코콩차
Nasal concha| 코콩카/터부네이트 | |
|---|---|
 | |
 | |
| 세부 사항 | |
| 식별자 | |
| 라틴어 | 콩채나물 |
| 메쉬 | D014420 |
| FMA | 57456 |
| 뼈의 해부학적 용어 | |

해부학에서, 여러 개의 콩채(/kknnk//) 또는 코 탁도(/[1][2]kknnki),/)는 인간과 다양한 동물에서 코의 호흡 통로에 돌출된 길고 좁고 구부러진 뼈 선반이다.콘채는 긴 조개껍데기처럼 생겼는데, 이 조개껍데기는 그들에게 이름을 지어주었다.콩카는 척추동물에서 [3]비강의 스펀지 모양의 뼈 중 하나이다.
인간의 경우, 콘채는 코 기도를 4개의 홈 모양의 공기 통로로 나누고, 흡입된 공기가 코 점막의 가능한 가장 큰 표면적을 중심으로 일정하고 규칙적인 패턴으로 흐르도록 하는 역할을 합니다.코 점막은 혈액 공급이 얕은 섬모성 점막으로 흡입된 공기를 정화 및 따뜻하게 하여 폐에 대비합니다.
이러한 뼈에 대한 동맥 순환이 급속히 확장되면 체심의 급격한 냉각에 반응하여 내부 압력이 급격히 증가할 수 있습니다.이러한 압력으로 인한 고통은 종종 "뇌 동결"이라고 불리며, 종종 아이스크림의 빠른 소비와 관련이 있습니다.점막의 정맥혈 공급의 얕음은 코피가 쉽게 발생하는 데 기여한다.
구조.
콘채는 두꺼운 혈관 및 발기성 선조직 [4]층을 가진 섬모성 호흡기 상피인 가성 원기둥으로 구성되어 있다.콘채는 코구멍에 가로로 위치하고 안쪽과 아래쪽으로 감겨져 코 기도 안으로 들어간다.각각의 쌍은 비중격으로 나누어진 비강 양쪽에 하나의 콩카로 [4]구성되어 있다.
우량한 콘채는 작은 구조로, 신경 말미에 의해 중간 콘채와 연결되어 후구를 보호하는 역할을 한다.후측 체골 부비강 개구부는 상부 미투스 [3]아래에 있습니다.
가운데 콘채가 더 작아요.인간의 경우, 그들은 보통 어린 손가락만큼 깁니다.상악부, 전방 및 중앙 체질 부비강 개구부 위로 아래로 돌출하여 부비강이 가압된 비강에 직접 접촉하지 않도록 완충 역할을 합니다.대부분의 흡입된 공기 흐름은 하등 [3]콩카와 중간 고기 사이를 이동합니다.
하등 콩채는 가장 크고, 사람의 검지 손가락만큼 길 수 있으며,[3] 공기 흐름 방향, 가습, 가열, 코로 흡입되는 공기 여과의 대부분을 담당한다.
하위 콘채는 하위 콘카가 차지하는 기도 공간의 총량을 추정하는 하위 콘카 분류 체계(하위 탁도 분류 체계)에 따라 1~4등급으로 분류된다.1등급은 기도의 0~25%, 2등급은 기도의 26~50%, 3등급은 기도의 51~75%, 4등급은 [5]기도의 76~100%입니다.
때로는 우량한 콘채보다 우량한 콘채가 있다.존재하는 경우, 이들은 보통 작은 볏의 형태를 취합니다.
기능.
콘채는 코의 점막 조직의 대부분을 구성하고 기능적인 호흡에 필요합니다.그들은 공기 흐름 압력과 온도 감지 신경 수용체로 풍부해져, 날씨 조건과 신체의 변화하는 요구에 [4]반응하여 코 막힘과 소화불량의 엄청난 발기 능력을 가능하게 합니다.게다가, 발기 조직은 종종 코 주기라고 불리는 부분 충혈과 탈혈의 눈에 띄지 않는 순환을 겪습니다.코 점막, 특히 콘채의 정맥총으로의 혈액 흐름은 익상구개 신경절에 의해 조절되고 코 안의 공기를 가열 또는 냉각시킵니다.
