2CID 27133670.
- ^ a b c Jenkins KM, Jones ME, Zikmund T, Boyde A, Daza JD (September 2017). "A Review of Tooth Implantation Among Rhynchocephalians (Lepidosauria)". Journal of Herpetology. 51 (3): 300–306. doi:10.1670/16-146. ISSN 0022-1511. S2CID 90519352.
- ^ Ford, David P.; Evans, Susan E.; Choiniere, Jonah N.; Fernandez, Vincent; Benson, Roger B. J. (2021-08-25). "A reassessment of the enigmatic diapsid Paliguana whitei and the early history of Lepidosauromorpha". Proceedings of the Royal Society B: Biological Sciences. 288 (1957): 20211084. doi:10.1098/rspb.2021.1084. ISSN 0962-8452. PMC 8385343. PMID 34428965.
- ^ a b c Sues, Hans‐Dieter; Schoch, Rainer R. (2023-11-07). "The oldest known rhynchocephalian reptile from the Middle Triassic (Ladinian) of Germany and its phylogenetic position among Lepidosauromorpha". The Anatomical Record. doi:10.1002/ar.25339. ISSN 1932-8486.
- ^ a b Herrera-Flores JA, Stubbs TL, Elsler A, Benton MJ (2018-04-06). "Taxonomic reassessment of Clevosaurus latidens Fraser, 1993 (Lepidosauria, Rhynchocephalia) and rhynchocephalian phylogeny based on parsimony and Bayesian inference". Journal of Paleontology. 92 (4): 734–742. Bibcode:2018JPal...92..734H. doi:10.1017/jpa.2017.136.
- ^ a b Matsumoto R, Evans SE (January 2017). "The palatal dentition of tetrapods and its functional significance". Journal of Anatomy. 230 (1): 47–65. doi:10.1111/joa.12534. PMC 5192890. PMID 27542892.
- ^ a b Jones ME (August 2008). "Skull shape and feeding strategy in Sphenodon and other Rhynchocephalia (Diapsida: Lepidosauria)". Journal of Morphology. 269 (8): 945–966. doi:10.1002/jmor.10634. PMID 18512698.
- ^ Jones ME, O'higgins P, Fagan MJ, Evans SE, Curtis N (July 2012). "Shearing mechanics and the influence of a flexible symphysis during oral food processing in Sphenodon (Lepidosauria: Rhynchocephalia)". Anatomical Record. 295 (7): 1075–1091. doi:10.1002/ar.22487. PMID 22644955.
- ^ Romo de Vivar PR, Martinelli AG, Schmaltz Hsiou A, Soares MB (2020-07-02). "A New Rhynchocephalian from the Late Triassic of Southern Brazil Enhances Eusphenodontian Diversity". Journal of Systematic Palaeontology. 18 (13): 1103–1126. doi:10.1080/14772019.2020.1732488. ISSN 1477-2019. S2CID 216226211.
- ^ a b c d Simões TR, Caldwell MW, Pierce SE (December 2020). "Sphenodontian phylogeny and the impact of model choice in Bayesian morphological clock estimates of divergence times and evolutionary rates". BMC Biology. 18 (1): 191. doi:10.1186/s12915-020-00901-5. PMC 7720557. PMID 33287835.
- ^ Chambi-Trowell, Sofia A. V.; Martinelli, Agustín G.; Whiteside, David I.; Vivar, Paulo R. Romo de; Soares, Marina Bento; Schultz, Cesar L.; Gill, Pamela G.; Benton, Michael J.; Rayfield, Emily J. (2021-06-03). "The diversity of Triassic South American sphenodontians: a new basal form, clevosaurs, and a revision of rhynchocephalian phylogeny". Journal of Systematic Palaeontology. 19 (11): 787–820. doi:10.1080/14772019.2021.1976292. hdl:1983/af14affc-a26e-426b-83ca-e1833e355882. ISSN 1477-2019. S2CID 240487298.
- ^ a b c DeMar, David G.; Jones, Marc E. H.; Carrano, Matthew T. (2022-12-31). "A nearly complete skeleton of a new eusphenodontian from the Upper Jurassic Morrison Formation, Wyoming, USA, provides insight into the evolution and diversity of Rhynchocephalia (Reptilia: Lepidosauria)". Journal of Systematic Palaeontology. 20 (1): 1–64. doi:10.1080/14772019.2022.2093139. hdl:2440/136608. ISSN 1477-2019. S2CID 252325953.
