세이 무리아
Seymouria| 세이 무리아 시간 범위: 초기 페름기 PreꞒ Ꞓ OSDCPTJKPg N. | |
|---|---|
 | |
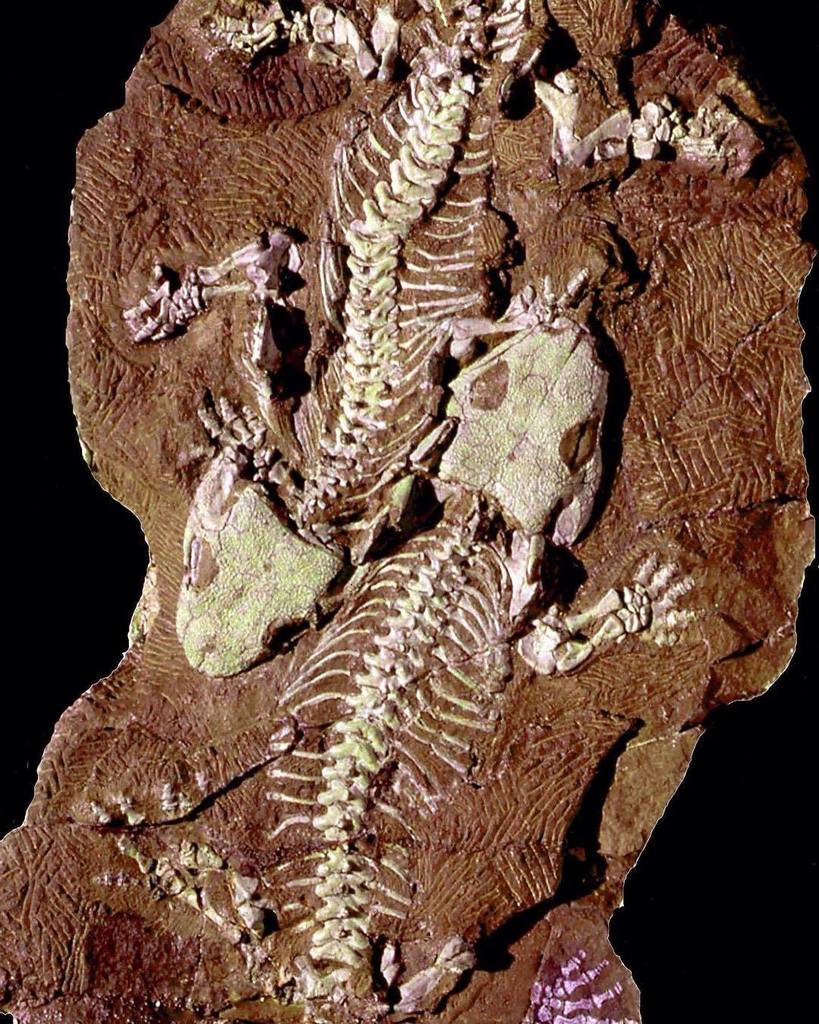
| 세이 무리아 baylorensis의 화석 국립 자연사 박물관에 전시된다. | |
| 과학적 분류 | |
| 왕국: | 동물계 |
| Phylum: | 척색 동물문. |
| 주문: | †Seymouriamorpha |
| 가족: | †Seymouriidae |
| 속: | †세이 무리아 Broili, 1904 |
| 종 | |
| |
| 동의어 해설 | |
| |
Seymouriamorph의 초기 북미와 유럽의 페름기에서 세이 무리아는 멸종된 유인원 속이다.[1] 비록 그들은 양서류(생물학적 의미에서), 세이 무리아 육상 생활 사실은 많은 파충류의 features—so 많은, 세이 무리아 처음 생각은 원시적인 파충류와 well-adapted 있었다.[2][3] 그것은 주로 두종, 세이 무리아 baylorensis[4]과 세이 무리아 sanjuanensis에서 알려져 있다.[5] 비록 그것의 화석은 단지 텍사스에서 발견되는 유형의 종, S.baylorensis, 더 전문화된 견고하다.[6] 반면에, 세이 무리아 sanjuanensis 더 자주 광범위하게 양은 풍부하다. 6뼈의 블록이 커틀러 형성 뉴 Mexico,[7]에서 발견되고 Tambach 형성, 독일은 서로 옆에 누워 있는 화석화된 것을 완전히 다 큰 뼈의 쌍을 포함해 이 작은 종들 여러 잘 보존된 화석에서 알려져 있다.[8]
For the first half of the 20th century, Seymouria was considered one of the oldest and most "primitive" known reptiles.[2][3] 고생물 학자들은 어떻게 일반적인 몸의 형태 captorhinids 것과 같은 초기 파충류의 닮고, 팔다리, 엉덩이, 두개골의 특정 각색 또한 초기 파충류의, 또는 멸종된 현대 양서류의 어떤 종들 당시에 알려져보다는 비슷한 점에 주목했다. 그strongly-built 사지와 등뼈 또한 그 아이디어는 세이 무리아 주로 물에서 아주 적은 시간을 보내는 지상파가 지지했다.[9] 그러나, 1950년대에는, 화석화된 올챙이 Discosauriscus는 가까운 세이 무리아 그 단체 내의 Seymouriamorpha 친척이었다에서 발견되었다. 이것은seymouriamorphs(세이 무리아 포함), 따라서 세이 무리아지 않는 진정한 파충류 하는 것이 아니라 밀접하게 파충류가 관련이 있다고 했고 양서류가 물에서 살고 있는 유충기 것을 보여 준다.[10] 비록 더 이상 가장 원시적인 파충류를 고려했을 때, 세이 무리아는 여전히 중요한 과도기적 화석을 넘어선 amniotes(파충류, 포유 동물, 새들)을 부여하는 양막성 알의 진화에reptile-like 계획 골격 기능의 획득입니다.[1]
역사

세이모우리아의 화석은 텍사스 주 베일러 카운티의 세이모어 마을 근처에서 처음 발견되었다(시모우리아 베이로렌시스, 마을과 카운티 모두를 지칭하는 형태종류의 이름은 세이모우리아 베이로렌시스(Seimouria baylorensis)이다. 처음 발견된 화석은 C가 수집한 개인들의 무리였다.1882년 H. 스턴버그. 그러나 이러한 화석들은 1930년까지 제대로 준비되지 않고 세이모우리아로 확인될 것이다.[11]