비폐 및 비흉부반사는 흡입의 깊이를 통해 호흡 메커니즘을 조절합니다.공기의 흐름, 코 안의 공기의 압력 및 공기의 질에 의해 유발되는 코 점막으로부터의 임펄스는 삼차신경에 의해 뇌간 호흡중심으로 전달되고 생성된 반응은 기관지, 늑간근 및 횡격막으로 전달된다.
콘채는 또한 코를 통해 흡입된 공기를 여과, 가열, 가습하는 역할을 한다.이 세 가지 중, 여과는 대부분 점액과 섬모 같은 다른 보다 효과적인 방법으로 이루어진다.공기가 콘채 위를 통과할 때 32–34°C(89–93°F)로 가열되고 가습(최대 98%의 수분 포화)된 후 [4]여과됩니다.
면역학적 역할
콩채의 발기성 조직(또는 라미나 프로프리아)을 덮고 있는 호흡 상피는 신체의 첫 번째 면역 방어선에 중요한 역할을 한다.호흡기 상피는 부분적으로 점액을 생성하는 고블렛 세포로 구성되어 있다.이 분비된 점액은 비강을 덮고 있으며, 공기 전달 입자가 2~3마이크로미터보다 커지도록 함으로서 필터 역할을 한다.호흡기 상피는 또한 림프계를 위한 접근 수단으로서 신체를 바이러스나 [3]박테리아에 의해 감염되는 것으로부터 보호하는 역할을 한다.
냄새
무엇보다도, 콘채는 섬세한 후각 상피를 보존하기 위해 필요한 습도를 제공하며, 이는 후각 수용체를 건강하고 경각심을 유지하는데 필요합니다.상피층이 건조해지거나 자극을 받으면 기능이 정지될 수 있습니다.이것은 보통 일시적인 상태이지만 시간이 지남에 따라 만성 무호흡증으로 [4]이어질 수 있다.또한 터비네이트는 코 안쪽의 표면적을 증가시켜 코 안쪽의 최대 점막 표면에 걸쳐 기류를 유도 및 편향시킴으로써 영감을 받은 공기를 추진시킬 수 있습니다.이것은 콘채에 의해 제공되는 습도와 여과와 결합되어 후각 신경 수용체가 [3]위치한 더 높고 매우 좁은 비강 기도의 영역으로 향기 분자를 더 많이 운반하는 데 도움을 줍니다.
우량 콘채는 요골판(코와 뇌를 분리하는 다공질 뼈판)을 관통하는 신경 축삭을 완전히 덮고 보호한다.중간 콘채의 일부 영역 또한 후구에 의해 내부 자극을 받는다.세 쌍의 콩채는 모두 삼차 신경(또는 다섯 번째 두개골 신경)[4]을 통해 통증과 온도 수용체에 의해 자극된다.연구는 이러한 신경 종말과 후각 수용체의 활성화 사이에 강한 연관성이 있다는 것을 보여주었지만, 과학은 아직 이러한 상호작용을 완전히 설명하지 못했다.
임상적 의의
기능 장애
임상적으로 터비네이트라고 불리는 크고 부은 콩채는 코 호흡의 차단을 초래할 수 있습니다.알레르기, 환경 자극 물질에 노출되거나 부비강 내의 지속적인 염증이 탁한 붓기를 초래할 수 있습니다.비중격의 변형은 또한 터빈산염을 [6]증가시킬 수 있다.
근본적인 알레르기 또는 자극제를 치료하면 탁한 붓기를 줄일 수 있습니다.해결되지 않는 경우 또는 편중격의 치료를 위해 탁상수술이 필요할 수 있습니다.
수술.
터비네이트 절제술은 터비네이트를 줄이거나 제거하는 수술이다.수면성형술이라고도 하는 양극성 고주파 절제술, 순수한 열의 사용에 의한 감소, 탁분할 등의 다양한 기술이 있습니다.
