형광 현미경
Fluorescence microscope
형광현미경은 유기물 [1][2]또는 무기물의 특성을 연구하기 위해 산란, 반사, 감쇠 또는 흡수 대신 형광을 사용하는 광학현미경이다.형광현미경은 형광영상의 [3]분해능을 높이기 위해 광학단면을 사용하는 공초점 현미경 등 복잡한 디자인이든 형광을 이용해 이미지를 생성하는 현미경을 말한다.
원칙
시료를 형광구에 의해 흡수되는 특정 파장(또는 파장)의 빛으로 조명하여 더 긴 파장(즉, 흡수된 빛과 다른 색)의 빛을 방출한다.조명 빛은 스펙트럼 방출 필터를 사용하여 훨씬 약한 방출 형광으로부터 분리된다.형광 현미경의 일반적인 구성 요소는 광원(제논 아크 램프 또는 수은 증발 램프가 일반적입니다. 보다 발전된 형태는 고출력 LED 및 레이저), 여기 필터, 이분법 미러(또는 이분법 빔플리터) 및 발광 필터(아래 그림 참조)입니다.필터와 이색 빔플리터는 [1]시료에 라벨을 붙일 때 사용되는 형광체의 스펙트럼 들뜸 및 방출 특성과 일치하도록 선택되었다.이와 같이, 한 번에 하나의 불소(컬러)의 분포를 촬영한다.여러 유형의 형광 포자의 멀티 컬러 이미지는 여러 단색 이미지를 [1]결합하여 합성해야 합니다.
사용되는 대부분의 형광 현미경은 에피 형광 현미경이며, 형광체의 들뜸과 형광체의 검출은 동일한 광로를 통해 이루어진다(즉, 대상을 통해).이러한 현미경은 생물학에서 널리 사용되며 공초점 현미경 및 전체 내부 반사 형광 현미경(TIRF)과 같은 고급 현미경 설계의 기초가 됩니다.
에피 형광 현미경법

형광 현미경, 특히 생명과학에서 사용되는 현미경은 다이어그램에 표시된 에피 형광 설계입니다.여기 파장의 빛은 대물렌즈를 통해 시료를 조사한다.시료에 의해 방출된 형광은 여진에 사용되는 것과 동일한 목적에 의해 검출기에 집중되며, 더 높은 분해능을 위해서는 더 높은 수치 개구부를 가진 대물 렌즈가 필요하다.대부분의 들뜸광은 시료를 투과하기 때문에 반사 들뜸광만이 방출광과 함께 목표물에 도달하기 때문에 에피 형광법은 높은 신호 대 잡음비를 제공한다.이색 빔플리터는 파장 고유의 필터 역할을 하며 형광 빛을 접안렌즈 또는 검출기에 투과하지만 나머지 들뜸 빛은 소스로 반사됩니다.
광원
형광 현미경 검사는 할로겐 램프와 같은 일부 광원이 제공할 [4]수 없는 강렬한 단색에 가까운 조명이 필요합니다.제논 아크 램프 또는 여기 필터가 있는 수은-증기 램프, 레이저, 초점원, 고출력 LED를 포함한 4가지 주요 광원이 사용된다. 레이저들은 제논 램프와 함께 공초점 현미경법, 전체 내부 반사 형광 현미경과 같은 보다 복잡한 형광 현미경 기술에 가장 널리 사용된다.와이드필드 에피 형광 현미경에는 이색 들뜸 필터가 있는 LED 및 수은 램프가 일반적으로 사용됩니다.광시야 에피 형광 현미경의 조명 경로에 2개의 마이크로렌즈 [5]어레이를 배치함으로써 변동 계수가 1~2%인 고균일한 조명을 실현할 수 있다.
샘플 준비

