소뇌의 해부학
Anatomy of the cerebellum| 소뇌 | |
|---|---|
 인간의 뇌를 그려 소뇌와 종아리 표시 | |
 | |
| 세부 사항 | |
| 일부 | 메트뇌론 |
| 동맥 | SCA, AICA, PICA |
| 정맥 | 우열 |
| 식별자 | |
| NeuroLex ID | birnlex_1489 |
| TA98 | A14.1.07.001 |
| TA2 | 5788 |
| 신경해부술의 해부학적 용어 | |
소뇌의 해부도는 3단계로 볼 수 있다.전체 해부학 수준에서, 소뇌는 단단히 접혀지고 구겨진 피질 층으로 구성되어 있으며, 그 아래에는 백질, 백질 속에 여러 개의 깊은 핵이 내장되어 있고,[1] 가운데에는 액체로 채워진 심실이 있습니다.중간 수준에서 소뇌와 그 보조 구조는 수백, [1]수천 개의 독립적으로 기능하는 모듈 또는 마이크로존으로 알려진 구획으로 분해될 수 있다.미시적 수준에서, 각 모듈은 매우 정형화된 [2]기하학으로 배열된 동일한 일련의 신경 요소들로 구성됩니다.
그로스 해부학
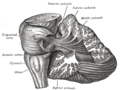
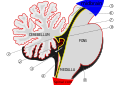
소뇌는 대뇌피질의 큰 덩어리와 그 앞에 있는 뇌간 부분의 앞부분에 위치하고 있다.그것은 단단한 경막의 층에 의해 위에 있는 대뇌와 분리되어 있다; 뇌의 다른 부분과의 모든 연결은 대뇌를 통해 이동한다.해부학자들은 소뇌를 종아리도 포함하는 중뇌의 일부로 분류한다; 중뇌는 차례로 마름모꼴 또는 "힌드브레인"의 윗부분이다.대뇌피질처럼, 소뇌는 두 개의 반구로 나뉘어져 있다; 또한 대뇌피질이라고 불리는 좁은 중간선 영역을 포함하고 있다.전체 구조를 10개의 작은 소엽으로 분할하기 위해 한 쌍의 큰 주름이 전통적으로 사용된다.
소뇌는 많은 수의 작은 과립 세포 때문에 뇌의 나머지 부분을 합친 것보다 더 많은 뉴런을 포함하고 있지만, 전체 [3]뇌 부피의 10%만을 차지한다.소뇌는 거의 2억 개의 입력 섬유를 받는다. 반면 시신경은 단지 백만 개의 섬유로 구성되어 있다.
소뇌의 특이한 표면 외관은 구조의 대부분이 매우 단단하게 접힌 회색 물질 층인 소뇌 피질로 이루어져 있다는 사실을 숨깁니다.인간의 소뇌피질이 완전히 펼쳐질 수 있다면 길이 약 1m, 폭 10cm의 신경조직 층이 생겨날 것으로 추정되며, 총 표면적은 500-1000평방센티미터이며, 모두 100-150입방센티미터의 [4][5]부피로 채워져 있다.대뇌피질의 회백질 아래에는 주로 대뇌피질을 오가는 골수 신경 섬유로 구성된 백색 물질이 있다.가지가 있고 나무와 같은 외관 때문에 소뇌에서 수목(arbor vitae)이라고 불리기도 하는 이 백색 물질 안에는 4개의 깊은 소뇌 핵이 포함되어 있습니다.
소뇌는 세 가지 기준에 따라 나눌 수 있습니다: 총 해부학, 계통학, 기능학.
총 해부학적 분할
전체 검사에서, 세 개의 엽을 소뇌에서 구별할 수 있습니다: 응집결절엽, 전엽('1차 균열'에 대한 동토층), 후엽('1차 균열'에 대한 동토층).후자는 중간선 소뇌와 외측 소뇌 반구로 더 나눌 수 있다.


