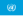단풍색
Autumn leaf color
단풍색은 가을철에 몇 주 동안 노란색, 주황색, 빨간색, 보라색, [1]갈색의 다양한 색조를 띠는 많은 낙엽성 나무와 관목의 정상적인 녹색 잎에 영향을 미치는 현상이다.이 현상은 일반적으로 영국식 영어로는 단풍[2] 또는[3] 단풍이라고 불리며, 미국식 영어로는 단풍,[4] 단풍 또는 단순[5] 단풍이라고 불린다.
캐나다와 미국의 일부 지역에서는 "잎 피핑" 관광이 경제 활동에 큰 기여를 하고 있다.이 관광 활동은 보통 북반구에서는 9월과 10월, 남반구에서는 4월과 5월 사이에 색상의 변화가 시작되고 잎이 떨어지기 시작할 때 일어난다.
클로로필 및 녹색/노란색/오렌지색



녹색 잎은 엽록소라고 불리는 세포소기관 안에 있는 엽록소라고 알려진 색소의 존재 때문에 녹색이다.성장기처럼 잎의 세포에 풍부하게 있을 때, 엽록소의 녹색은 잎에 존재할 수 있는 다른 색소의 색을 지배하고 가립니다.그래서 여름의 잎들은 특징적으로 [6]푸르다.
클로로필은 중요한 기능을 가지고 있다: 그것은 태양 광선을 포착하고 그 결과 생기는 에너지를 식물 먹이의 제조에 사용한다 – 물과 이산화탄소에서 생산되는 단당이다.이 설탕들은 식물 영양의 기초가 됩니다 – 성장과 발달을 위해 필요한 탄수화물의 유일한 공급원입니다.그들의 식품 제조 과정에서 엽록소는 분해되고, 따라서 지속적으로 "소모되고" 있다.그러나, 성장기에는, 식물은 엽록소를 보충하여 공급량이 높고 잎이 녹색으로 유지되도록 한다.
늦여름에, 낮 시간이 짧아지고 온도가 내려가면서, 각 잎의 밑부분에 특별한 코르크 세포 층이 형성되면서 잎 안으로 그리고 잎 밖으로 액체를 운반하는 정맥이 점차적으로 닫힙니다.이 코르크 층이 발달함에 따라, 처음에는 천천히, 나중에는 더 빠르게 잎에 수분과 미네랄의 섭취가 감소합니다.이 기간 동안, 잎에 있는 엽록소의 양이 감소하기 시작합니다.종종, 정맥들 사이의 조직이 거의 완전히 변한 후에도 여전히 녹색을 띤다만, 정맥들은 여전히 녹색이다.
클로로필은 엽록체의 틸라코이드 막에 위치하고 아포단백질과 여러 개의 리간드로 구성되어 있으며, 그 중 가장 중요한 것은 클로로필 a와 b이다.가을이 되면 이 단지는 무너진다.클로로필 분해는 먼저 일어나는 것으로 생각된다.연구에 따르면 클로로필 분해의 시작은 클로로필 b 환원효소에 의해 촉매되며, 클로로필 b는 7-히드록시메틸 클로로필 a로 환원되고, 클로로필 [7]a는 클로로필 a로 환원된다.이것은 아포단백질의 분해가 일어나는 복합체를 불안정하게 만드는 것으로 여겨진다.아포단백질의 분해에서 중요한 효소는 FtsH6이며, FtsH 계열의 단백질 분해 [8]효소에 속한다.
클로로필은 비형광성 클로로필 [9]이화물로 알려진 무색의 테트라피롤로 분해된다.클로로필이 분해됨에 따라 황색 크산토필과 오렌지색 베타카로틴의 숨겨진 색소가 드러난다.