시료가 형광 현미경 검사에 적합하려면 형광체여야 합니다.형광 샘플을 만드는 방법은 여러 가지가 있습니다. 주요 기술은 형광 얼룩으로 라벨을 붙이거나 생물학적 샘플의 경우 형광 단백질의 발현입니다.또는 시료의 고유 형광(즉, 자가 형광)을 [1]사용할 수 있다.생명과학에서 형광 현미경은 단백질이나 다른 관심 분자의 분포를 감지하기 위해 검체의 특이하고 민감한 염색을 가능하게 하는 강력한 도구이다.그 결과 생물학적 시료의 형광염색을 위한 다양한 기술이 있다.
생체 형광 얼룩
많은 형광 얼룩이 다양한 생물학적 분자를 위해 디자인되었습니다.이들 중 일부는 본질적으로 형광성이고 관심 있는 생물학적 분자와 결합하는 작은 분자들이다.주요 예로는 DNA의 작은 홈과 결합하는 DAPI와 Hoechst(자외선 파장 빛에 의해 들뜸), DRAQ5와 DRAQ7(적색 빛에 의해 최적으로 들뜸)과 같은 핵산 얼룩이 있다.다른 것들은 특정 세포 구조를 결합하고 형광 리포터에 의해 유도된 약물, 독소 또는 펩타이드이다.이러한 형광 염색의 주요 예는 포유류 세포에서 액틴 섬유를 염색하는 데 사용되는 팔로이딘이다.콜라겐 하이브리드화 펩타이드로 알려진 새로운 펩타이드는 또한 불소체와 결합되어 변성된 콜라겐 섬유를 염색하는 데 사용될 수 있습니다.식물세포벽의 염색은 셀룰로오스 또는 펙틴과 결합하는 얼룩 또는 염료를 사용하여 이루어진다.식물 세포의 생생한 영상 촬영이 가능한 높은 특이성을 가진 형광 탐침의 탐색은 현재 [7]진행 중이다.
형광체 또는 형광체라고 불리는 형광체 분자가 많이 있는데, 형광체에는 형광체, Alexa Fluors, 또는 DyLight 488과 같은 형광체 분자가 있으며, 형광체에는 다른 분자와 화학적으로 연결되어 샘플 내에서 관심 대상을 결합할 수 있습니다.
면역 형광
면역 형광은 세포 내의 특정 단백질이나 다른 분자에 라벨을 붙이기 위해 항체에 대한 매우 특이적인 결합을 사용하는 기술이다.시료는 관심 분자에 특이적인 1차 항체로 처리된다.불소포자는 1차 항체에 직접 결합할 수 있다.또는 제1항체에 특이적으로 결합하는 불소체와 결합하는 2차 항체를 사용할 수 있다.예를 들어 플루오로포어에 의해 유도된 2차 항쥐 항체와 조합된 튜불린을 인식하는 마우스 내에서 배양된 1차 항체를 사용하여 세포 내의 미소관을 표시할 수 있다.
형광단백질
유전학에 대한 현대적 이해와 DNA를 수정하는 데 이용 가능한 기술들은 과학자들이 형광 단백질 리포터를 가지고 다니기 위해 단백질을 유전적으로 변형할 수 있게 해준다.생물학적 샘플에서 이것은 과학자가 직접 관심 단백질을 형광체로 만들 수 있게 해준다.그러면 살아있는 세포를 포함하여 단백질 위치를 직접 추적할 수 있습니다.
제한 사항
형광포자는 광표백이라고 불리는 과정에서 빛을 발하면서 형광발광 능력을 잃는다.광표백은 형광분자가 형광 중에 들뜬 전자로부터 화학적 손상을 축적할 때 발생합니다.광 표백은 형광 현미경으로 샘플을 관찰할 수 있는 시간을 크게 제한할 수 있습니다.광표백을 줄이기 위한 몇 가지 기법이 존재한다. 예를 들어, 보다 견고한 형광체의 사용, 조명을 최소화하거나 광보호 스캐빈저 화학물질을 사용한다.
형광 리포터 단백질을 사용한 형광 현미경 검사는 살아있는 세포를 형광 현미경으로 분석할 수 있게 했지만, 세포는 특히 짧은 파장 빛에 광독성에 취약하다.또한 형광분자는 조명 하에서 반응성 화학종을 발생시키는 경향이 있어 광독성 효과를 높일 수 있다.