계통발생 및 기능분할
소뇌는 또한 계통발생학적 기준(각 부위의 진화 연령)과 기능적 기준(각 부위가 가지고 있는 들어오고 나가는 연결과 정상적인 소뇌 기능에서의 역할)에 따라 세 부분으로 나눌 수 있다.계통학적으로 가장 오래된 것부터 최신 것까지 세 가지 부분이 있습니다.
| 기능적 명칭(유전학적 명칭) | 해부학적 부분 | 역할. |
| 전정맥류 (아키세레벨룸) | 응집결절엽(및 바로 인접한 대뇌) | 전정맥은 균형과 눈의 움직임을 조절합니다.그것은 반고리관과 전정핵 양쪽에서 전정입력을 받아 섬유질을 다시 내측과 외측 전정핵으로 보낸다.그것은 또한 상위 콜로쿨리와 시각 피질(후자는 피질-폰토-세레벨 경로를 형성하며 폰틴 핵을 통해)로부터 시각적 입력을 받는다.전정맥의 병변은 균형과 보행에 지장을 준다.바이벤터 소엽으로 알려진 또 다른 작은 지역이 있습니다. |
| 스피노세레벨룸 (대퇴골) | 반구의 버미스 및 중간부("파라버미스") | 척추골은 몸과 사지의 움직임을 조절한다.그것은 시각 및 청각 시스템뿐만 아니라 척수의 등쪽 기둥(척추신경 포함)과 삼차 신경으로부터 고유 수용 입력을 받습니다.그것은 다시 대뇌 피질(중뇌와 시상)과 뇌간(지골의 망상 형성, 그리고 수질의 전정핵을 통해)에 섬유질을 투사하여 하강 운동 시스템의 변조를 제공한다.척추신경종은 우주에서 다양한 신체 부위의 위치에 대한 데이터를 받을 때 감각 지도를 포함합니다: 특히, 해충은 몸통과 사지 근위부로부터 섬유질을 받는 반면, 반구의 중간 부분은 사지 원위부로부터 섬유질을 받습니다.척추신경종은 움직임의 과정 동안 신체 부위의 미래 위치를 예측하기 위해 "피드 포워드" 방식으로 고유 수용적 입력을 정교하게 할 수 있습니다. |
| 뇌뇌 (네오세레벨룸, 폰토세레벨룸) | 반구의 외측부 | 네오세레벨룸은 움직임을 계획하고 행동을 위한 감각 정보를 평가하는 데 관여합니다.대뇌피질(특히 두정엽)에서 폰틴핵(대뇌에서 피질-전뇌-전뇌 경로 형성)과 치핵(소뇌에서)을 통해 단독으로 입력을 받아 주로 복측 시상(이후 전뇌피질 및 1차 운동 영역에 연결된 섬유)을 보낸다.대뇌피질)과 적색핵(소뇌 반구와 다시 연결되는 하감람핵에 연결됨).네오세레벨룸은 곧 일어날[6] 계획운동에 관여하며 순수하게 인지기능도 가지고 있다. |
소뇌의 기능에 대해 이해되는 것의 대부분은 부상이나 질병으로 고통받거나 동물 병변 연구를 통해 인간 환자의 초점 병변의 영향에 대한 세심한 기록에서 비롯된다.
세포 해부학
기능 섹션에서 더 자세히 설명했듯이, 소뇌는 신경 신호의 흐름이 거의 완전히 단방향이라는 점에서 대부분의 다른 뇌 영역과 다릅니다. 즉, 소뇌의 신경 요소들 사이에는 사실상 역방향 연결이 없습니다.따라서 셀룰러 구조를 설명하는 가장 논리적인 방법은 입력에서 시작하여 출력까지의 연결 순서를 따르는 것입니다.
심층핵
소뇌의 네 개의 깊은 핵은 상아핵, 색전형핵, 구상핵, 그리고 fastigii핵으로 뇌의 특정 부분과 정보를 주고받으며 의사소통의 주요 중추 역할을 합니다.게다가, 이 핵들은 뇌의 다른 부분들로부터 억제 신호와 흥분 신호를 모두 받고, 이것은 차례로 핵의 발신 [7]신호에 영향을 미친다.(구형핵과 색전상핵은 삽입된 핵을 구성합니다.)
