두팔로포드 눈
Cephalopod eye
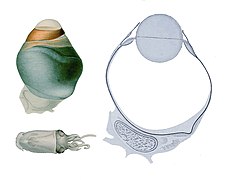
두팔로포드(Cephalopods)는, 활동적인 해양 포식자로서, 수생 환경에서 사용하기에 특화된 감각 기관을 가지고 있다.[1] 그들은 홍채, 원형 렌즈, 유리강(아이젤), 색소 세포, 빛에 민감한 망막에서 빛을 뇌로 전달해주는 광수용체 세포로 구성된 카메라 형태의 눈을 가지고 있다.[2] 지난 140년 동안 카메라 형태의 두엽수 눈은 수렴 진화의 예로서 척추동물 눈과 비교되어 왔는데, 두 가지 유형의 유기체가 카메라-눈 특성을 독립적으로 진화시켰으며 두 가지 모두 유사한 기능을 공유한다. 이것이 진정으로 수렴 진화인지 아니면 평행 진화인지에 대한 논쟁이 존재한다.[3] 척추동물 카메라 눈과는 달리, 두족류들은 신체 표면의 침입으로서(뇌의 밖으로 자라지 않고), 결과적으로 각막이 눈의 구조적인 부분과는 반대로 눈 위에 놓여 있다.[4] 척추동물 눈과는 달리, 두족류 눈은 사람의 눈 속의 렌즈가 하는 것처럼 모양을 바꾸는 것보다 카메라나 망원경의 렌즈와 같이 움직임을 통해 집중된다. 눈은 렌즈와 마찬가지로 대략적으로 구면이며, 완전히 내부에 있다.[5]
세팔로포드의 눈은 망막의 뒷면을 통과하는 망막축이 있을 정도로 발달하므로 시신경은 눈을 빠져나오기 위해 광수용체층을 통과할 필요가 없고 척추동물의 자연적이고 중심적이며 생리적인 맹점을 갖지 않는다.[6]
렌즈에 사용된 크리스탈린은 척추동물 크리스탈린과는 독립적으로 발달한 것으로 보이며, 이는 렌즈의 동종 원점을 암시한다.[7]
대부분의 두족류들은 눈의 총 위치를 매우 미세하게 조절할 수 있는 복잡한 추가 안구 근육 시스템을 가지고 있다. 문어는 항상 수평이 되도록 동공 방향을 유지하는 자율적인 반응을 가지고 있다.[1]
편광
오징어와 문어, 그리고 잠재적으로 오징어와 오징어 등 여러 종류의 두족류들이 편광의 방향을 구별할 수 있는 눈을 가지고 있다는 것이 기록되어 있다. 이 민감도는 이웃한 광수용체의 직교조직에 기인한다.(세팔로포드는 다른 연체동물과 유사한 rhabdoms라고 불리는 수용세포를 가지고 있다.) 예를 들어, 척추동물의 눈은 로드와 콘의 시각적 색소가 반 무작위로 배열되기 때문에 일반적으로 양극화 차이에 둔감하며, 따라서 빛의 전자 벡터 축 방향에도 동일하게 민감하다. 두팔로포드 눈의 시각적 색소 분자는 직교 조직으로 인해 가벼운 전자 벡터 축과 적절하게 정렬되었을 때 가장 높은 광 흡수를 하여 양극화 차이에 대한 민감성을 허용한다.[8] 이 능력의 정확한 기능은 입증되지 않았지만, 색깔을 바꾸는 두족류 사이의 먹이 탐지, 항법, 그리고 아마도 의사소통을 위한 것으로 가정되었다.[8][9]
Bathyteuthis sp의 눈.