전 세계의 다양한 고생물학자들은 19세기 말과 20세기 초에 그들 자신의 세이모우리아 베이로렌시스 화석을 발견했다. 세이모우리아라고 명시적으로 이름 붙여진 최초의 화석은 불완전한 두개골의 한 쌍이었는데, 그 중 하나는 몇 개의 가슴과 척추 원소로 보존되어 있었다. 이 화석들은 1904년 독일의 고생물학자 페르디난드 브로일리에 의해 설명되었으며, 현재 뮌헨에 저장되어 있다.[4] 미국의 고생물학자 S.W. 윌리스톤은 이후 1911년에 거의 완전한 골격을 묘사했고, 그가 최근에 단편적인 팔다리와 척추뼈에서 이름을 붙인 세손인 "Desmospontdylus anormus"는 세이모우리아의 청소년이나 심지어 배아적인 개인을 대표할 가능성이 있다고 언급했다.[2]
마찬가지로 영국의 고생물학자 D.M.S. 왓슨은 1918년에 유명한 에드워드 드링거 코프가 1896년에 지명한 의심스러운 세대인 콘노덱테스가 세이모우리아와 동의어였을 가능성이 높다고 지적했다.[3] 로버트 브룸(1922년)은 그 이름이 먼저 발표되었기 때문에 속주를 코노덱테스(Conodectes)라고 불러야 한다고 주장했지만, 알프레드 로머(1928년)는 세이모우리아라는 이름이 과학계 내에서 너무 인기가 많아 대체할 수 없다고 지적하며 반대했다.[12][9] 이 시기 동안 세이모우리아는 일반적으로 매우 초기 파충류로 보였는데, 이 파충류는 체격이 건장한 다른 많은 페름 파충류나 파충류 같은 테트라포드도 포함시킨 "코틸로룡"이라고 알려진 진화 등급의 일부였다. 그러나 많은 고생물학자들은 의심의 여지 없이 "라비린토돈트" 양서류인 색전체와 많은 유사점을 지적하며 파충류에 대한 충성을 확신하지 못했다. 파충류(즉, 다른 "코틸로룡")와 양서류(즉, 엠볼로메레스)의 이러한 특징의 조합은 세이모우리아가 두 집단 사이의 진화적 전환의 중심이었다는 증거였다. 그럼에도 불구하고, 그것의 생물학에 대해 그것이 정말로 어떤 그룹의 일부분인지 결론지을 만큼 충분히 알려져 있지 않았다. 빗자루(1922년)[12]와 러시아의 고생물학자 피터 수스킨(1925년)[13]은 수륙양용군 사이의 배치를 지지했지만, 이 무렵의 대부분의 연구에서는 극히 "원초적인" 파충류라고 잠정적으로 간주했다; 이것들은 테오도르 E가 발표한 이 종에 언급된 물질의 포괄적 재방사를 포함하고 있었다. 1939년 화이트.[11]

그러나 세이모우리아가 생물학적으로 파충류가 아니라는 간접 증거가 나오기 시작했다. 1940년대까지, 몇몇 제네라가 세이무리아모폴라 그룹의 일부로서 세이모우리아와 연결되었다. 코틀라시아와 같은 일부 시모우리아모르프들은 수생 습관의 증거를 가지고 있었고, 심지어 세이모우리아 자체도 가끔 물속에서만 사용할 수 있는 수평선, 감각 구조를 가지고 있다고 주장되어 왔다.[11] 왓슨(1942)[14]과 로머(1947)는 [15]각각 세이모우리아의 분류에 대해 입장을 번복해 파충류보다는 양서류에 속하게 했다. 아마도 가장 충격적인 증거는 1952년 체코의 고생물학자 즈데노크 슈피나르가 선구리아모프 디스코사우루스(Discosauriscus)의 청소년 화석에 보존된 아가미를 보고했을 때 나왔을 것이다. 이것은 생물학적으로 말하면 시무리아모르프들이 유충기를 가지고 있고, 따라서 양서류라는 것을 분명히 증명했다.[10] 그럼에도 불구하고 세이모우리아와 파충류의 유사성이 높은 것은 세이모우아모프들이 오히려 양수류의 조상에 가깝다는 생각을 뒷받침해 주었다.