피질층


소뇌의 세포 구조(세포 조직)는 매우 균일하며, 연결이 수직 회로 소자의 대략적인 3차원 배열로 구성되어 있습니다.이러한 조직의 통일성은 신경 회로를 비교적 쉽게 연구할 수 있게 합니다.
소뇌 피질에는 세 개의 층이 있습니다; 바깥쪽에서 안쪽 층까지, 이것들은 분자층, 푸르키네, 그리고 입상층입니다.소뇌피질의 기능은 본질적으로 깊은 핵을 통해 흐르는 정보를 조절하는 것이다.소뇌의 미세 회로는 그림 5에 도식화되어 있다.이끼 낀 섬유와 기어오르는 섬유는 감각 운동 정보를 깊은 핵으로 전달하고, 이는 다시 다양한 운동 전 영역에 전달하여 운동 활동의 이득과 시기를 조절합니다.이끼와 기어오르는 섬유는 또한 이 정보를 소뇌 피질에 공급하고, 소뇌 피질은 다양한 계산을 수행하여 푸르키네 세포 발화를 조절합니다.Purkinje 뉴런은 강력한 억제 시냅스를 통해 깊은 핵으로 피드백합니다.이 시냅스는 이끼 낀 섬유와 기어오르는 섬유들이 깊은 핵을 활성화하는 정도를 조절하고, 따라서 소뇌가 운동 기능에 미치는 궁극적인 영향을 조절합니다.소뇌피질에 있는 거의 모든 시냅스의 시냅스 강도는 시냅스 가소성을 겪는 것으로 나타났다.이것은 소뇌 피질의 회로가 소뇌의 출력을 지속적으로 조정하고 미세 조정할 수 있도록 하여, 일부 유형의 운동 학습과 조정의 기초를 형성합니다.소뇌 피질의 각 층은 이 회로를 구성하는 다양한 세포 유형을 포함한다.
분자층
이 소뇌 피질의 가장 바깥쪽 층은 두 가지 종류의 억제성 인터요론을 포함합니다: 성질과 바구니 세포.또한 Purkinje 뉴런의 수지상 결정체와 과립 세포로부터의 평행 섬유 조직을 포함합니다.성질과 바스켓 세포 모두 푸르키네 세포 수상돌기에 GABAergic 시냅스를 형성합니다.
푸르키네층
중간층은 오직 한 종류의 세포 몸체를 포함하고 있는데, 그것은 큰 푸르키네 세포이다.Purkinje 세포는 소뇌 피질의 1차 통합 뉴런이며 소뇌 피질의 유일한 출력을 제공합니다.Purkinje 세포 수상돌기는 분자층까지 도달하는 수백 개의 가시가 있는 큰 나무입니다(그림 6).이 수상목재들은 거의 모든 것이 평면에 놓여 있으며, 인접한 푸르키네 나무들이 평행 평면에 놓여 있다.과립 세포에서 나오는 각 평행 섬유는 많은 층을 통과하는 와이어처럼 이러한 나무들을 통해 직교로 흐릅니다.Purkinje 뉴런은 GABAergic(억제성 시냅스를 갖는다는 뜻)으로 뇌간에 있는 깊은 소뇌와 전정핵의 뉴런을 가지고 있습니다.각 Purkinje 세포는 10만~20만 개의 평행섬유로부터 흥분성 입력을 받는다.평행 섬유는 Purkinje 세포의 단순(전부 또는 무, 진폭 불변) 스파이크의 원인이 된다고 합니다.
푸르키네 세포는 또한 등반섬유를 통해 하감람핵으로부터 입력을 받는다.이 상호작용에 대한 좋은 기억은 등반 섬유가 반대쪽 하등 올리브에서 유래한다는 것을 고려할 때 "다른 올리브 나무를 기어오른다"는 문구이다.평행 섬유로부터의 10만 개 이상의 입력과는 대조적으로, 각각의 Purkinje 세포는 정확히 하나의 상승 섬유로부터 입력을 받습니다; 그러나 이 단일 섬유는 Purkinje 세포의 수상돌기를 "클립"하고, 그것들을 감아 많은 시냅스를 만듭니다.순입력이 너무 강해서 등반 섬유로부터의 단일 활동 전위는 푸르키네 세포에서 "복잡한 스파이크"를 생성할 수 있습니다. 즉,[8] 진폭 감소와 함께 연속적으로 여러 스파이크가 폭발하고, 그 후 단순한 스파이크가 억제되는 동안 멈춥니다.
