폴리아민
PolyamineA폴리아민 유기 화합물 두개 이상 아미노산 그룹.알킬기 폴리아민들 자연스럽게 있지만 몇가지 합성을 한다.Alkylpolyamines, 흡습성과 수용성 무색이다.중성 pH가까이에, 그들은 암모늄 파생 상품으로 존재한다.[1]방 온도에서 가장 향기로운 폴리아민들은 결정질 고체이다.
천연 폴리아민들
Low-molecular-weight 선형 폴리아민들 삶의 모든 형태에서 발견된다.주된 예는 triamine과tetraamine 스페르미와 스페르민.그들은 구조상으로나 생합성으로 그 diamines 보통과 카다베린과 관련 있다.Polyamine 신진대사 효소를 오르니틴 decarboxylase(ODC)의 활동에 의해 규제된다.[2]폴리아민은 포유류의 [3]뇌에서 고농도로 발견된다.
- 천연 폴리아민
합성 폴리아민
몇몇 합성 폴리아민은 화학 산업과 연구실에서 사용된다.주로 모터 오일의 첨가제 및 에폭시 [4]수지와 공동 반응제(경화제)로 사용됩니다.많은 합성 폴리아민은 NCHCHN22 링크를 갖추고 있습니다.
- 디에틸렌트리아민(약칭 디엔 또는 DETA), (HN-CHCH-NH-CH-NH222222).관련된 페르메틸화 유도체 펜타메틸디에틸렌트리아민은 유기리튬 화학에서 킬레이트제로 사용된다.
- 트리에틸렌테트라민(트리엔 또는 테타, HN-CH-NH-CH-NH-NH22222222), 테트라에틸렌페타민(TEPA2222222222, HN-CH-NH-CH-NH-NH-CH-NH-CH-NH),
- 대환상 폴리아민: 1,4,7-트리아자사이클로난(NHCH22)3 및 사이클렌(NHCH22)4관련된 테트라아자 매크로 사이클은 사이클람이다.
- 트리스(2-아미노에틸)아민(N(CHCHNH222))3은 분지 폴리아민이다.관련된 삼각 폴리아민은 1,1,1-트리(아미노메틸)에탄이다.
- 폴리에틸렌아민은 아지리딘에서 유래한 중합체이다.
- 합성 폴리아민
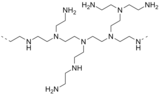
폴리에틸렌아민 서브유닛
amine.svg)


ethane.svg)

니트로겐 사이에 메틸렌 브리지만을 가진 폴리아민의 드문 예로는 1,3,5-트리아지난이 있다.
생물학적 기능
폴리아민의 생합성은 고도로 조절되는 것으로 알려져 있지만, 폴리아민의 생물학적 기능은 부분적으로만 이해된다.양이온성 암모늄 형태에서는 DNA에 결합하고 구조적으로는 일정한 간격으로 발견되는 양이온을 가진 화합물을 나타냅니다(점전하인 Mg 또는2+
Ca와는 대조적으로2+
).그들은 또한 번역 [5]중에 프로그램된 리보솜 프레임시프트의 촉진자 역할을 하는 것으로 밝혀졌다.
폴리아민 생합성 억제는 세포 성장을 늦추거나 멈추게 한다.외인성 폴리아민의 공급은 이 세포들의 성장을 회복시킨다.대부분의 진핵 세포는 세포막에서 외인성 폴리아민의 내부화를 촉진하는 폴리아민 운반 ATP 효소를 발현한다.이 시스템은 빠르게 증식하는 세포에서 매우 활성화되어 있으며 현재 [6]개발 중인 일부 화학 요법의 대상이다.
폴리아민은 또한 NMDA 수용체와 AMPA 수용체를 포함한 다양한 이온 채널의 조절제이다.이들은 내향-정류 칼륨 채널을 차단하여 채널의 전류가 내향으로 정류되도록 함으로써 세포막 전체의 세포 에너지, 즉+
K 이온 구배를 보존한다.또한 폴리아민은 콜리신 E7 오퍼론의 SOS 응답 발현을 개시하고 콜리신 E7 흡수에 필수적인 단백질을 다운 조절함으로써 스트레스 [7]조건 하에서 콜리신 생성 대장균에 생존 우위를 부여한다.
폴리아민은 혈액-뇌 [8]장벽의 투과성을 높일 수 있다.
이들은 식물의 장기 노화를 조절하는 데 관여하며, 따라서 식물 [9]호르몬으로 간주됩니다.또한 이들은 프로그램된 세포사 [10]조절에 직접 관여한다.
Homology-directed DNA수선
폴리아민은 상동성 재조합(HR) 매개 이중 가닥 절단([11]DSB) 복구를 촉진합니다.폴리아민은 RAD51 재조합효소의 DNA 가닥 교환 활성을 강화한다.폴리아민의 고갈은 세포를 이온화 방사선과 자외선 방사선과 같은 유전독성 물질에 민감하게 만든다.RAD51에 대한 폴리아민의 영향은 상동 이중 DNA의 포획을 강화하고 RAD-51 매개 상동 DNA 쌍 및 교환 [11]활동을 촉진하는 능력에서 발생한다.폴리아민은 재조합효소 활성을 조절하는데 진화적으로 보존된 역할을 하는 것으로 보인다.
Biosynthesis 스퍼미 딘, 특히, thermospermine.