다른 색상의 원인이 되는 안료

카로티노이드
카로티노이드는 일년 내내 잎에 존재하지만, 오렌지-노란색은 보통 녹색 엽록소에 [6]의해 가려집니다.가을이 다가옴에 따라 식물 내부와 외부의 어떤 영향이 클로로필을 소모되는 속도보다 더 느린 속도로 대체한다.이 기간 동안, 엽록소의 총 공급이 점차 감소하면서, "마스크링" 효과는 서서히 사라집니다.그러면 엽록소와 함께 존재하는 다른 색소들이 잎의 세포를 [6]통해 보이기 시작합니다.이것들은 카로티노이드이고 노란색, 갈색, 주황색 그리고 그 사이의 많은 색조를 제공합니다.
카로티노이드는 엽록소 색소와 함께 잎 세포 내의 플라스티드라고 불리는 작은 구조에서 발생합니다.때때로, 그것들은 잎에 매우 풍부해서 심지어 여름에도 식물에 황록색을 입힌다.하지만 보통 잎들이 엽록소를 잃기 시작하는 가을에 처음으로 눈에 띄게 된다.
카로티노이드는 당근, 옥수수, 카나리아, 그리고 수선화뿐만 아니라 계란 노른자, 루타바가, 버터컵, 그리고 바나나에 특징적인 색을 주면서 많은 생물에서 흔하다.
그들의 빛나는 노란색과 오렌지는 히코리, 재, 단풍나무, 노란 포플러, 아스펜, 자작나무, 검은 체리, 무화나무, 목화나무, 사사프라, 그리고 앨더와 같은 단단한 나무의 잎을 물들인다.카로티노이드는 나무 [6]종의 약 15-30%를 색소화하는 주요 색소이다.
안토시아닌류
가을 단풍을 장식하는 붉은색, 보라색, 그리고 그들의 혼합된 조합은 안토시아닌이라고 불리는 세포에 있는 다른 색소 그룹으로부터 옵니다.카로티노이드와는 달리, 이 색소들은 성장기 내내 잎에 존재하지 않지만,[6] 여름이 끝날 무렵에 활발하게 생산된다.그것들은 늦여름에 잎 세포의 수액 속에서 발달하며, 이 발달은 식물 내부와 외부 모두에서 많은 영향의 복잡한 상호작용의 결과이다.잎의 인산염 수치가 [10]감소함에 따라 밝은 빛이 존재하는 당이 분해되는 것에 따라 형성됩니다.

여름 생육기에는 인산염 수치가 높다.그것은 엽록소에 의해 만들어진 당을 분해하는 데 중요한 역할을 하지만, 가을에는 다른 화학 물질과 영양소와 함께, 인산염이 잎에서 식물의 줄기 안으로 이동합니다.이 경우 설탕 분해 과정이 바뀌어 안토시아닌 색소가 생성된다.이 기간 동안 빛이 밝을수록 안토시아닌의 생산량이 증가하고 색상의 디스플레이가 더욱 밝아집니다.가을의 낮이 밝고 시원하고, 밤은 쌀쌀하지만 얼지 않을 때, 가장 밝은 색이 발달합니다.
안토시아닌은 초봄에 싹에서 펼쳐질 때 아주 어린 잎의 가장자리를 일시적으로 물들인다.그들은 또한 크랜베리, 빨간 사과, 블루베리, 체리, 딸기, 자두와 같은 흔한 과일에도 친숙한 색을 입힌다.
안토시아닌은 온대 지역의 나무 종의 약 10%에 존재하지만, 특정 지역(가장 유명한 뉴잉글랜드 북부)에서는 70%까지 안토시아닌을 [6]생성할 수 있습니다.가을 숲에서는 단풍나무, 떡갈나무, 신나무, 단풍나무, 개나무, 투펠로, 벚나무, 감나무에서 생생하게 볼 수 있다.이 같은 색소들은 종종 카로티노이드의 색과 결합하여 많은 단단한 나무 종들의 전형적인 더 깊은 주황색, 불타는 붉은색, 그리고 브론즈를 만듭니다.