Purkinje 층 바로 아래에는 매우 긴 수상돌기가 Purkinje 층과 입상 층 사이의 경계를 따라 이동하는 Lugaro 세포가 있습니다.
입상층
가장 안쪽에 있는 층은 세 종류의 세포로 이루어진 세포 본체를 포함합니다: 수 많은 작은 과립 세포, 약간 더 큰 단극성 붓 세포[9], 그리고 훨씬 더 큰 골지 세포.이끼가 낀 섬유는 그들의 주요 원점인 폰틴 핵에서 입상층으로 들어간다.이 섬유들은 과립 세포와 깊은 소뇌 핵의 세포와 흥분성 시냅스를 형성합니다.과립 세포는 평행 섬유로 알려진 T자형 축삭을 표면 분자층으로 보내 푸르키네 세포 수상돌기와 수십만 개의 시냅스를 형성합니다.인간의 소뇌는 600억에서 800억 개의 과립 세포를 포함하고 있으며, 이 단일 세포 유형을 뇌에서 단연코 가장 많은 뉴런으로 만듭니다.골지 세포는 과립 세포에 억제 피드백을 제공하며, 과립 세포와 시냅스를 형성하고 축삭을 분자 층에 투영합니다.
대뇌피질과의 관계
신피질과 소뇌의 국소장 전위는 깨어 있는 [10]동물에서 (6-40Hz)로 일관되게 진동한다.이것들은 대뇌피질에서 [11]나오는 출력의 통제하에 있는 것으로 보인다.이 출력은 신피질의 5/6층 뉴런에서 이 프로젝트를 통해 종아리나 하등 올리브로 가는 경로에 의해 조정됩니다.핀을 통해 과립세포와 골지뉴런과 시냅스하는 이끼성 섬유로 이동하고 흥분성 평행섬유를 통해 푸르키네뉴런을 목표로 합니다.만약 하등 올리브라면 흥분성 섬유질 주입을 통해 푸르키네 [11]뉴런으로 보내질 것이다.이것들은 이 출력을 복측외측 시상부를 통해 대뇌피질로 돌려보내 루프를 완성시킵니다.
소뇌와 관련된 가장 큰 경로는 피질세뇌경로이다.대뇌피질에서 발생하는 이 섬유들은 먼저 폰틴핵에서 일방적으로 종단된다.그리고 나서 섬유는 갈라져 중간 소뇌를 형성하고 소뇌 피질에서 이끼가 낀 섬유로 끝납니다.이 경로는 진행 중인 움직임과 다가오는 움직임에 대해 소뇌에 알리는 신호를 전달합니다.이는 모터 [12]활동을 지속적으로 조정하는 데 도움이 됩니다.
이 운동의 시작은 피질영양뇌 경로를 통해 소뇌로 전달됩니다.그 시냅스들은 망상형성 중에 일방적으로 하부와 중부를 거쳐 [12]소뇌로 들어간다.
운동피질과 체질감각피질은 편측 하방 및 부속 감람핵으로 돌출되어 올리보세뇌관을 형성합니다.하감람핵에서 양쪽으로 피질-올리브 섬유가 시냅스한다.그 질서는 반대쪽 소뇌 피질의 '체 지도'에 있는 올리보 세레벨로 돌출부에 보존된다.동물실험의 정지상태에서 올리비뉴런 그룹은 5~10Hz(충동/초)로 동시에 방출된다.소뇌피질에서, 푸르키네 세포의 반응은 복잡한 [13]스파이크의 형태를 취한다.
소뇌는 소뇌 시상관을 통해 대뇌피질로 돌기를 돌려보낸다.
소뇌 측면 확장 또는 신뇌는 인지 기능과 관련될 수 있으며 해부학적으로 측면 전전두피질과 관련이 있다.이는 음성 중 가장 큰 활동을 나타내며, (시상을 통해) 운동 음성 [13]영역과의 가능한 연결과 일치하는 일방적 우세를 보인다.