투과 및 반사광 현미경 기술과 달리 형광 현미경 검사는 형광 라벨이 부착된 특정 구조만 관찰할 수 있습니다.예를 들어 형광현미경법으로 형광DNA 염색으로 제조된 조직시료를 관찰하는 것은 세포 내 DNA의 구성만 밝혀낼 뿐 세포 형태에 대해서는 아무것도 드러내지 않는다.
비형광 영상(예: 밝기 영역)의 형광 신호를 추정하도록 제안하는 계산 기법은 이러한 [8]우려를 줄일 수 있다.일반적으로 이러한 접근법에는 착색 세포에 대한 심층 컨볼루션 신경망을 훈련시킨 다음 비배열 표본에 대한 형광을 추정하는 것이 포함된다.따라서 조사 대상 셀과 네트워크를 훈련하기 위해 사용되는 셀을 분리함으로써 이미징을 보다 빠르게 수행하고 광독성을 줄일 수 있습니다.
서브회절 기술
빛의 파동성은 회절 한계로 인해 빛이 집중될 수 있는 지점의 크기를 제한합니다.이 제한은 19세기에 에른스트 아베에 의해 설명되었고 "광학 현미경의 분해능을 사용된 빛의 파장의 약 절반으로 제한"했다.형광 현미경은 특수한 광학 구성에 의해 이 한계를 넘어서 도달하는 것을 목표로 하는 많은 기술의 중심이다.
20세기에 현미경 검사 기술의 몇 가지 개선이 발명되었고 어느 정도 분해능과 대조를 증가시켰다.그러나 그들은 회절 한계를 극복하지 못했다.1978년 4Pi 현미경을 공초점 레이저 주사 형광 현미경으로 사용하여 이 장벽을 깨기 위한 첫 번째 이론 아이디어가 개발되었습니다. 이 현미경은 빛이 모든 면에서 이상적으로 초점을 맞추고 '포인트 바이 포인트' 여기와 '포인트 바이 포인트'[9] 검출을 결합함으로써 물체를 스캔하는 데 사용됩니다.그러나 1994년 [10]4pi 현미경의 첫 번째 실험 시연은 4pi 현미경을 통해 이루어졌으며, 4pi 현미경은 2개의 대향 대물렌즈 또는 적색 편이광과 다광자 들뜸을 이용한 2광자 들뜸 현미경을 사용하여 가능한 초점 방향의 양을 극대화하였다.
통합상관현미경은 형광현미경과 전자현미경을 결합한 것이다.이를 통해 형광 현미경의 데이터를 라벨링 도구로 [11]사용하면서 전자 현미경으로 초미세 구조와 상황 정보를 시각화할 수 있다.
실제로 서브 회절 분해능을 달성한 최초의 기술은 1994년에 제안된 STED 현미경 검사였다.이 방법과 RESOLFT 개념을 따르는 모든 기술은 빛과 형광 분자 사이의 강한 비선형 상호작용에 의존한다.분자는 각각의 특정 위치에서 구별 가능한 분자 상태 사이에서 강하게 구동되며, 그래서 마침내 빛은 공간의 작은 부분에서만 방출될 수 있고, 따라서 분해능이 높아집니다.
1990년대에도 광시야 현미경을 이용한 초해상도 현미경법이 개발됐다.SPDM 국재 현미경법과 구조화된 레이저 조명(공간 변조 조명, SMI)[12]의 개발로 형광 마커로 얼룩진 세포 나노 구조의 크기 분해능이 크게 향상되었습니다.SPDM의 원리와 SMI의 결합으로 Vericoto SMI 현미경이 [13][14]개발되었습니다.녹색 형광 단백질(GFP)과 같은 정상적인 점멸 형광 염료의 단일 분자 검출은 SPDM의 추가 개발로 달성될 수 있습니다. SPDMphymod 기술은 두 가지 다른 형광 분자 유형을 분자 수준에서 검출 및 계수할 수 있게 합니다(이 기술을 2색 국소라고 합니다).2CLM).[15]
또는 광활성 국재 현미경의 등장은 형광 분자의 비율이 각각 매우 작은 단일 분자의 점멸 또는 전환에 의존함으로써 유사한 결과를 얻을 수 있다.적용된 빛에 대한 분자의 확률적 응답은 또한 매우 비선형적인 상호작용에 해당하며, 이는 서브회절 분해능으로 이어진다.
형광 마이크로그래프 갤러리