1966년 피터 폴 본은 시모우리아가 유타의 오르간 록 셰일로부터 남아있다고 묘사했다. 두개골의 일종인 이 유골들은 새로운 종인 세이모우리아 산후아넨시스를 상징한다.[5] 이 종의 화석은 세이모우리아 베이로렌시스보다 풍부하고 널리 퍼져 있는 것으로 밝혀졌다. 나중에 폴 E에 의해 몇 종 더 이름이 붙여졌다. 올슨, 비록 그들의 타당성이 S. 산후아넨시스의 그것보다 더 의심스럽기는 했지만. 예를 들어, 오클라호마의 치카샤 형성의 거의 완전한 몰골로 알려진 세이모우리아 아길리스(Olson, 1980년)는 미셸 로린과 로버트 R에 의해 재지정되었다. 2001년에 낙하산 매크로레터에게 레이스를 보내라.[16] 텍사스에서 발견된 뇌사 사건에서 1년 전에 묘사된 세이모우리아 그란디스는 다른 어떤 테트라포드에도 다시 언급되지 않았지만 여전히 잘 알려져 있지 않다. 랭스턴(1963)은 캐나다 동부 해안 프린스 에드워드 섬의 페미니아 퇴적물에서 대퇴골과 구별할 수 없는 대퇴골이 발견됐다고 보고했다.[17] 설리번&레이즈(1999년)가 처음 설명한 대로 오클라호마의 리차드 스퍼 채석장에서도 세이모우리아와 같은 골격 유적이 알려져 있다.[18]
S.산후아넨시스 해골 6개가 들어 있는 침전물 덩어리가 베르만, 레이즈 & 에베르스(1987년)가 설명한 뉴멕시코의 커틀러 형성에서 발견되었다.[7] 1993년 베르만앤마텐스는 독일 탐바흐 형성의 S.산후아넨시스 화석을 묘사하면서 최초의 세이모우리아 유적이 북아메리카 밖에 남아 있다고 보고했다.[17] 탐바치 포메이션은 또한 커틀러 포메이션과 비슷한 품질의 S. 산후아넨시스 화석을 생산했다. 예를 들어 2000년 베르만과 그의 동료들은 "탐바흐 연인들"에 대해 설명했는데, S.산주아넨시스의 완전하고 완전하게 표현된 두 개의 해골은 서로 옆에 놓여 있었다(그들이 짝짓기 중 죽은 커플인지 여부는 알 수 없지만).[8] 탐바흐 형성은 또한 발달적으로 가장 어린 것으로 알려진 세이모우리아 화석을 생산해 주로 청소년들로부터 알려진 디스코사우리스쿠스와의 비교를 도왔다.[19]
설명
세이모우리아 개인은 큰 머리와 짧은 목, 육중한 팔다리와 넓은 발을 가진 튼튼한 체격의 동물이었다.[2][15] 그것들은 꽤 작았고, 겨우 2피트(60cm) 길이에 불과했다. 그 두개골은 위에서 보았을 때 상자 모양의 것이었고 대략 삼각형 모양이었지만, 그것은 대부분의 다른 시무리아모르프들보다 낮고 길었다. 척추뼈에는 넓고 부어오른 신경 아치(척추 위 부분)가 있었다. 전체적으로 몸 모양은 현대의 파충류나 파충류처럼 생긴 포충류, 디아데토모르프, 파라렙타이드와 비슷했다. 집합적으로 이러한 종류의 동물들은 쇄골(자연적인 관계 기반 그룹)을 형성하지는 않지만 과거에 "코틸로룡"이라고 일컬어 왔다.
스컬

두개골은 많은 더 작은 접시 모양의 뼈들로 구성되어 있었다. 세이모우리아에 존재하는 두개골 뼈의 구성은 훨씬 더 오래된 테트라포드 및 테트라포드 친척들의 그것과 매우 유사했다. 예를 들어, 그것은 Ventastega나 embolomeres와 같은 동물들에게 존재하는 플레시오모르픽("원발적") 조건인 임시간 뼈를 유지한다.[15] 두개골 뼈는 고대 양서류와 캡토리니드 파충류에서 흔히 볼 수 있듯이 질감이 매우 강했다. 게다가 두개골의 뒷부분에는 옆구리를 따라 길게 뻗은 커다란 절개가 있었다. 이 절개는 오치(otic notch)라고 불렸으며, 같은 일반 부위의 유사한 절개는 대부분의 고생대 양서류에서 흔히 볼 수 있으나('labyrinthodonts, 가끔 부르는 것처럼 labyrinthonts') 양서류에서는 알 수 없다. oic 노치의 하단 가장자리는 편평골에 의해 형성되었고, 위쪽 가장자리는 suppratemporal과 tableular alges(otic flanges)의 하강된 flanges에 의해 형성되었다. 표에는 두개골 후면에서 볼 수 있는 두 번째 하강 플랜지가 있는데, 이 플랜지(후두 플랜지라고 알려져 있음)는 뇌케이스와 연결되고 두개골 측면 사이의 공간을 부분적으로 가렸다. octic과 후두엽 플랜지의 발달은 세이모우리아(특히 S. baylorenis)에서 다른 어떤 시모우리아모르프보다 더 크다.[6]