절개 시에는 터빈산염이 호흡에 필수적이기 때문에 소량의 탁조직만 제거된다.하등 또는 중간 터빈산염의 감소 위험에는 빈 코 [6]증후군이 포함된다.하우저 박사: "이는 특히 내부 비강 [7]판막에서 중요한 역할을 하기 때문에 전방 하방 터보(IT) 절제술의 경우에 해당됩니다."
콩차불소증은 중간 탁도의 비정상적인 기화작용으로 부비동 오스티아의 정상적인 환기를 방해할 수 있으며 재발성 부비동염을 일으킬 수 있다.
기타 동물

일반적으로 동물에서, 코 콘채는 비강에 위치한 얇은 뼈나 연골로 이루어진 복잡한 구조이다.여기에는 두 가지 기능을 수행할 수 있는 점막이 있습니다.공기 중의 화학물질을 흡수할 수 있는 면적을 늘려 후각을 향상시키고, 흡입한 공기를 따뜻하게 하고 습하게 하며, 내쉬는 공기에서 열과 수분을 추출하여 폐의 탈수를 예방할 수 있다.후각 터비네이트는 모든 살아있는 [citation needed]네발동물에서 발견되고 호흡 터비네이트는 대부분의 포유류와 조류에서 발견됩니다.
호흡 터빈산염을 가진 동물은 폐를 건조시키지 않고 호흡할 수 있고, 결과적으로 [8]신진대사가 빨라진다.예를 들어, 에뮤가 숨을 내쉴 때, 에뮤의 코는 공기 중의 수분을 응축시켜 [9]재사용을 위해 흡수합니다.개와 다른 개들은 잘 발달된 코 터빈산염을 [10]가지고 있다.이러한 터빈산염은 역류 열 교환 시스템에서 [10]상완골(상완골에 위치한 터빈산염) 표면의 작은 동맥과 정맥 사이의 열 교환을 허용합니다.개는 고양이의 매복 포식과는 대조적으로 오랫동안 쫓을 수 있고, 이 복잡한 터비네이트는 [10]이것을 가능하게 하는데 중요한 역할을 합니다.이와 같은 복잡한 혼탁 구조는 건조한 [11]환경에서 물을 보존하는 데 도움이 됩니다.개에서 잘 발달된 터비네이트의 수분 보존과 온도 조절 능력은 개들이 가혹한 북극 환경과 북부 유라시아와 북미의 추운 지역에서 생존할 수 있게 해준 중요한 적응이었을 수 있습니다.건조하고 매우 [11]춥다.
파충류와 더 원시적인 시냅시드는 [12]건조를 막기보다는 냄새를 감지하는 데 관여하는 후각 터빈산염을 가지고 있다.포유류의 맥실로터베이트는 수분을 모으기 위해 공기 흐름의 경로에 위치하는 반면, 포유류와 파충류 모두의 감각 터빈산염은 [13]공기의 흐름으로부터 멀리 코의 뒤쪽과 위에 위치해요.글라노수쿠스는 비강에서 낮은 곳에 위치한 융기를 가지고 있는데, 이는 공기 흐름의 직접적인 경로에 있는 최대 자극제를 가지고 있었음을 나타냅니다.맥실로터베이트는 매우 얇거나 연골이기 때문에 보존되지 않았을 수 있다.또한 이러한 융기들이 [14]터비네이트가 아닌 후각 상피와 연관되어 있을 가능성도 제기되었다.그럼에도 불구하고, 맥실로터르베이트의 존재 가능성은 글라노수쿠스가 코의 통로를 건조시키지 않고 빠르게 숨을 쉴 수 있었고, 따라서 [8][12][14]내온증이었을 수 있다는 것을 암시한다.