피질성 세뇌 섬유에 의해 소뇌와 연결된 결합 영역에 병변이 발생하면 인지 정서 증후군이 발생할 수 있습니다.이것은 감소된 추리력, 부주의, 말의 문법적 오류, 빈약한 공간 감각, 그리고 부분적인 기억 [13]상실의 형태로 인지 결함을 초래한다.
혈액 공급


상소뇌동맥(SCA), 전방하소뇌동맥(AICA), 후방하소뇌동맥(PICA) 등 3개의 동맥이 소뇌에 혈액을 공급한다(그림 7).
SCA는 후대뇌동맥으로 분기되는 부분보다 약간 낮은 기저동맥의 외측부로 분기된다.여기서, 그것은 소뇌에 도달하기 전에 뒤쪽으로 종아리 주변을 감싼다.SCA는 대부분의 소뇌피질, 소뇌핵, 그리고 상위 [14]소뇌페달에 혈액을 공급한다.
AICA는 척추 동맥의 접합부 바로 위에 있는 기저 동맥의 측면 부분을 분리한다.그것의 기원에서, 그것은 소뇌에 도달하기 전에 소뇌의 각도에서 종아리 아랫부분을 따라 분기합니다.이 동맥은 하소뇌의 앞부분인 중소뇌와 안면신경(CN VII)과 전정신경(CN VII)에 혈액을 공급한다.AICA의 폐색은 얼굴의 마비, 마비, 감각 상실을 일으킬 수 있으며 청각 장애를 일으킬 수도 있다.또한 소뇌 각도의 경색을 일으킬 수 있습니다.이것은 과판증(등골근육의 기능, CN VII에 의해 내감됨)과 현기증(전정 반원관의 내림프 가속에서 CN VII의 변화로 인한 잘못된 해석)을 초래할 수 있다.
PICA는 척수동맥의 외측부분과 기저동맥과의 접합부 바로 아래쪽으로 갈라진다.소뇌의 하부 표면에 도달하기 전에, PICA는 여러 개의 뇌 신경 핵에 혈액을 공급하면서 수질에 가지를 보낸다.소뇌에서 PICA는 소뇌의 후하부, 하소뇌 족저핵, 모호한핵, 미주신동핵, 척수삼차핵, 단독핵 및 전정핵에 혈액을 공급한다.
척추동물의 변이
척추동물의 종류에 따라 소뇌의 크기와 모양에 상당한 차이가 있다.그것은 일반적으로 연골과 뼈 있는 물고기, 새, 포유류에서 가장 크지만 파충류에서는 다소 작다.사람에게서 발견되는 커다란 쌍으로 된 복잡한 엽은 전형적인 포유동물이지만, 소뇌는 일반적으로 다른 집단에서 하나의 중앙 엽이며, 매끄럽거나 약간 홈이 나 있다.포유동물에서 신뇌는 질량에 의해 소뇌의 주요 부분이지만, 다른 척추동물에서는 전형적으로 척추신경이다.[15]
양서류, 칠성장어, 먹장어에서는 소뇌가 거의 발달하지 않았습니다. 후자의 두 그룹에서는 소뇌가 뇌간과 거의 구별되지 않습니다.척수돌기는 이러한 그룹에 존재하지만, 일차 구조는 전정맥에 [15]대응하는 작은 쌍의 핵이다.
소아과
소뇌는 [16]해부학에서 볼 수 있는 일반적인 3가지 패턴을 따르며, 3가지 주요 입력 및 출력 소뇌 다발(섬유 다발)을 가지고 있습니다.이것들은 상(상완결막), 중(상완골), 하(하완골 및 대퇴골) 소뇌 족저이다.