탈카르복실화 S-아데노실-L-메티오닌(SAM)의 아미노프로필기를 이용하여 푸트레신으로부터 스펠미딘을 합성한다.그 반응은 스펠미딘 합성효소에 [12]의해 촉매된다.
스펠민은 스펠민 합성효소의 존재 하에서 SAM과 스펠미딘의 반응으로부터 합성된다.
폴리아민은 폴리아민 사이클에서 빠른 상호변환을 거치며, 이 경우 푸트레신은 스펠미딘과 스펠민의 합성을 유도하고, 이러한 폴리아민의 분해로 푸트레신을 형성하며, 이는 [12]사이클을 다시 시작할 수 있다.
테르모스페르민(NH-2(CH2)-3NH-(CH2)-34NH-2(CH)-NH2)-NH)는 스펠민의 구조 이성질체이며 새로운 유형의 식물 성장 조절제이다.그것은 ACAULIS5(ACL5)[13]라는 유전자에 의해 암호화되는 서모스페르민 합성효소의 작용에 의해 스펠미딘으로부터 생산된다.
Polyamine analogues
세포 성장에 있어 폴리아민의 중요한 역할은 폴리아민 대사를 방해하는 많은 약물의 개발을 이끌었다.이 약들은 암 치료에 사용된다.폴리아민은 증식 [14]및 아포토시스의 제한으로 이어지는 세포에서 p53을 상향 조절한다.그것은 또한 ER [15]양성 유방암에서 에스트로겐 수용체 알파의 발현을 감소시킨다.
레퍼런스
- ^ Eller, Karsten; Henkes, Erhard; Rossbacher, Roland; Höke, Hartmut (2005). "Amines, Aliphatic". Ullmann's Encyclopedia of Industrial Chemistry. Weinheim: Wiley-VCH. doi:10.1002/14356007.a02_001.
- ^ Pegg, AE; McCann, PP (1982). "Polyamine metabolism and function". American Journal of Physiology. 243 (5): 212–21. doi:10.1152/ajpcell.1982.243.5.C212. PMID 6814260.
- ^ Seiler, N (1992). "Polyamines". Handbook of Neurochemistry. Vol. 1. New York, NY: Plenum Publishing Corp. pp. 223–55.
- ^ Lawrence, Stephen A. (2004). Amines: synthesis, properties and applications. Cambridge University Press. p. 64. ISBN 978-0-521-78284-5.
- ^ Rato C; Amirova S.R; Bates D.G; Stansfield I; Wallace H.M (June 2011). "Translational recoding as a feedback controller: systems approaches reveal polyamine-specific effects on the antizyme ribosomal frameshift". Nucleic Acids Res. 39 (11): 4587–4597. doi:10.1093/nar/gkq1349. PMC 3113565. PMID 21303766.
- ^ Wang C, Delcros JG, Cannon L, et al. (November 2003). "Defining the molecular requirements for the selective delivery of polyamine conjugates into cells containing active polyamine transporters". J. Med. Chem. 46 (24): 5129–38. doi:10.1021/jm030223a. PMID 14613316.
- ^ Yi-Hsuan Pan; Chen-Chung Liao (May 2006). "The critical roles of polyamines regulating ColE7 production and restricting ColE7 uptake of the colicin-producing Escherichia coli". J. Biol. Chem. 281 (19): 13083–13091. doi:10.1074/jbc.M511365200. PMID 16549429.
- ^ Zhang L, Lee HK, Pruess TH, White HS, Bulaj G (March 2009). "Synthesis and applications of polyamine amino acid residues: improving the bioactivity of an analgesic neuropeptide, neurotensin". J. Med. Chem. 52 (6): 1514–7. doi:10.1021/jm801481y. PMC 2694617. PMID 19236044.
- ^ Pandey S, Ranade SA, Nagar PK, Kumar N (September 2000). "Role of polyamines and ethylene as modulators of plant senescence". J. Biosci. 25 (3): 291–9. doi:10.1007/BF02703938. PMID 11022232. S2CID 21925829.
- ^ Moschou, PN; Roubelakis-Angelakis, KA (Nov 11, 2013). "Polyamines and programmed cell death". Journal of Experimental Botany. 65 (5): 1285–1296. doi:10.1093/jxb/ert373. PMID 24218329.
- ^ a b Lee CY, Su GC, Huang WY, Ko MY, Yeh HY, Chang GD, Lin SJ, Chi P. 폴리아민에 의한 호몰로지 유도 DNA 수복 촉진.NAT Communic.2019년 1월 8일;10 (1):65.doi:10.1038/s41467-018-08011-1.PMID:30622262,PMCID:PMC6325121
- ^ a b Pál M, Szalai G, Janda T (2015). "Speculation: Polyamines are important in abiotic stress signaling" (PDF). Plant Science. 237: 16–23. doi:10.1016/j.plantsci.2015.05.003. PMID 26089148.
- ^ Takano, A; Kakehi, J; Takahashi, T (April 2012). "Thermospermine is not a minor polyamine in the plant kingdom". Plant Cell Physiol. 53 (4): 606–16. doi:10.1093/pcp/pcs019. PMID 22366038.
- ^ Huang, Yi; Pledgie, Allison; Rubin, Ethel; Marton, Laurence J.; Woster, Patrick M.; Sukumar, Saraswati; Casero, Robert A.; Davidson, Nancy E. (September 2005). "Role of p53/p21(Waf1/Cip1) in the regulation of polyamine analogue-induced growth inhibition and cell death in human breast cancer cells". Cancer Biology & Therapy. 4 (9): 1006–1013. doi:10.4161/cbt.4.9.1970. PMC 3639297. PMID 16131835.
- ^ Huang, Y; Keen, JC; Pledgie, A; Marton, LJ; Zhu, T; Sukumar, S; Park, BH; Blair, B; Brenner, K; Casero, RA Jr; Davidson, NE (2006). "Polyamine analogues down-regulate estrogen receptor alpha expression in human breast cancer cells". J Biol Chem. 281 (28): 19055–63. doi:10.1074/jbc.M600910200. PMC 3623667. PMID 16679312.