세포벽
잎의 갈색은 색소의 결과라기보다는 세포벽의 결과인데,[6] 색소가 보이지 않을 때 명백할 수 있다.
단풍의 기능
낙엽성 식물은 전통적으로 가을에 잎을 잃는다고 믿었는데, 이는 낙엽성 식물의 유지비가 낮은 빛과 추운 [11]기온의 겨울 기간 동안 광합성의 혜택을 능가하기 때문이다.많은 경우, 이것은 지나치게 단순한 것으로 판명되었습니다 – 다른 요인에는 곤충의 포식,[12] 수분 손실, 강풍이나 강설로 인한 피해 등이 포함됩니다.
안토시아닌은 가을에 활발하게 생산되지만 낙엽에는 관여하지 않는다.낙엽에서 색소 생성의 역할에 대한 많은 가설이 제안되었고, 일반적으로 두 가지 범주로 분류된다:[6] 동물과의 상호작용과 비생물학적 요인으로부터의 보호.
포토 프로텍션
광보호 이론에 따르면, 안토시아닌은 낮은 [13][14]온도에서 빛의 해로운 영향으로부터 잎을 보호합니다.낙엽이 지려고 하기 때문에 나무 보호는 그다지 중요하지 않습니다.그러나 광산화 및 광억제는 특히 저온에서 영양소를 재흡수하는 과정을 덜 효율적으로 만든다.광보호 이론에 따르면 잎을 안토시아닌으로 보호함으로써 나무는 영양소(특히 질소)를 더 효율적으로 재흡수할 수 있다.
공진화

공진화 [15]이론에 따르면, 이 색깔들은 겨울을 나기 위해 나무를 숙주로 사용하는 진딧물과 같은 곤충들에게 경고 신호이다.만약 그 색깔이 곤충에 대한 화학적 방어의 양과 관련이 있다면, 곤충들은 붉은 잎을 피하고 그들의 건강을 증진시킬 것이다; 동시에, 붉은 잎을 가진 나무들은 기생충의 부하를 줄이기 때문에 유리하다.이는 야생 사과와 달리 일부 재배 사과 품종이 가을에 붉은 잎이 없는 사과나무의 사례에서 나타났다.붉은 잎을 가진 사과나무를 피하는 진딧물의 비율은 그렇지 [16]않은 진딧물보다 더 많이 자라고 발달한다.게다가 붉은 잎을 가진 품종이 더 작은 열매를 맺기 때문에 과일 크기, 잎 색깔, 진딧물 저항성 사이에 트레이드오프가 존재하며, 이는 진딧물 [16]침입 감소의 더 큰 필요성과 관련된 붉은 잎의 생산 비용을 시사한다.
진딧물의 생존을 감소시키는 붉은 잎의 나무와 마찬가지로, 밝은 잎을 가진 나무 종은 밝은 잎이 없는 나무보다 더 전문적인 진딧물 해충을 선택하는 경향이 있다(가을철에 [17]병충해와 함께 진화하는 나무에만 가을 색이 유용하다).한 연구는 단풍나무의 곤충 초본(잎을 먹는 손상)을 시뮬레이션한 결과 [18]손상되지 않은 나무보다 더 일찍 붉은색을 띠는 것을 발견했다.
가을 색깔의 공진화 이론은 2001년 W.[17] D. 해밀턴에 의해 진화 신호론의 예로 제안되었습니다.붉은 잎과 같은 생물학적 신호는 생산 비용이 많이 들기 때문에 대개 정직하기 때문에 저품질의 개인들이 가짜를 하지 못하고 속일 수 없는 것으로 신호인의 진정한 품질을 알리는 것이라고 주장합니다.단풍색은 생산하는데 비용이 많이 들거나 가짜가 불가능하다면 신호가 될 것이다.[citation needed]
가을 전에 잎 색깔을 바꾸는 것은 초식동물의 [19]위장을 약화시키는 데 도움이 될 수 있는 적응으로 제안되었다.