| 페둔클 | 묘사 |
| 우월한 | 전척추신경로에서 나오는 구심성 섬유는 이 페둔클을 통해 전소뇌엽으로 전달되지만, 대부분의 섬유는 유출성 섬유이다.따라서, 상위 소뇌 육교는 소뇌의 주요 출력 경로이다.대부분의 섬유는 치핵 내에서 발생하며, 치핵은 다시 붉은 핵, 시상 복측/복측 전방 핵 및 수질을 포함한 다양한 중뇌 구조에 투영됩니다.치아의 치핵 > 적색핵 > 시상전피질(치핵 > 시상전피질)과 소뇌 시상피질(세뇌 > 시상전피질)은 이 페둔클을 통과하는 두 가지 주요 경로로 운동 계획에 중요하다. |
| 가운데 | 이것은 거대한 피질핵(대뇌피질 > pons > 소뇌)의 일부로서 폰틴핵에서 유래한 구심성 섬유로 전적으로 구성되어 있다.이 섬유들은 대뇌 신피질의 감각과 운동 영역에서 내려와 중간 소뇌를 세 개의 소뇌 중 가장 큰 것으로 만듭니다. |
| 열등한 | 이것은 주로 균형 및 자세 유지와 같은 운동 전정 기능과 고유 수용성 감각 입력을 통합하는 것과 관련된 많은 유형의 입력 및 출력 섬유를 운반합니다.몸으로부터의 자체 수용 정보는 배측척추신경로를 통해 소뇌로 운반된다.이 관은 하부 소뇌를 통과하여 고생뇌 내의 시냅스를 통과한다.전정 정보가 대뇌동맥에 투영됩니다. 하등 올리브의 등반 섬유는 하등 소뇌를 통과한다. 이 페둔클은 또한 Purkinje 세포에서 직접 정보를 Pons와 medulla 사이의 접합부에 위치한 등쪽 뇌간 전정핵으로 전달합니다. |
소뇌에는 각각 이끼가 낀 섬유와 기어오르는 섬유로 구성된 두 가지 범주로 세 가지 입력 소스가 있습니다.이끼성 섬유는 대뇌피질에서 정보를 운반하는 뇌엽에 위치한 뉴런 클러스터인 폰틴 핵에서 유래할 수 있습니다.그들은 또한 측척수에서 유래한 척추신경관 내에서 발생할 수 있다.소뇌에서 나오는 대부분의 분비물은 세 개의 족부를 통해 나오기 전에 처음에는 깊은 소뇌 핵으로 시냅스된다.가장 주목할 만한 예외는 Purkinje 세포에 의한 전정핵의 직접적인 억제이다.
발전
배아 발달의 초기 단계 동안, 뇌는 세 개의 뚜렷한 부분으로 형성되기 시작합니다: 프로센스팔론, 중간뇌, 그리고 마름뇌.마름뇌는 배아 뇌의 가장 꼬리가 있는 부분이며, 소뇌가 발달하는 부분은 이 부분입니다.태아의 마름모꼴 부분을 따라 마름모꼴이라고 불리는 8개의 붓기가 생긴다.소뇌는 결국 뇌와 척수를 형성하는 구조인 신경관의 경음판에 위치한 두 개의 마름모세포에서 발생한다.소뇌가 형성되는 특정한 마름모세포는 꼬리 근처인 마름모세포 1(Rh.1)과 앞 [17]근처인 복강근이다.
두 개의 주요 부위가 소뇌를 구성하는 뉴런을 발생시키는 것으로 생각된다.첫 번째 부위는 제4심실 지붕에 있는 심실 영역입니다.이 지역은 푸르키네 세포와 깊은 소뇌 핵 뉴런을 생산한다.이 세포들은 소뇌피질과 소뇌의 주요 출력 뉴런이다.두 번째 생식대(세포 출생지)는 마름모꼴 입술로 알려져 있으며, 뉴런은 인간 배아 27주차에 외부 입상층으로 이동한다.소뇌의 바깥쪽에 있는 이 세포 층은 과립 뉴런을 생산합니다.과립 뉴런은 이 외부 층에서 이동하여 내부 과립 [18]층으로 알려진 내부 층을 형성합니다.외부 과립층은 성숙한 소뇌에서 더 이상 존재하지 않고 내부 과립층에 과립세포만 남습니다.소뇌의 백색 물질은 소뇌의 세 번째 생식 영역일 수 있지만, 생식 영역으로서의 기능은 논란의 여지가 있다.
기타 이미지
소뇌의 투사 섬유를 나타내는 절개
제4심실 지붕 구조입니다.화살은 마젠디의 구멍에 있다.