두개골의 감각 기관은 또한 일련의 독특한 특징들로 언급될 만하다. 궤적(눈구멍)은 비록 청소년기의 코에 조금 더 가까웠지만 두개골의 길이를 중간 정도 낮추고 있었다. 그것들은 날카로운 앞 가장자리를 가진 다른 시모우리모형의 원형 궤도에 비해 더 치명적이었다.[11] 몇몇 저자들은 시모우리아의 몇몇 표본이 궤도를 둘러싼 뼈와 oic 노치 앞에 있는 불분명한 홈을 가지고 있다고 지적했다. 이 홈들은 세이모우리아 유충기 추정 등 수생동물에게 유용한 압력 감지 기관의 거미줄인 가로선 시스템의 잔해일 가능성이 높다.[11][15][5] 많은 표본들은 심지어 청소년도 [7][6]아닌 그들의 측면선의 잔존물을 가지고 있지 않다.[19] 대략 두정골의 중간에는 소나무 포아멘으로 알려진 작은 구멍이 있었는데, 두정골이라고 알려진 감각 기관을 가지고 있었을지도 모른다. 소나무 포아멘은 다른 시모우리아모르프보다 세이모우리아에서 작다.[8]
두개골의 뇌통과 벽 사이에 놓여 있는 막대 모양의 뼈인 그 가시가 가늘어졌다. 그것은 뇌케이스를 otic 노치의 위쪽 가장자리에 연결시켰고, 아마도 otic 노치 안에 놓여 있는 tympuminum (eardrum)에 의해 수신되는 진동의 통로 역할을 했을 것이다. 이런 방법으로 외부 세계에서 뇌로 소리를 전달할 수 있었다. 사각형의 구성은 비암니오테 테트라포드(non amniote tetrapod)와 암니오테(amniote) 사이의 중간이다. 한편으로 오트 노치와의 연결은 특이한데, 진정한 파충류나 다른 양수동물들이 오트노치를 잃었기 때문에, 고막과 스태프는 턱관절의 사분골 쪽으로 아래쪽으로 움직일 수밖에 없었다. 반면 세이모우리아의 가냘프고 예민한 구조는 대부분의 비암니오테 테트라포드보다 특화된 것으로 청각에는 쓸모가 없지만 두개골 보강에는 유용한 두꺼운 가마를 가지고 있다.[9] 세이모우리아 베이로렌시스의 내이에는 전정(아래가 아닌) 뒤쪽에 위치한 달팽이관(Cocharle)이 있으며, 앞쪽 반원관(Bony가 아닌 거위) 대정맥(Supraoccipital)으로 둘러싸인 것 같다. 이러한 특징들은 진짜 파충류나 시냅스보다 더 원시적이다.[20]
입천장(입천장)은 암니오테와 비암니오테 테트라포드 둘 다와 어느 정도 유사성이 있었다. 한편으로는 '라비린토돈트' 양서류의 특징처럼 미로 같은 내부 에나멜 접힘으로 몇 개의 고립된 송곳니를 간직하고 있었다. 반면 입 앞쪽에 있는 보머 뼈는 상당히 좁았고, 인접한 초아내(비강에서 입까지 이어지는 구멍)는 양수처럼 크고 가깝게 붙어 있었다. 미각은 일반적으로 고체 뼈로, 길고 얇은 컬트리폼 과정(뇌경부의 앞쪽 가지)으로 분리되어 있을 뿐이며, 잔존성 인터테리오이드 빈점(중간선을 따라 한 쌍의 구멍)만 있다. 고립된 송곳니와는 별개로, 입천장은 또한 피테리고이드 뼈의 뒷부분에서 방사되는 작은 틀니로 덮여 있다.[21] 세이모우리아(Seimouria)는 프테리고이드(공식적으로 가로 플랜지라고 알려져 있음)의 프롱과 같은 가지와 더불어 프테리고이드와는 별개의 에피페리고이드 뼈가 있는 등 미각의 몇 가지 암모테 같은 특징을 가지고 있다. 그러나 이러한 특성은 여러 비암니오테 테트라포드에서 관찰되어 왔기 때문에 암니오테로서의 지위를 나타내는 것은 아니다.[6]
아래턱은 몇 가지 플레시오모르픽 특성을 유지했다. 예를 들어, 하악골의 안쪽 가장자리는 세 개의 코로노이드 뼈를 가지고 있었다.[11] 하악골은 또한 2005년 커틀러 형성 표본 중 하나를 재조사하는 동안만 이 특징이 확인되었지만 메켈리안 페네스트라라고 알려진 안쪽 가장자리를 따라 적어도 하나의 큰 구멍을 유지했다.[21] 이 특성들 중 어느 것도 양수에서는 흔한 것이 아니다. 이 뇌사에는 다양한 테트라포도모르프들과 공통적으로 특징들이 모자이크되어 있었다. 뇌케이스 측면의 홈과 신경 개구부의 체계는 생선의 메갈리치스와 유달리 유사했으며, 치석기층은 또 다른 플레시오모르픽 기능이다. 그러나 내경동맥은 뼈 복합체 후미 부근의 뇌관을 관통하는데, 이는 양수체와 유사한 파생적 특징이다.[11]
후두골