베리를 가진 많은 식물들은 특히 눈에 보이는 베리와/또는 잎 색깔, 특히 밝은 붉은색을 가진 새들을 유혹합니다.새들은 식사를 하는 반면, 관목, 덩굴, 또는 전형적으로 작은 나무들은 소화되지 않은 씨앗을 운반하여 새들의 거름과 함께 퇴적시킨다.옻나무는 밝은 붉은 잎을 가진 것으로 특히 유명하다.
알레오파시
단풍나무의 일부 종의 선명한 붉은 가을 색은 엽록소 분해 과정과 분리된 과정으로 만들어집니다.이 나무가 변화하고 힘든 계절의 에너지 수요에 대처하는 데 어려움을 겪을 때, 단풍나무는 안토시아닌을 생성하기 위한 추가적인 대사 지출에 관여합니다.시각적인 붉은 색조를 만드는 이러한 안토시아닌은 근처의 [20]묘목의 성장을 방해함으로써 종간 경쟁을 돕는 것으로 밝혀졌다.
관광업
낙엽수가 발견되는 곳이면 가을의 색채가 나타나지만, 가장 밝은 색상의 단풍은 캐나다 남부 대부분, 미국 북부 일부 지역, 알프스 북쪽의 북유럽과 서유럽, 흑해 근처의 러시아의 코카서스 지역, 그리고 동쪽을 포함한 북반구에서 발견된다.ia(중국 북부와 동부, 그리고 한국과 [21][22]일본을 포함)
남반구에서는 아르헨티나 남부와 중부, 브라질 남부와 남동부, 호주 남동부(태즈메이니아 [23]포함)에서 다채로운 단풍을 볼 수 있다.
기후의 영향
서유럽(남유럽 제외)에 비해, 북미에서는 훨씬 더 많은 수목종([24]서유럽에서는 각각 51종과 3종에 비해 800종 이상, 약 70개의 떡갈나무)이 제공되고 있어 더 다양한 색채를 더하고 있다.주된 이유는 빙하기의 영향이 다르기 때문이다 – 북미에서는 종들이 남북 산맥을 따라 더 많은 남쪽 지역에서 보호되었지만,[25] 유럽에서는 그렇지 않았다.
지구 온난화와 대기 중의 이산화탄소 수치 상승은 향후 북부 경목림에서 색이 변하고 낙엽이 지는 일반적인 가을 광경을 지연시키고 산림 생산성을 [26]높일 수 있다.특히, 미국 북동부의 더 높은 가을 기온이 색의 [27]변화를 지연시키고 있다.포플러 나무로 실험한 결과, 포플러 나무는 온도 [26]변화와는 무관하게 CO 수치가2 높을수록 더 오래 녹색을 유지하는 것으로 나타났습니다.하지만, 2년에 걸친 실험은 시간이 지남에 따라 성숙한 숲이 얼마나 영향을 받을 수 있는지를 나타내기에는 너무 짧았다.하바륨 표본 150년을 사용한 다른 연구들은 19세기 이후 가을이 시작될 때 한 달 이상 지연되는 것을 발견했고 곤충, 바이러스, 가뭄 스트레스가 단풍나무의 [27][28]가을 색채 시기에 영향을 줄 수 있다는 것을 발견했다.또한, 지면에 가까운 오존 수치의 증가(지구권 오존 오염)와 같은 다른 요소들은 이산화탄소의 [29]증가의 유익한 효과를 부정할 수 있다.





.jpg)




레퍼런스
![]() 이 문서에는 미국 정부 문서인 "USDA 산림청"의 퍼블릭 도메인 자료가 포함되어 있습니다.
이 문서에는 미국 정부 문서인 "USDA 산림청"의 퍼블릭 도메인 자료가 포함되어 있습니다.
- ^ "The Science of Color in Autumn Leaves". usna.usda.gov. United States National Arboretum. October 6, 2011. Archived from the original on January 11, 2018. Retrieved June 18, 2015.