비강 터빈산염의 뼈는 매우 연약하고 화석으로 살아남는 경우는 드물다.특히 화석 [15]조류에서 발견된 것은 없다.그러나 일부 화석에는 그들의 존재에 대한 간접적인 증거가 있다.호흡 터비네이트를 지탱하는 것과 같은 기본적인 능선은 트리낙소돈과 디아데모돈과 같은 진보된 트라이아스기 신노돈에서 발견되었습니다.이것은 그들이 꽤 높은 신진대사율을 [16][17][18][19]가지고 있었을지도 모른다는 것을 암시한다.고생물학자 존 루벤과 다른 사람들은 공룡에서 코 터빈산염의 증거가 발견되지 않았다고 주장했다.그들이 조사한 모든 공룡들은 코 터빈산염을 수용하기엔 너무 좁고 짧다고 주장하는 코 통로를 가지고 있었다. 그래서 공룡들은 쉬는 동안 포유동물이나 새와 같은 신진대사에 필요한 호흡수를 유지하지 못했을 것이다. 왜냐하면 그들의 폐가 [13][20][21]말라버렸을 것이기 때문이다.그러나 이 주장에 대한 반대 의견도 제기되고 있다.비강 터빈산염은 쥐목, 프로셀라리목, 매목과 같은 일부 조류에 없거나 매우 작습니다.개미핥기, 박쥐, 코끼리, 고래, 그리고 대부분의 영장류와 같은 포유동물들에서는 그들은 또한 없거나 매우 작습니다, 비록 이 동물들은 완전히 흡열하고 어떤 경우에는 매우 [22][23][24][25]활동적입니다.게다가, 골화된 탁골은 강룡과 [26]공룡 사이차니아에서 확인되었다.
「 」를 참조해 주세요.
기타 이미지
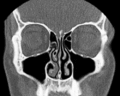
코콘채:차단/프리
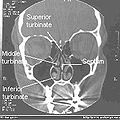
일반 코 CT 전면 단면

비강의 관상부.

우측 비강 기도

코콘채
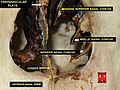
코콩차






메모들
- ^ Maynard, Robert Lewis; Downes, Noel (2019). "Nasal Cavity". Anatomy and Histology of the Laboratory Rat in Toxicology and Biomedical Research. Elsevier. pp. 109–121. doi:10.1016/b978-0-12-811837-5.00010-1. ISBN 978-0-12-811837-5.
In man, the three conchae define the meatuses: Inferior meatus: between inferior concha/turbinate and floor of nasal cavity; Middle meatus: between middle concha/turbinate and inferior concha; Superior meatus: between superior concha/turbinate and middle concha.
- ^ Carlson, Bruce M. (2019). "The Respiratory System". The Human Body. Elsevier. pp. 303–319. doi:10.1016/b978-0-12-804254-0.00011-9. ISBN 978-0-12-804254-0.
- ^ a b c d e f 인체 해부학 2013-01-21 Wayback Machine Gray, Henry(1918) 비강에서 보관된 인체 해부학.
- ^ a b c d e f 터보네이트 기능 장애: 하등 터비네이트가 비기도 폐색에서 차지하는 역할에 집중하세요.2006-06-22 Wayback Machine S.S. Reddy 등에서의 아카이브이비인후과 그랜드 라운드 프레젠테이션, UTB
- ^ Camacho, M.; Zaghi, S.; Certal, V.; Abdullatif, J.; Means, C.; Acevedo, J.; Liu, S.; Brietzke, S. E.; Kushida, C. A.; Capasso, R. (2014). "Inferior Turbinate classification system, grades 1 to 4: Development and validation study". The Laryngoscope. 125: 296–302. doi:10.1002/lary.24923.
- ^ a b Sinus Info Center에서 하부 터보네이트 감소/제거.
- ^ 하우스 SM코막힘 증후군의 외과적 치료.이비인후과 두경부외과의 아카이브> 제133권 (제9호) 2007년 9월 : 858-863.
- ^ a b Zimmer, C. (1994). "The Importance of Noses". Discover. 15 (8).
- ^ Maloney, S. K.; Dawson, T. J. (1998). "Ventilatory accommodation of oxygen demand and respiratory water loss in a large bird, the Emu (Dromaius novaehollandiae), and a re-examination of ventilatory allometry for birds". Physiological Zoology. 71 (6): 712–719. doi:10.1086/515997. PMID 9798259.