척추기둥은 엉덩이 앞에 총 24개의 척추뼈가 있어 상당히 짧다.[8] 척추뼈는 위장이며, 각 척추골은 흉골(Pleurocentrum)이라고 알려진 더 크고 다소 스풀 모양의 구성 요소와, 중간중간으로 알려진 더 작은 쐐기 모양의(또는 앞쪽에서 초승달 모양의) 성분을 가지고 있다는 것을 의미한다. 흉골 위에 놓여 있는 신경 아치는 흉골 자체보다 약 3배 넓은 넓이의 테이블 모양의 지가포피시스(공동판)를 가진 넓은 구조로 부풀어 있다. 어떤 척추골은 신경 가시가 있으며, 신경 가시가 부분적으로 중하로 세분되어 있는 반면, 다른 척추골은 단면적으로 타원형이다. 등뼈의 갈비뼈는 수평으로 뻗어 신경아치의 중앙과 측면의 두 곳에서 척추에 붙는다.[11] 목은 거의 없고 척추만 몇 개 길다. 첫 번째 목 척추뼈인 아틀라스는 작은 중심과 함께 성숙한 개인에게만 존재하는 흉골의 감소도 가지고 있었다. 아틀란탈 늑골(현재의 경우)은 아틀란트의 중간중간과 후축 척추의 중간중간(암모테와 같이) 사이에 끼여 있었지만, 목의 이 영역의 낮은 뼈 발달은 양모테의 특징적인 아틀라스 축 복합체와 대비된다.[22] 게다가, 이후의 연구에서는 지도책 중간 부분이 왼쪽과 오른쪽 부분으로 나뉘어져서, 양서류 등급의 테트라포드에 더 가깝다는 것을 밝혀냈다.[7] 다른 거의 모든 고생대 테트라포드(암니오테 또는 그 밖의 것)와는 달리 세이모우리아는 다른 시모우리아모형의 얇고 둥근 배비늘조차 전혀 비늘의 뼈 잔해물이 부족하다.[6][8]
가슴(어깨) 대들보에는 파충류와 같은 여러 가지 특징이 있다. 예를 들어, 어깨뼈와 코라코이드(각각 어깨 소켓 위와 아래에 놓여 있는 뼈판)는 하나의 큰 어깨뼈가 아니라 별도의 뼈다귀다. 마찬가지로 쇄골 사이는 납작하고 버섯모양으로 길고 얇은 '줄기'가 달려 있었다. 상완골(전완골)은 약간 뒤틀리고 상자 모양의 L자 모양으로 근육 부착을 위한 부위가 넓었다. 이 형태는 "침엽수"라고 설명되어 왔으며, 테트라포드(tetrahedral)에 대해서는 플레시오모르픽이다. 반면에, 유머러스에는 파충류처럼 생긴 적응도 있는데, 그것은 엔테피콘디라 포아멘으로 알려진 구멍이다. 반지름은 대략 모래시계 모양이었다. 얼나는 비슷하지만 육지 테트라포드에서 흔히 볼 수 있지만 수륙양용 또는 수륙양용에서는 드물게 볼 수 있듯이 뚜렷한 올레크라논 공정을 보유하고 있기 때문에 더 길다. 카퍼스는 10개의 뼈를 가지고 있고, 손에는 5개의 튼튼한 손가락이 있다. 손목뼈는 완전히 발달되어 있고 서로 밀접하게 접촉하고 있는데, 이것은 또 다른 불임성의 징후다. 발랑(손가락 뼈)은 손가락 끝을 향해 크기가 작아지는데, 손가락 끝은 각각 발톱 없이 작고 둥근 부분으로 끝난다. 팔랑어 공식(손가락당 팔랑어 수, 엄지손가락부터 새끼손가락까지)은 2-3-4-4-3이다.[8]
두 개의 천골(엉덩이) 척추뼈가 존재했지만, 첫 번째 척추골은 장골(엉덩이 위쪽 칼날)과 접촉하는 크고 튼튼한 늑골을 가지고 있었다.[11][8] 일부 연구에서는 천골 척추가 단 하나밖에 없었다고 주장했는데, 두 번째 천골은 사실 첫 번째 천골보다 짧고 곡선이 더 많은 늑골을 가지고 있기 때문에 첫 번째 천골이라고 한다.[7] 각 장골은 옆으로 볼 때 낮고 눈물방울 모양이며, 엉덩이 전체 밑부분은 아래쪽에서 볼 때 직사각형인 하나의 튼튼한 푸부아찌아찌아찌아찌아찌아찌아찌아찌아찌아찌아찌아찌아찌아찌아찌아찌아찌아찌아찌아찌아찌아찌아찌아찌아찌아찌아찌아 고관절과 어깨 소켓 모두 수평 아래 45도로 방향을 잡았다. 대퇴골은 상완골과 마찬가지로 튼튼하고 경골과 섬유는 반경과 척골과 유사한 모래시계 모양의 뼈다귀가 튼튼하다.[11] 타르수스(앵클)는 이전의 테트라포드(12개)와 암모테(8개 이하) 사이의 중간인 11개의 뼈를 포함한다. 발톱이 다섯 개인 발은 손과 상당히 비슷하며, 팔색조 공식은 2-3-4-5-3이다.[8]
꼬리뼈는 기껏해야 20개 정도밖에 되지 않았다. 꼬리의 밑부분을 지나, 카우달들은 그들의 아랫부분을 따라 뼈로 된 가시들을 얻기 시작한다, 쉐브론이라고 알려져 있다. 이것들은 시료에 따라 세 번째에서 여섯 번째 카우달 근처에서 나타나기 시작한다. 갈비뼈는 처음 5~6개의 꼬리 부분에만 존재한다. 갈비뼈는 꼬리 밑부분에서 길지만 곧 줄어들며 일반적으로 갈비뼈가 나타나는 부분과 같은 부위에 사라진다.[8]
종별차이
세이모우리아 베일로렌시스와 세이모우리아 산후아넨시스는 두개골의 다른 뼈들 사이의 형태와 연결의 몇 가지 차이점을 근거로 서로 구별할 수 있다. 예를 들어, 오치(Otic notch) 위쪽에 있는 뼈의 하강 플랜지('타블라 호른' 또는 '오치 과정'으로 불리기도 한다)는 S. baylorensis에서 S. baylorenis에서 훨씬 더 잘 발달되어 있으며, 두개골 후면을 향해 더 넓게 아래로 뻗어 나가면서 삼각형 모양(옆에서 볼 때)을 얻는다.[8] S.산주아넨시스에서는 후전골은 둔탁하고 쐐기 모양의 봉합을 통해 두정골과 접촉하는 반면, 두 뼈 사이의 연결은 S. 베이로렌시스에서는 완전히 직선이다.