- ^ Apesteguía, Sebastián; Novas, Fernando E. (2003-10-09). "Large Cretaceous sphenodontian from Patagonia provides insight into lepidosaur evolution in Gondwana". Nature. 425 (6958): 609–612. doi:10.1038/nature01995. ISSN 0028-0836.
- ^ Jones ME (2009). Koppe T, Meyer G, Alt KW, Brook A (eds.). "Dentary Tooth Shape in Sphenodon and Its Fossil Relatives (Diapsida: Lepidosauria: Rhynchocephalia)". Frontiers of Oral Biology. Basel: Karger. 13: 9–15. doi:10.1159/000242382. ISBN 978-3-8055-9229-1. PMID 19828962.
- ^ Villa, Andrea; Montie, Roel; Röper, Martin; Rothgaenger, Monika; Rauhut, Oliver W. M. (2021-05-03). "Sphenofontis velserae gen. et sp. nov., a new rhynchocephalian from the Late Jurassic of Brunn (Solnhofen Archipelago, southern Germany)". PeerJ. 9: e11363. doi:10.7717/peerj.11363. ISSN 2167-8359. PMC 8101455. PMID 33987027.
- ^ Bever GS, Norell MA (November 2017). "A new rhynchocephalian (Reptilia: Lepidosauria) from the Late Jurassic of Solnhofen (Germany) and the origin of the marine Pleurosauridae". Royal Society Open Science. 4 (11): 170570. doi:10.1098/rsos.170570. PMC 5717629. PMID 29291055.
- ^ Jones ME, Lucas PW, Tucker AS, Watson AP, Sertich JJ, Foster JR, et al. (June 2018). "Neutron scanning reveals unexpected complexity in the enamel thickness of an herbivorous Jurassic reptile". Journal of the Royal Society, Interface. 15 (143): 20180039. doi:10.1098/rsif.2018.0039. PMC 6030635. PMID 29899156.
- ^ Rauhut OW, Heyng AM, López-Arbarello A, Hecker A (2012). Farke AA (ed.). "A new rhynchocephalian from the late jurassic of Germany with a dentition that is unique amongst tetrapods". PLOS ONE. 7 (10): e46839. Bibcode:2012PLoSO...746839R. doi:10.1371/journal.pone.0046839. PMC 3485277. PMID 23118861.
- ^ Folinsbee, Kaila E.; Müller, Johannes; Reisz, Robert R. (2007-06-12). "Canine grooves: morphology, function, and relevance to venom". Journal of Vertebrate Paleontology. 27 (2): 547–551. doi:10.1671/0272-4634(2007)27[547:CGMFAR]2.0.CO;2. ISSN 0272-4634.
- ^ Folinsbee, Kaila E.; Müller, Johannes; Reisz, Robert R. (2007-06-12). "Canine grooves: morphology, function, and relevance to venom". Journal of Vertebrate Paleontology. 27 (2): 547–551. doi:10.1671/0272-4634(2007)27[547:CGMFAR]2.0.CO;2. ISSN 0272-4634.
- ^ Chambi-Trowell SA, Whiteside DI, Benton MJ, Rayfield EJ (November 2020). Lautenschlager S (ed.). "Biomechanical properties of the jaws of two species of Clevosaurus and a reanalysis of rhynchocephalian dentary morphospace". Palaeontology. 63 (6): 919–939. Bibcode:2020Palgy..63..919C. doi:10.1111/pala.12493. S2CID 220902843.
- ^ Herrera-Flores JA, Elsler A, Stubbs TL, Benton MJ (2021). "Slow and fast evolutionary rates in the history of lepidosaurs". Palaeontology. 65. doi:10.1111/pala.12579. ISSN 1475-4983. S2CID 244019684.
- ^ Jones ME (2006). "The Early Jurassic clevosaurs from China (Diapsida: Lepidosauria)". New Mexico Museum of Natural History and Science Bulletin. 37: 548–562.