인간의 뇌 정중 시상도
주요 랜드마크를 나타내는 숫자가 있는 인간 소뇌의 정면도




레퍼런스
- ^ a b Knierim, James. "Chapter 5: Cerebellum". Neuroscience Online: An Electronic Textbook for the Neurosciences.
- ^ Friede, Reinhard L. (1973-03-01). "Dating the development of human cerebellum". Acta Neuropathologica. 23 (1): 48–58. doi:10.1007/BF00689004. ISSN 1432-0533. PMID 4698523. S2CID 5387374.
- ^ 머리부터 발끝까지
- ^ Edwards CR, Newman S, Bismark A, et al. (2008). "Cerebellum volume and eyeblink conditioning in schizophrenia". Psychiatry Res. 162 (3): 185–194. doi:10.1016/j.pscychresns.2007.06.001. PMC 2366060. PMID 18222655.
- ^ Hutchinson S, Lee LH, Gaab N, Schlaug G (2003). "Cerebellar volume of musicians". Cereb. Cortex. 13 (9): 943–9. doi:10.1093/cercor/13.9.943. PMID 12902393.
- ^ Kingsley, RE (2000). Concise Text of Neuroscience (2nd ed.). Lippincott Williams and Wilkins. ISBN 0-683-30460-7.
- ^ Harting, J.K. "The Global Cerebellum '97". University of Wisconsin Medical School.
- ^ Häusser, Michael; Clark, Beverley A.; Davie, Jenny T. (2008-07-23). "The origin of the complex spike in cerebellar Purkinje cells". Journal of Neuroscience. 28 (30): 7599–7609. doi:10.1523/JNEUROSCI.0559-08.2008. ISSN 0270-6474. PMC 2730632. PMID 18650337.
- ^ Kinney GA, Overstreet LS, Slater NT (September 1997). "Prolonged physiological entrapment of glutamate in the synaptic cleft of cerebellar unipolar brush cells" (PDF). J Neurophysiol. 78 (3): 1320–33. doi:10.1152/jn.1997.78.3.1320. PMID 9310423.
- ^ Soteropoulos DS, Baker SN (2006). "Cortico-cerebellar coherence during a precision grip task in the monkey". J Neurophysiol. 95 (2): 1194–206. doi:10.1152/jn.00935.2005. PMID 16424458.
- ^ a b Ros H, Sachdev RN, Yu Y, Sestan N, McCormick DA (2009). "Neocortical networks entrain neuronal circuits in cerebellar cortex". Journal of Neuroscience. 29 (33): 10309–20. doi:10.1523/JNEUROSCI.2327-09.2009. PMC 3137973. PMID 19692605.
- ^ a b Gartner, Leslie P.; Patestas, Maria A. (2009). Textbook of Neuroanatomy. Wiley-Blackwell. p. 464. ISBN 9781405103404.
- ^ a b c Mtui, Estomih; Gruener, Gregory; Dockery, Peter (2016). Fitzgerald's Clinical Neuroanatomy and Neuroscience (7th ed.). Elsevier. pp. 243–252.
- ^ Gray, Henry; Lewis, Warren Harmon (1918). Anatomy of the human body (20th ed.). Philadelphia: Lea & Febiger.
- ^ a b Romer, Alfred Sherwood; Parsons, Thomas S. (1977). The Vertebrate Body. Philadelphia, PA: Holt-Saunders International. p. 531. ISBN 0-03-910284-X.
- ^ "List of Three's". www.meddean.luc.edu.
- ^ Muller F, O'Rahilly R (1990). "The human brain at stages 21–23, with particular reference to the cerebral cortical plate and to the development of the cerebellum". Anat Embryol (Berl). 182 (4): 375–400. doi:10.1007/BF02433497. PMID 2252222. S2CID 33485509.
- ^ Smeyne, Richard J.; Goldowitz, Dan (May 1989). "Development and death of external granular layer cells in the weaver mouse cerebellum: a quantitative study". The Journal of Neuroscience. 9 (5): 1608–20. doi:10.1523/JNEUROSCI.09-05-01608.1989. PMC 6569844. PMID 2723742.