일부 저자들은 S. baylorenis의 postparietals가 S. s. sanjuanensis의 postparietals보다 작다고 주장해 왔지만, S. sanjuanensis의 일부 표본들(예: "탐바치 애호가들")도 postietals를 가지고 있었다. 게다가, "탐바흐 애호가들"은 4각형 주갈뼈를 가지고 있는데, 이것은 S. 산후아넨시스보다는 S. 베일로렌시스 뼈와 더 비슷하다. 이러한 표본에서 두 종의 특징을 결합하면 두 종은 강하게 구별되는 두 개의 진화 경로가 아니라 연속적인 혈통의 일부라는 것을 나타낼 수 있다. 마찬가지로, 두개골 후면의 비율과 관련된 일부 차이는 대부분의 S.산후아넨시스 표본이 그 종의 성인 구성원인 '탐바치 애호가'가 발견되기 이전에 완전히 자라지 않았다는 사실을 보여주는 유물로 간주될 수 있다.[8]
그럼에도 불구하고 두 종 사이에 몇 가지 특징이 여전히 뚜렷하게 구별되고 있다. 눈 앞에 있는 열상골은 S. 베일로렌시스(S. baylorensis)에서 궤도의 앞쪽 가장자리만 차지하고 있다. 그러나 S.산후아넨시스의 표본에는 궤도 아래 작은 거리를 연장하는 열경부의 가지가 있다. S.산주아넨시스에서는 궤도의 후미 가장자리의 상당 부분이 S. 베일로렌시스(Bayorenssis)에서 더 직사각형인 쉐브론 모양의 후두골(Postorbital bone)에 의해 형성된다. S.산주아넨시스(S. sanjuanensis)의 lacrimal과 postorbital의 모양은 다른 symouriamphins의 상태와 밀접하게 일치하는 반면, S. baylorensis의 상태는 더 독특하고 파생된다.[8]
주둥이의 옆면을 이루는 이빨을 가진 맥시야 뼈는 S. 베일로렌시스에서도 뚜렷하게 독특하다. S. 산후아넨시스에서는 맥시야가 낮고, 많은 날카롭고 촘촘히 간격을 두고 있는 이빨이 그 길이를 따라 뻗어 있었다. 이 상태는 다른 시무리아모르프들과 비슷하다. 그러나 S. baylorensis는 콧구멍이 더 크고, 치아는 일반적으로 훨씬 크고, 수가 적고, 크기가 균질하지 않다.[8] 비록 엑토프테리고이드가 S. 베일로렌시스에서는 더 삼각형이고 S. 산후아넨시스에서는 직사각형이지만, 미각은 일반적으로 두 종 사이에 유사하다.[21]
Paleobiology
라이프스타일
로머(1928년)는 세이모우리아 해골의 생물학적 함의를 논한 최초의 저자들 중 한 명이었다. 그는 건장한 팔다리와 넓은 체구의 몸이 무질서하게 뻗은 걸음걸이를 한 튼튼한 육지동물이라는 생각을 뒷받침한다고 주장했다. 그러나 그는 또한 퍼미언 트랙웨이가 이 시기부터의 지상 테트라포드는 배꼽을 잡아 끄는 것이 아니라 오히려 몸을 지상에서 멀리할 수 있을 정도로 강하다는 생각을 대체로 지지하고 있다고 지적했다. 그 무렵의 다른 고생물학자들과 마찬가지로 로머는 세이모우리아는 파충류(혹은 암니오트) 번식 방식을 가지고 있으며, 알이 마른 땅에 놓여 있고, 원소로부터 압류막으로 보호되어 있다고 추측했다.[9]
화이트(1939년)는 생물학적 함의에 대해 상세히 기술했다. 그는 오틱노치의 존재는 두개골 내에 부착할 수 있는 표면적 턱근육의 양을 낮춰 턱의 힘을 감소시킨다는 점에 주목했다. 게다가 이렇게 큰 절개가 있어 두개골은 더욱 깨지기 쉬웠을 것이다. 전체적으로 그는 세이모우리아가 크고 활동적인 먹잇감을 상대할 수 있을 것 같지 않다는 것을 알았다. 그럼에도 불구하고 미각의 근육 부착 부위는 동시대 양서류보다 잘 발달되어 있었다. 흰색은 세이모우리아가 무척추동물, 작은 물고기, 그리고 심지어 어떤 식물 물질을 먹고 사는 육식성 일반주의자였고 잡식성 동물이었다고 추론했다. 그의 계산에 의하면 식인종인지도 모른다.[11]
화이트는 또한 비정상적으로 부어오른 척추뼈에 대해서도 주의를 끌었는데, 이것은 측면(측면-측면) 움직임을 촉진시키지만 등뼈의 비틀림(뒤틀림)은 금지한다. 이것은 세이모우리아가 낮은 팔다리, 그렇지 않았다면 비틀림에 취약했을 넓고 상체 육체를 가지고 있었기 때문에 유익했을 것이다. 이것은 또한 캡토히니드, 디아데토모르프, 그리고 다른 "코틸로룡"에서 이 특성의 존재를 설명할 수 있다. 아마도 척추뼈가 부어오른 것은 나중에 파충류에서 강한 고관절 근육으로 대체될 비틀림을 막기 위한 중간 전략이었을 것이다. 다소 발달되지 않은 세이모우리아의 엉덩이 근육은 이 가설과 일맥상통한다. 그럼에도 불구하고 이러한 척추뼈는 활발한 걸음걸이보다 빠른 속도로 비틀림으로부터 방어하는 데 비효율적이었기 때문에, 시모우리아는 아마도 빨리 움직이는 동물은 아니었을 것이다.[11]
화이트는 세이모리아를 육지에서는 꽤 유능하다고 여겼지만, 몇 가지 다른 생활양식에 대해서도 의논했다. 그는 세이모우리아도 수영을 잘한다고 추정했는데, 이는 그가 이 동물이 현대의 악어들과 비슷한 깊고 강력한 꼬리를 가지고 있다고 추정했기 때문이다. 다만 반수나 수생 포식자에 취약했을 것이고, 서식지 선호로 인해 세이모우리아 화석이 육지 퇴적물에서 더 흔했다고도 언급했다. 베르만 외 (2000년) 탐바흐 형성이 세이모우리아 화석을 보존하면서도 수생동물이 완전히 부족했기 때문에 이 가설을 뒷받침했다. 이들은 또 '탐바흐 애호가들'의 잘 발달된 손목과 발목뼈를 육지애착에 대한 지지로 지적했다. 로머(1928년)[9]와 화이트(1939년)[11]는 앞다리의 강한 근육질에도 불구하고 세이모우리아에서 적응을 파고들 만한 증거를 거의 발견하지 못했다.