- ^ Cleary TJ, Benson RB, Evans SE, Barrett PM (March 2018). "Lepidosaurian diversity in the Mesozoic-Palaeogene: the potential roles of sampling biases and environmental drivers". Royal Society Open Science. 5 (3): 171830. Bibcode:2018RSOS....571830C. doi:10.1098/rsos.171830. PMC 5882712. PMID 29657788.
- ^ a b Apesteguía S, Daza JD, Simões TR, Rage JC (September 2016). "The first iguanian lizard from the Mesozoic of Africa". Royal Society Open Science. 3 (9): 160462. Bibcode:2016RSOS....360462A. doi:10.1098/rsos.160462. PMC 5043327. PMID 27703708.
- ^ Simões TR, Wilner E, Caldwell MW, Weinschütz LC, Kellner AW (August 2015). "A stem acrodontan lizard in the Cretaceous of Brazil revises early lizard evolution in Gondwana". Nature Communications. 6 (1): 8149. Bibcode:2015NatCo...6.8149S. doi:10.1038/ncomms9149. PMC 4560825. PMID 26306778.
- ^ a b Jones ME, Tennyson AJ, Worthy JP, Evans SE, Worthy TH (April 2009). "A sphenodontine (Rhynchocephalia) from the Miocene of New Zealand and palaeobiogeography of the tuatara (Sphenodon)". Proceedings of the Royal Society B: Biological Sciences. 276 (1660): 1385–90. doi:10.1098/rspb.2008.1785. PMC 2660973. PMID 19203920.
- ^ Agnolín, Federico L.; Aranciaga Rolando, A. Mauro; Chimento, Nicolás R.; Novas, Fernando E. (October 2023). "New small reptile remains from the Late Cretaceous of Patagonia increase morphological diversity of sphenodontids (Lepidosauria)". Proceedings of the Geologists' Association. doi:10.1016/j.pgeola.2023.09.007.
- ^ Anantharaman, S.; DeMar, David G.; Sivakumar, R.; Dassarma, Dilip Chandra; Wilson Mantilla, Gregory P.; Wilson Mantilla, Jeffrey A. (2022-06-30). "First rhynchocephalian (Reptilia, Lepidosauria) from the Cretaceous–Paleogene of India". Journal of Vertebrate Paleontology. 42 (1): e2118059. Bibcode:2022JVPal..42E8059A. doi:10.1080/02724634.2022.2118059. ISSN 0272-4634. S2CID 252558728.
- ^ Apesteguía S, Gómez RO, Rougier GW (October 2014). "The youngest South American rhynchocephalian, a survivor of the K/Pg extinction". Proceedings of the Royal Society B: Biological Sciences. 281 (1792): 20140811. doi:10.1098/rspb.2014.0811. PMC 4150314. PMID 25143041.
더보기
- Apesteguia S, Rougier GW (2007). "A Late Campanian Sphenodontid Maxilla from Northern Patagonia" (PDF). American Museum Novitates (3581): 1–12. doi:10.1206/0003-0082(2007)3581[1:ALCSMF]2.0.CO;2. Retrieved 2019-03-30.
- Daugherty CH, Cree A, Hay JM, Thompson MB (1990). "Neglected taxonomy and continuing extinctions of tuatara (Sphenodon)". Nature. 347 (6289): 177–179. Bibcode:1990Natur.347..177D. doi:10.1038/347177a0. S2CID 4342765.
- Evans SE (November 2003). "At the feet of the dinosaurs: the early history and radiation of lizards" (PDF). Biological Reviews of the Cambridge Philosophical Society. 78 (4): 513–51. doi:10.1017/S1464793103006134. PMID 14700390. S2CID 4845536.
- Gemmell NJ, Rutherford K, Prost S, Tollis M, Winter D, Macey JR, et al. (August 2020). "The tuatara genome reveals ancient features of amniote evolution". Nature. 584 (7821): 403–409. doi:10.1038/s41586-020-2561-9. PMC 7116210. PMID 32760000.
외부 링크
 위키미디어 커먼즈의 스페노돈티아 관련 미디어
위키미디어 커먼즈의 스페노돈티아 관련 미디어 Wikis종의 Rynchocephalia 관련 자료
Wikis종의 Rynchocephalia 관련 자료
| ||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||