액틴 필라멘트를 시각화하기 위해 팔로이딘으로 염색된 골육종 세포의 z-투영.영상은 공초점 현미경으로 촬영되었고, 후속 디콘볼루션은 실험적으로 도출된 점 확산 함수를 사용하여 수행되었다.

현미경 아래 내피세포가 있어요핵은 DAPI로 파란색으로 염색되고, 미세관은 FITC에 결합된 항체로 녹색으로 표시되며, 액틴 필라멘트는 TRITC에 결합된 팔로이딘으로 빨간색으로 표시된다.소폐동맥내피(BPAE)세포
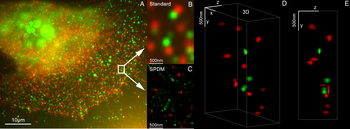
유방 세포에 Her2 및 Her3를 사용한 3D 이중 색상 초해상도 현미경 검사, 표준 염료:알렉사 488, 알렉사 568LIMON 현미경 검사

RFP 및 GFP 형광 마커와 융합된 일부 막 단백질에 의해 시각화된 효모 세포막.양쪽 마커에서 빛이 들어오면 노란색이 됩니다.

초해상도 현미경 검사: 인간 암세포에서 단일 YFP 분자 검출.Verico-SMI/SPDMphymod 현미경으로 측정한 15 nm 범위의 일반적인 거리 측정

초해상도 현미경 : Verico-SMI/SPDMphymod 현미경으로 측정한 광시야 영역(470μm2)에서 GFP 및 RFP 융합단백질(2CLM) 120,000개 국재화 현미경

인간 야생형 및 P239S 돌연변이 팔라딘의 DNA 발현 형광 현미경 검사.

혈액 세포에 빨간 색으로 피해 지역을 보여 주는 것에 태양 폭발의 Fluorescence 현미경 영상 병리학.