성적 이형성
일부 저자들은 세이모우리아에 존재하는 성적 이형성에 찬성한다고 주장했지만, 다른 저자들은 이 가설로 납득이 가지 않는다. 화이트(1939년)는 세이모우리아 베이로렌시스 일부 표본에 세 번째 꼬리 척추뼈에 처음 나타난 쉐브론(꼬리 척추뼈 밑면에 있는 뼈의 가시)이 있고, 다른 표본은 6일에 처음 나타났다고 주장했다. 그는 일부 표본에서 나중에 수세미가 나타난 것은 그들이 내부 성기를 보관할 공간이 더 필요한 남성임을 나타내는 것이라고 가정했다. 이러한 유형의 성차별은 거북이와 악어 모두에게 보고되었다. 그는 이를 근거로 세이모우리아 암컷이 거북이나 악어처럼 육지에서 큰 얄키움 알을 낳는다는 아이디어도 지지했다.[11] 이후 본(1966)은 세이모우리아 산후아넨시스(Seimouria sanjuanensis)에서 쉐브론 획득과 특정 두개골 비율의 상관관계를 발견했고, 그것들 역시 성적 이형성의 예라고 제안했다.[5]
그러나 베르만, 레이즈, 앤 엘베르스(1987)는 화이트(1939년)와 본(1966년)의 방법론을 비판했다. 그들은 화이트의 관찰이 아마도 동물들의 성과는 관련이 없을 것이라고 주장했다. 이것은 커틀러 포메이션 표본들 중 일부가 다섯 번째 꼬리 척추에 처음 나타난 쉐브론을 가지고 있다는 사실에 의해 뒷받침되었다. 비록 성기 크기가 골격에 영향을 미칠 정도로 남성들 사이에서 가변적일 수 있었지만, 화이트가 관찰한 차이는 개인의 골격 변화, 진화적 다양성 또는 성적 이형성과 무관한 다른 요인에 의해 발생한다는 것이 더 유력한 설명이었다. 마찬가지로, 그들은 두개골 비율이 시모우리아 화석에 이형성이 존재한다는 본(1966)의 제안을 지지한다는 데 동의했지만, 화이트의 기준으로 '여성'으로 여겨지는 화석을 이용한 성관계와 어떻게 연관시키는지에 대해서는 동의하지 않았다.[7] 화석화된 유충의 발견은 세이모우리아가 수생유충 단계를 가졌을 가능성이 있다는 것을 보여주었고, 세이모우리아가 육지에 알을 낳았다는 이전의 가설들을 밝혀냈다.[10]
역사학과 발전
오클라호마주 리차드 스퍼스에서 발견된 표본의 역사학적 증거는 세이모우리아의 생물학에 대한 추가 정보를 제공했다. 대퇴골은 수많은 대퇴골 운하를 뚫은 항성 행렬이 특징인 내부 구조를 가진 것으로 밝혀졌다. 성장이 느린 나머지 라인은 불분명하고 간격이 촘촘하지만 뼈 발달 과정에서 성장이 어느 때라도 멈췄다는 증거는 없다. 대부분의 리삼피비아인들과 마찬가지로 중공도 열려 있고 해면골도 소량 있다. 스펀지오사 뼈의 발달은 아켈로마(지상 양서류)에 비해 약간 높지만, 라인수쿠스나 트리메로하키스 같은 수서 양서류에 비해 훨씬 범위가 넓지 않다. 세이모우리아의 척추뼈는 디스코사우리스쿠스에 비해 모양이 탄탄하고 다공성이 높음에도 연골의 양이 적다. 시모우리아(Seimouria)는 우기와 건기의 변동으로 인한 환경적 스트레스로 인해 매우 일찍 변형을 겪은 것으로 추측된다.[23]
참조
- ^ Jump up to: a b Laurin, Michel (January 1, 1996). "Seymouria". Tree of Life Web Project. Paris: Muséum National d'Histoire Naturelle. Retrieved February 24, 2018.
- ^ Jump up to: a b c d Williston, S.W. (1911). "Restoration of Seymouria baylorensis Broili, an American Cotylosaur". The Journal of Geology. 19 (3): 232–237. Bibcode:1911JG.....19..232W. doi:10.1086/621840. S2CID 140644621.
- ^ Jump up to: a b c Watson, D.M.S. (1918). "On Seymouria, the most primitive known reptile". Proceedings of the Zoological Society of London. 88 (3–4): 267–301. doi:10.1111/j.1096-3642.1918.tb02098.x.
- ^ Jump up to: a b Broili, Ferdinand (1904). "Permische Stegocephalen und Reptilien aus Texas". Palaeontographica. 51: 1–48.
- ^ Jump up to: a b c d Vaughn, Peter Paul (May 1966). "Seymouria from the Lower Permian of Southeastern Utah, and Possible Sexual Dimorphism in That Genus". Journal of Paleontology. 40 (3): 603–612. JSTOR 1301745.
- ^ Jump up to: a b c d e Laurin, Michel (13 July 1996). "A Redescription of the Cranial Anatomy of Seymouria baylorensis, the best known Seymouriamorph (Vertebrata: Seymouriamorpha)" (PDF). PaleoBios. 17 (1): 1–16.
- ^ Jump up to: a b c d e f g Berman, David S.; Reisz, Robert R.; Eberth, David A. (1987). "Seymouria sanjuanensis (Amphibia, Batrachosauria) from the Lower Permian Cutler Formation of north-central New Mexico and the occurrence of sexual dimorphism in that genus questioned". Canadian Journal of Earth Sciences. 24 (9): 1769–1784. Bibcode:1987CaJES..24.1769B. doi:10.1139/e87-169.