- ^ a b c 왕(2008) 페이지 88.
- ^ a b 왕(2008) 페이지 87.
- ^ a b Hillenius, W.J. (1994). "Turbinates in therapsids: Evidence for Late Permian origins of mammalian endothermy". Evolution. 48 (2): 207–229. doi:10.2307/2410089. JSTOR 2410089. PMID 28568303.
- ^ a b Ruben, J.A.; Jones, T.D. (2000). "Selective factors associated with the origin of fur and feathers". American Zoologist. 40 (4): 585–596. doi:10.1093/icb/40.4.585.
- ^ a b Kemp, T.S. (2006). "The origin of mammalian endothermy: a paradigm for the evolution of complex biological structure". Zoological Journal of the Linnean Society. 147 (4): 473–488. doi:10.1111/j.1096-3642.2006.00226.x.
- ^ Witmer, L.M. (August 2001). "Nostril Position in Dinosaurs and Other Vertebrates and Its Significance for Nasal Function". Science. 293 (5531): 850–853. CiteSeerX 10.1.1.629.1744. doi:10.1126/science.1062681. PMID 11486085.
- ^ Brink, A.S. (1955). "A study on the skeleton of Diademodon". Palaeontologia Africana. 3: 3–39.
- ^ Kemp, T.S. (1982). Mammal-like reptiles and the origin of mammals. London: Academic Press. p. 363. ISBN 978-0-12-404120-2.
- ^ Hillenius, W.H. (1992). "The evolution of nasal turbinates and mammalian endothermy". Paleobiology. 18 (1): 17–29. doi:10.1017/S0094837300012197. JSTOR 2400978.
- ^ Ruben, J. (1995). "The evolution of endothermy in mammals and birds: from physiology to fossils". Annual Review of Physiology. 57: 69–95. doi:10.1146/annurev.ph.57.030195.000441. PMID 7778882.
- ^ Ruben, J.A., Jones, T.D., Geist, N.R. and Hillenius, W. J. (November 1997). "Lung structure and ventilation in theropod dinosaurs and early birds". Science. 278 (5341): 1267–1270. Bibcode:1997Sci...278.1267R. doi:10.1126/science.278.5341.1267.
{{cite journal}}: CS1 maint: 여러 이름: 작성자 목록(링크) - ^ Ruben, J.A., Hillenius, W.J., Geist, N.R., Leitch, A., Jones, T.D., Currie, P.J., Horner, J.R., and Espe, G. (August 1996). "The metabolic status of some Late Cretaceous dinosaurs". Science. 273 (5279): 1204–1207. Bibcode:1996Sci...273.1204R. doi:10.1126/science.273.5279.1204.
{{cite journal}}: CS1 maint: 여러 이름: 작성자 목록(링크) - ^ Bang, B.G. (1966). "The olfactory apparatus of Procellariiformes". Acta Anatomica. 65 (1): 391–415. doi:10.1159/000142884. PMID 5965973.
- ^ Bang, B.G. (1971). "Functional anatomy of the olfactory system in 23 orders of birds". Acta Anatomica. 79. 79: 1–76. doi:10.1159/isbn.978-3-318-01866-0. PMID 5133493.
- ^ Scott, J.H. (1954). "Heat regulating function of the nasal mucous membrane". Journal of Larynology and Otology. 68 (5): 308–317. doi:10.1017/S0022215100049707. PMID 13163588.
- ^ Coulombe, H.N., Sam H. Ridgway, S.H., and Evans, W.E. (1965). "Respiratory water exchange in two species of porpoise". Science. 149 (3679): 86–88. Bibcode:1965Sci...149...86C. doi:10.1126/science.149.3679.86. PMID 17737801.
{{cite journal}}: CS1 maint: 여러 이름: 작성자 목록(링크) - ^ Maryańska, T. (1977). "Ankylosauridae (Dinosauria) from Mongolia". Palaeontologia Polonica. 37: 85–151.
레퍼런스
- 왕샤오밍(2008) 개: 그들의 화석 친척과 진화사 컬럼비아 대학 출판부.ISBN 9780231509435.