- ^ Wade, Paul; Arnold, Kathy (September 16, 2014). "New England in the Fall: Trip of a Lifetime - Telegraph". telegraph.co.uk. The Daily Telegraph. Retrieved June 18, 2015.
- ^ "BBC - Gardening - How to be a gardener - The gardening year - Autumn's theme". bbc.co.uk. BBC. September 17, 2014. Retrieved June 18, 2015.
- ^ "US Forest Service - Caring for the land and serving people". fs.fed.us. United States Forest Service. 2014. Retrieved June 18, 2015.
- ^ "MaineFoliage.com - Maine's Official Fall Foliage Website". MaineFoliage.com. MaineFoliage.com. 2013. Retrieved June 18, 2015.
- ^ a b c d e f g h Archetti, Marco; Döring, Thomas F.; Hagen, Snorre B.; Hughes, Nicole M.; Leather, Simon R.; Lee, David W.; Lev-Yadun, Simcha; Manetas, Yiannis; Ougham, Helen J. (2011). "Unravelling the evolution of autumn colours: an interdisciplinary approach". Trends in Ecology & Evolution. 24 (3): 166–73. doi:10.1016/j.tree.2008.10.006. PMID 19178979.
- ^ Horie, Y.; Ito, H.; Kusaba, M.; Tanaka, R.; Tanaka, A. (2009). "Participation of Chlorophyll b Reductase in the Initial Step of the Degradation of Light-harvesting Chlorophyll a/b-Protein Complexes in Arabidopsis". Journal of Biological Chemistry. 284 (26): 17449–56. doi:10.1074/jbc.M109.008912. PMC 2719385. PMID 19403948.
- ^ Zelisko, A.; Garcia-Lorenzo, M.; Jackowski, G.; Jansson, S.; Funk, C. (2005). "AtFtsH6 is involved in the degradation of the light-harvesting complex II during high-light acclimation and senescence". Proceedings of the National Academy of Sciences. 102 (38): 13699–704. Bibcode:2005PNAS..10213699Z. doi:10.1073/pnas.0503472102. PMC 1224624. PMID 16157880.
- ^ Hortensteiner, S. (2006). "Chlorophyll degradation during senescence". Annual Review of Plant Biology. 57: 55–77. doi:10.1146/annurev.arplant.57.032905.105212. PMID 16669755.
- ^ Davies, Kevin M. (2004). Plant pigments and their manipulation. Wiley-Blackwell. p. 6. ISBN 978-1-4051-1737-1.
- ^ Thomas, H; Stoddart, J L (1980). "Leaf Senescence". Annual Review of Plant Physiology. 31: 83–111. doi:10.1146/annurev.pp.31.060180.000503.
- ^ Labandeira, C. C.; Dilcher, DL; Davis, DR; Wagner, DL (1994). "Ninety-seven million years of angiosperm-insect association: paleobiological insights into the meaning of coevolution". Proceedings of the National Academy of Sciences. 91 (25): 12278–82. Bibcode:1994PNAS...9112278L. doi:10.1073/pnas.91.25.12278. PMC 45420. PMID 11607501.
- ^ Lee, David; Gould, Kevin (2002). "Why Leaves Turn Red". American Scientist. 90 (6): 524–531. Bibcode:2002AmSci..90..524L. doi:10.1511/2002.6.524.
- ^ Lee, D; Gould, K (2002). "Anthocyanins in leaves and other vegetative organs: An introduction". Advances in Botanical Research. 37: 1–16. doi:10.1016/S0065-2296(02)37040-X. ISBN 978-0-12-005937-9.
- ^ Archetti, M; Brown, S. P. (June 2004). "The coevolution theory of autumn colours". Proc. Biol. Sci. 271 (1545): 1219–23. doi:10.1098/rspb.2004.2728. PMC 1691721. PMID 15306345.