- ^ Jump up to: a b c d e f g h i j k l m n Berman, David S.; Henrici, Amy C.; Sumida, Stuart S.; Martens, Thomas (2000). "Redescription of Seymouria sanjuanensis (Seymouriamorpha) from the Lower Permian of Germany based on complete, mature specimens with a discussion of paleoecology of the Bromacker locality assemblage". Journal of Vertebrate Paleontology. 20 (2): 253–268. doi:10.1671/0272-4634(2000)020[0253:ROSSSF]2.0.CO;2. JSTOR 4524091.
- ^ Jump up to: a b c d e Romer, Alfred S. (1928). "A Skeletal Model of the Primitive Reptile Seymouria, and the Phylogenetic Position of that type". The Journal of Geology. 36 (3): 248–260. Bibcode:1928JG.....36..248R. doi:10.1086/623510. JSTOR 30060526. S2CID 129513686.
- ^ Jump up to: a b c Bratislava, Jozef Klembara (1995-03-01). "The external gills and ornamentation of skull roof bones of the Lower Permian tetrapod Discosauriscus (Kuhn 1933) with remarks to its ontogeny". Paläontologische Zeitschrift. 69 (1): 265–281. doi:10.1007/BF02985990. S2CID 87658588.
- ^ Jump up to: a b c d e f g h i j k l m n White, T.E. (1939). "Osteology of Seymouria baylorensis Broili". Bulletin of the Museum of Comparative Zoology. 85 (5): 323–409.
- ^ Jump up to: a b Broom, R. (1922). "On the Persistence of the Mesopterygoid in certain Reptilian Skulls". Proceedings of the Zoological Society of London. 1922:pt.1-2 [pp.1-481]: 455–460.
- ^ Sushkin, Peter P. (18 December 1925). "On the representatives of the Seymouriamorpha, supposed primitive Reptiles, from the Upper Permian of Russia, and on their phylogenetic relations". Occasional Papers of the Boston Society of Natural History. 5: 179–181.
- ^ Watson, D. M. S. (1942). "On Permian and Triassic Tetrapods". Geological Magazine. 79 (2): 81–116. Bibcode:1942GeoM...79...81W. doi:10.1017/S0016756800073593. ISSN 1469-5081.
- ^ Jump up to: a b c d Romer, Alfred Sherwood (1947). "Review of the Labyrinthodontia". Bulletin of the Museum of Comparative Zoology at Harvard College. 99 (1): 7–368.
- ^ Laurin, Michel; Reisz, Robert R. (September 2001). "The reptile Macroleter: First vertebrate evidence for correlation of Upper Permian continental strata of North America and Russia" (PDF). Geological Society of America Bulletin. 113 (9): 1229–1233. Bibcode:2001GSAB..113.1229R. doi:10.1130/0016-7606(2001)113<1229:trmfve>2.0.co;2. Archived from the original (PDF) on March 11, 2012.
- ^ Jump up to: a b Berman, David S.; Martens, Thomas (25 February 1993). "First occurrence of Seymouria (Amphibia: Batrachosauria) in the Lower Permian Rotliegend of central Germany". Annals of the Carnegie Museum. 62 (1): 63–79 – via Biostor.
- ^ Sullivan, Corwin; Reisz, Robert R. (1999). "First record of Seymouria (Vertebrata: Seymouriamorpha) from Early Permian fissure fills at Richards Spur, Oklahoma". Canadian Journal of Earth Sciences. 36 (8): 1257–1266. doi:10.1139/e99-035.
- ^ Jump up to: a b Klembara, Jozef; Berman, David S.; Henrici, Amy C.; Cernansky, Andrej; Werneberg, Ralf; Martens, Thomas (30 March 2007). "First description of skull of Lower Permian Seymouria sanjuanensis (Seymouriamorpha: Seymouriidae) at an early juvenile growth stage". Annals of Carnegie Museum. 76 (1): 53–72. doi:10.2992/0097-4463(2007)76[53:FDOSOL]2.0.CO;2.
- ^ Klembara, Jozef; Hain, Miroslav; Ruta, Marcello; Berman, David S.; Pierce, Stephanie E.; Henrici, Amy C. (2019). "Inner ear morphology of diadectomorphs and seymouriamorphs (Tetrapoda) uncovered by high-resolution x-ray microcomputed tomography, and the origin of the amniote crown group". Palaeontology. 63: 131–154. doi:10.1111/pala.12448. ISSN 1475-4983.
- ^ Jump up to: a b c Klembara, Jozef; Berman, David S.; Henrici, Amy C.; Cernansky, Andrej (30 December 2005). "New structures and reconstructions of the skull of the Seymouriamorph Seymouria sanjuanensis Vaughn". Annals of Carnegie Museum. 74 (4): 217–225. doi:10.2992/0097-4463(2005)74[217:NSAROT]2.0.CO;2.
- ^ Sumida, Stuart S.; Lombard, Eric; Berman, David S. (29 May 1992). "Morphology of the atlas-axis complex of the late Palaeozoic tetrapod suborders Diadectomorpha and Seymouriamorpha". Philosophical Transactions of the Royal Society B: Biological Sciences. 336 (1227): 259–273. Bibcode:1992RSPTB.336..259S. doi:10.1098/rstb.1992.0060.
- ^ Bazzana, Kayla D.; Gee, Bryan M.; Bevitt, Joseph J.; Reisz, Robert R. (2020-03-10). "Postcranial anatomy and histology of Seymouria, and the terrestriality of seymouriamorphs". PeerJ. 8: e8698. doi:10.7717/peerj.8698. ISSN 2167-8359. PMC 7069408. PMID 32195050.








{kind=link}
{kind=link}
{kind=link}