문어(옥토퍼스속눈

갑오징어 눈

노틸러스 (Nautilus pompilius) 눈




.jpg)
진화론
두족류 내에서와 척추동물 내에서 카메라 눈의 진화가 병렬 진화인지 아니면 수렴 진화인지에 대한 의견의 불일치는 대부분 해결되었지만 여전히 존재한다. 현재의 상황은 그들의 유사한 카메라 타입의 눈을 위한 융합적 진화의 것이다.
평행 진화
평행진화라고 주장하는 사람들은 이 눈발달을 위한 유전정보를 담고 있는 공통된 조상이 있었다는 증거가 있다고 말한다. 이것은 눈발달을 위해 표현하는 유전자 Pax6를 포함하는 모든 다변측정감시 유기체에 의해 증명된다.[10]
수렴 진화
수렴 진화를 지지하는 사람들은 이 공통 조상이 두족류 동물과 척추동물 둘 다 상당한 차이로 앞서갔을 것이라고 말한다. 카메라형 눈의 표현을 가진 공통 조상은 두족류에서 카메라형 눈의 진화 전 약 2억 7천만 년, 척추동물에서 카메라형 눈의 진화 전 약 1억 1천만 년에서 2억 6천만 년 전에 존재했을 것이다.[11] 이에 대한 또 다른 근거는 두족류 동물과 척추동물 모두에서 발생하는 독립된 Pax6 변형으로 인한 표현의 차이다. 세팔로포드는 척추동물의 팍스6와 비교할 때 유사한 유전자 발현을 허용하지만, 독자적으로 발생하여 척추동물이 공유하지 않는 5가지 변종의 팍스6를 게놈에 포함한다.[12]
연구 및 의료용도
이 분야에서 대두되고 있는 주요 의학적 용도는 안구 발달과 안구 질환에 대한 연구용이다. 안구 유전자 발현에 대한 새로운 연구는 유사한 인간의 눈과 융합 진화의 증거 때문에 두족류 눈을 사용하여 수행되고 있다. 이 연구들은 비록 드로필라 연구가 가장 보편적인 것으로 남아있지만, 눈 발달 중 유전자 발현을 위한 이전의 드로필라 연구를 가장 정확한 것으로 대체한다. 그들이 유사하다는 결론은 두 가지 특성이 유사한 환경에서 유사한 압력에 의해 자연 선택을 통해 형성되었을 것이기 때문에, 두 유기체의 눈에 유사한 안구 질환의 발현이 있을 것이라는 것을 의미하기 때문에, 애초에 의학 사용에 대한 비교에 대한 신뢰도를 높인다.[2]
두팔로포드 눈 실험의 장점은 두팔로포드가 그들의 발달과정을 재활성화하는 능력 때문에 그들의 눈을 재생시킬 수 있다는 것인데, 이것은 질병의 효과를 연구할 때 동일한 두팔로포드에 대한 연구가 하나의 시험 샘플을 지나도록 계속할 수 있게 한다. 이것은 또한 재생이 두족류 게놈에 보존되는 방법과 그것이 카메라 눈을 위해 발현되는 유전자와 함께 인간 게놈에 어느 정도 보존될 수 있는지에 관한 보다 복잡한 연구를 가능하게 한다.[2]
참고 항목
참조
- ^ Jump up to: a b 부델만 BU. "세팔로포드는 장기, 신경, 뇌를 감지한다. 고성능과 라이프스타일을 위한 적응." 해양 및 담수 행동 및 생리학. 제25권, 제1-3호 13-33페이지.
- ^ Jump up to: a b c Serb, Jeanne M. (2008). "Toward Developing Models to Study the Disease, Ecology, and Evolution of the Eye in Mollusca*" (PDF). American Malacological Bulletin. 26 (1–2): 3–18. doi:10.4003/006.026.0202. S2CID 1557944. Archived from the original (PDF) on 2014-12-18. Retrieved 2014-11-18.
- ^ Serb, J.; Eernisse, D. (2008). "Charting Evolution's Trajectory: Using Molluscan Eye Diversity to Understand Parallel and Convergent Evolution". Evolution: Education & Outreach. 1 (4): 439–447. doi:10.1007/s12052-008-0084-1.
- ^ Hanke, Frederike D.; Kelber, Almut (2020-01-14). "The Eye of the Common Octopus (Octopus vulgaris)". Frontiers in Physiology. 10: 1637. doi:10.3389/fphys.2019.01637. ISSN 1664-042X. PMC 6971404. PMID 32009987.
- ^ Yamamoto, M. (Feb 1985). "Ontogeny of the visual system in the cuttlefish, Sepiella japonica. I. Morphological differentiation of the visual cell". The Journal of Comparative Neurology. 232 (3): 347–361. doi:10.1002/cne.902320307. ISSN 0021-9967. PMID 2857734. S2CID 24458056.
- ^ Gregory, T. Ryan (2008). "Introduction". Evolution: Education and Outreach. 1 (4): 352–354. doi:10.1007/s12052-008-0073-4. ISSN 1936-6426.
- ^ SAMIR K. BRAHMA1 (1978). "Ontogeny of lens crystallins in marine cephalopods" (PDF). Embryol. Exp. Morph. 46 (1): 111–118. PMID 359745.
- ^ Jump up to: a b Mathger, L.M.; Shashar, N.; Hanlon, R.T. (2009). "Do cephalopods communicate using polarized light reflections from their skin?". Journal of Experimental Biology. 212 (Pt 14): 2133–2140. doi:10.1242/jeb.020800. PMID 19561202.
- ^ Shashar, N; Rutledge, P; Cronin, T (1996). "Polarization vision in cuttlefish in a concealed communication channel?". Journal of Experimental Biology. 199 (9): 2077–2084. PMID 9319987.
- ^ Gehring, W. J. (2004). "Historical perspective on the development and evolution of eyes and photoreceptors". The International Journal of Developmental Biology. 48 (8–9): 707–717. doi:10.1387/ijdb.041900wg. PMID 15558463.
- ^ Fernald, Russell D. (29 September 2006). "Casting a genetic light on the evolution of eyes". Science. 313 (5795): 1914–1918. Bibcode:2006Sci...313.1914F. doi:10.1126/science.1127889. PMID 17008522. S2CID 84439732.
- ^ Yoshida, Masa-aki; Yura, Kei; Ogura, Atsushi (2014). "Cephalopod eye evolution was modulated by the acquisition of Pax-6 splicing variants". Scientific Reports. 4 (4256): 4256. Bibcode:2014NatSR...4E4256Y. doi:10.1038/srep04256. PMC 3942700. PMID 24594543.



| show 동물의 시각 |
|---|
.jpg)