- ^ a b Archetti, M. (2009). "Evidence from the domestication of apple for the maintenance of autumn colours by coevolution". Proceedings of the Royal Society B: Biological Sciences. 276 (1667): 2575–80. doi:10.1098/rspb.2009.0355. PMC 2684696. PMID 19369261.
- ^ a b Hamilton, W. D.; Brown, S. P. (2001). "Autumn tree colours as a handicap signal". Proceedings of the Royal Society B: Biological Sciences. 268 (1475): 1489–93. doi:10.1098/rspb.2001.1672. PMC 1088768. PMID 11454293.
- ^ Forkner, Rebecca E. (May 1, 2014). "Simulated herbivory advances autumn phenology in Acer rubrum". International Journal of Biometeorology. 58 (4): 499–507. Bibcode:2014IJBm...58..499F. doi:10.1007/s00484-013-0701-8. PMID 23832182. S2CID 24879283.
- ^ Lev-Yadun, Simcha; Dafni, Amots; Flaishman, Moshe A.; Inbar, Moshe; Izhaki, Ido; Katzir, Gadi; Ne'eman, Gidi (2004). "Plant coloration undermines herbivorous insect camouflage". BioEssays. 26 (10): 1126–30. doi:10.1002/bies.20112. PMID 15382135.
- ^ (Frey & Eldridge,[citation needed] 2005년)
- ^ "Pest Alert". South Dakota State University. August 30, 1998. Archived from the original on October 20, 2006. Retrieved November 28, 2006.
- ^ Altman, Daniel (November 8, 2006). "Fall foliage sets Japan ablaze". Taipei Times. Retrieved November 28, 2006.
- ^ Hutchinson, Carrie (March 2, 2019). "The 5 Best Places in Australia to See Autumn Colours". Qantas. Retrieved July 22, 2019.
- ^ 하인즈 엘렌버그, H. 엘렌버그:식물 Mitteleuropas mit den Alpen:ökologischer, dynamischer und histor Sicht, UTB, 슈투트가르트, 제5판, 독일어, ISBN 3-8252-8104-3, 1996년[page needed]
- ^ "Botanik online: Pflanzengesellschaften - Sommergrüne Laub- und Mischlaubwälder" (in German). University of Hamburg Biology Server. Archived from the original on October 6, 2014. Retrieved July 29, 2020.
- ^ a b Taylor, Gail; Tallis, Matthew J.; Giardina, Christian P.; Percy, Kevin E.; Miglietta, Franco; Gupta, Pooja S.; Gioli, Beniamino; Calfapietra, Carlo; Gielen, Birgit (2007). "Future atmospheric CO2 leads to delayed autumnal senescence". Global Change Biology. 14 (2): 264–75. Bibcode:2008GCBio..14..264T. CiteSeerX 10.1.1.384.1142. doi:10.1111/j.1365-2486.2007.01473.x.
- ^ a b Gibbens, Sarah (November 24, 2021). "Fall foliage was disrupted by climate change. It might be the new normal". Environment. National Geographic.
- ^ Garretson, Alexis; Forkner, Rebecca E. (2021). "Herbaria Reveal Herbivory and Pathogen Increases and Shifts in Senescence for Northeastern United States Maples Over 150 Years". Frontiers in Forests and Global Change. 4: 185. doi:10.3389/ffgc.2021.664763.
- ^ "Forests Could Benefit When Fall Color Comes Late". Newswise. Retrieved August 17, 2008.
추가 정보
- Guy, Robert D.; Krakowski, Jodie (2003). "Autumn Colours – Nature's Canvas is a Silk Parasol" (PDF). Davidsonia. 14 (4): 111–20. Archived from the original (PDF) on October 19, 2013. Retrieved May 18, 2007.
외부 링크
- 헨리 데이비드 소로의 가을 색조
- 가을에 흔히 볼 수 있는 나무를 색깔로 구별하다
- Sanderson, Katharine (2007). "Why autumn leaves turn red". Nature. doi:10.1038/news.2007.202.
| 국립도서관 | |
|---|---|
| 다른. | |