「 」를 참조해 주세요.
- 형광 영상
- 생명 과학에서 Fluorescence.
- Correlative Light-Electron 현미경 검사법
- 엘리자베스 해리, 박테리아 세포 이하의 단백질의 시각화에 형광 현미경 기술의 선구자이다.
- 녹색 형광 단백질(GFP)
- 수은등
- 현미경
- 전자 현미경)Cathodoluminescence 검사
- 스토크스 시프트
- 제논 아크 램프
레퍼런스
- ^ a b c d Spring KR, Davidson MW. "Introduction to Fluorescence Microscopy". Nikon MicroscopyU. Retrieved 28 September 2008.
- ^ "The Fluorescence Microscope". Microscopes—Help Scientists Explore Hidden Worlds. The Nobel Foundation. Retrieved 28 September 2008.
- ^ Juan Carlos Stockert, Alfonso Blázquez-Castro (2017). Fluorescence Microscopy in Life Sciences. Bentham Science Publishers. ISBN 978-1-68108-519-7. Retrieved 17 December 2017.
- ^ Huang B (March 2010). "Super resolution fluorescence microscopy". Annual Review of Biochemistry. 78: 993–1016. doi:10.1146/annurev.biochem.77.061906.092014. PMC 2835776. PMID 19489737.
- ^ F.A.W. Coumans; E. van der Pol; L.W.M.M. Terstappen (2012). "Flat-top illumination profile in an epi-fluorescence microscope by dual micro lens arrays". Cytometry Part A. 81 (4): 324–331. doi:10.1002/cyto.a.22029. PMID 22392641. S2CID 13812696.
- ^ Colin, S., Coelho, L.P., Sunagawa, S., Bowler, C., Karsenti, E., Bork, P., Pepperkok, R. 및 De Vargas, C. (2017) "환경 미생물 생물 생물학과 생태학을 위한 정량적 3D 영상촬영"eLife, 6: e26066.doi: 10.7554/eLife.26066.002.
 자료는 Creative Commons Attribution 4.0 International License에 따라 제공되는 이 소스로부터 복사되었습니다.
자료는 Creative Commons Attribution 4.0 International License에 따라 제공되는 이 소스로부터 복사되었습니다. - ^ Bidhendi, AJ; Chebli, Y; Geitmann, A (May 2020). "Fluorescence Visualization of Cellulose and Pectin in the Primary Plant Cell Wall". Journal of Microscopy. 278 (3): 164–181. doi:10.1111/jmi.12895. PMID 32270489. S2CID 215619998.
- ^ Kandel, Mikhail E.; He, Yuchen R.; Lee, Young Jae; Chen, Taylor Hsuan-Yu; Sullivan, Kathryn Michele; Aydin, Onur; Saif, M. Taher A.; Kong, Hyunjoon; Sobh, Nahil; Popescu, Gabriel (2020). "Phase imaging with computational specificity (PICS) for measuring dry mass changes in sub-cellular compartments". Nature Communications. 11: 6256. arXiv:2002.08361. Bibcode:2020NatCo..11.6256K. doi:10.1038/s41467-020-20062-x. PMC 7721808. PMID 33288761. S2CID 212725023.
- ^ Cremer, C; Cremer, T (1978). "Considerations on a laser-scanning-microscope with high resolution and depth of field" (PDF). Microscopica Acta. 81 (1): 31–44. PMID 713859.
- ^ S.W. Hell, E.H.K. Stelzer, S. Lindek, C. Cremer; Stelzer; Lindek; Cremer (1994). "Confocal microscopy with an increased detection aperture: type-B 4Pi confocal microscopy". Optics Letters. 19 (3): 222–224. Bibcode:1994OptL...19..222H. CiteSeerX 10.1.1.501.598. doi:10.1364/OL.19.000222. PMID 19829598.
{{cite journal}}: CS1 maint: 여러 이름: 작성자 목록(링크) - ^ Baarle, Kaitlin van. "Correlative microscopy: Opening up worlds of information with fluorescence". Retrieved 16 February 2017.
- ^ Hausmann, Michael; Schneider, Bernhard; Bradl, Joachim; Cremer, Christoph G. (1997), "High-precision distance microscopy of 3D nanostructures by a spatially modulated excitation fluorescence microscope" (PDF), in Bigio, Irving J; Schneckenburger, Herbert; Slavik, Jan; et al. (eds.), Optical Biopsies and Microscopic Techniques II, Optical Biopsies and Microscopic Techniques II, vol. 3197, p. 217, doi:10.1117/12.297969, S2CID 49339042
- ^ Reymann, J; Baddeley, D; Gunkel, M; Lemmer, P; Stadter, W; Jegou, T; Rippe, K; Cremer, C; Birk, U (2008). "High-precision structural analysis of subnuclear complexes in fixed and live cells via spatially modulated illumination (SMI) microscopy" (PDF). Chromosome Research. 16 (3): 367–82. doi:10.1007/s10577-008-1238-2. PMID 18461478. S2CID 22811346.
- ^ Baddeley, D; Batram, C; Weiland, Y; Cremer, C; Birk, UJ (2003). "Nanostructure analysis using spatially modulated illumination microscopy" (PDF). Nature Protocols. 2 (10): 2640–6. doi:10.1038/nprot.2007.399. PMID 17948007. S2CID 22042676.[데드링크]
- ^ Gunkel, M; Erdel, F; Rippe, K; Lemmer, P; Kaufmann, R; Hörmann, C; Amberger, R; Cremer, C (2009). "Dual color localization microscopy of cellular nanostructures" (PDF). Biotechnology Journal. 4 (6): 927–38. doi:10.1002/biot.200900005. PMID 19548231. S2CID 18162278.
외부 링크
| 라이브러리 리소스 정보 형광 현미경법 |
- Fluorophores.org, 형광[permanent dead link] 염료 데이터베이스
- 현미경 자원 센터
- 형광 현미경 및 공초점 현미경을 포함한 다양한 종류의 현미경에 대한 애니메이션과 설명(파리 수드 대학)
| 조명 및 콘트라스트법 |  | |
|---|---|---|
| 형광법 | ||
| 서브회절 제한 기술 | ||


