피토글로빈
Phytoglobin이 기사의 주요 기고자는 그 주제와 밀접한 관련이 있는 것으로 보인다. (2017년 4월) |
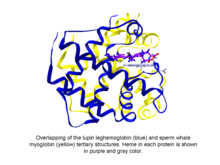
피토글로빈은 글로빈 슈퍼 패밀리로 분류되는 구상 식물(알개와 육지 식물) 단백질로, 헤메(heme), 즉 프로토폴피린 IX-Fe, 보형물 그룹이다.가장 초기에 알려진 피토글로빈은 콩 뿌리 결절의 붉은 색소를 분광학적으로 화학적으로 분석한 후 쿠보가 1939년에 발견한 레그헤모글로빈이다.[1]쿠보의 보고가 있은 후 몇 십 년 후, 베인슈테인과 협력자들에 의해 루핀 피토글로빈(레그헤모글로빈으로 알려진)의 결정체가 이 단백질의 3차 구조와 향유고래 미오글로빈의 3차 구조가 현저하게 유사하다는 것을 밝혀내, 쿠보가 발견한 피토글로빈이 정말로 글로빈과 일치한다는 것을 알 수 있었다.[2]
피토글로빈의 중요한 기능 중 하나는 질소산화물 이산화효소 활성이다.[3]
분배 및 분류
피토글로빈(약칭 피토그브스)은 원시적인 생물학자와 진화된 단조류와 디코트를 포함한 조류와 육지 식물에서 확인되어 식물에서 보편적으로 분포한다.다음과 같이 분류할 수 있다.[3][4]
- 2/2 피토글로빈(트롤드 글로빈 계열, TrHb2 하위 계열)
- Phytogb3: 조류와 육지 식물에서 발견된다.
- 3/3 피토글로빈(미오글로빈 계열)
비레구메 공생 글로빈(SymPhytogbs)[4]은 피토그브1과 피토그브2 사이에 산재해 있다.[3]공생 글로빈은 일반적으로 질소 고정을 수행하는 공생 박테리아에 산소를 공급한다.콩류에서 박테리아는 공포증이 있지만, 몇몇 식물에서는 Frankia가 대신 그 일을 한다.[3]
구조 특성
피토그는 3개의 인트론에 의해 방해를 받는 유전자에 의해 코딩된다. (4개의 인트론 함유 피토그브 유전자가 이끼 Physcomitrella patens에서 검출되었지만)[5]피토그브 유전자의 첫 번째와 세 번째 인트론은 미오글로빈 유전자와 같은 위치에 국부화되어 있는데, 이는 피토그브와 미오글로빈 유전자가 6억년 이상 전에 공통의 조상으로부터 진화했음을 시사한다.[6]피토그브 유전자에 제2의 인트론의 존재는 이론적 분석을 통해 고에 의해 예측되었는데,[7] 이는 마커와 협력자들에 의해 콩 lb 유전자를 복제하고 염기서열화함으로써 더욱 입증되었다.[8]

피토그스는 분자 질량이 ~17~19kDa인 단층 단백질이다.그러나 높은 농도(1mM 이상)에서는 피토그브스가 조광기를 형성할 수 있다.피토그브스 폴리펩타이드 체인은 헤미가 위치한 소수성 주머니를 형성하는 글로빈 폴딩으로 알려진 6~7개의 나선형(A~H라는 글자로 이름 지어짐)의 특정한 배열로 접힌다.피토그브에서는 나선 A, E, F가 헬리컬 B, G, H와 중첩되고 헬리컬 B와 E가 각각 헬리컬 G와 H와 중첩되는 [9][10]3/3과 2/2 폴딩의 두 종류가 확인되었다.

다른 글로빈과 마찬가지로 피토그브스의 헤메-Fe는 근위부에서 히스 아미노산(근위부 히스)에 의해 조정된다.헴-Fe의 원위 부위는 다양한 리간드(산소와 질소산화물 등) 또는 원위(흔히 히스) 아미노산이 점유할 수 있으며, 각각 펜타(Penta-coordinated Phytogbs) 또는 육각(His)에 자리를 준다.Pytogbs2, SymPhytogbs 및 Lbs는 주로 펜타코디네이션이고 Pytogbs1은 육각코디네이션이고 Pytogbs0과 Pytogbs3는 펜타코디네이션과 육각코디네이션의 조합이다.[4]헤메-Fe 조정은 각각 운동 상수 k와on k의off 결과로 리간드 바인딩과 방출 속도를 조절하기 때문에 피토그브(및 기타 글로빈) 기능에 필수적이다.예를 들어 O2(KO2)에 대한 콩 Lb와 쌀 피토그b1의 친화력은 k가on 130, 68 mM−1 s−1, k가off 5.6, 0.038−1 s, KO2(kon/k에off 의한 O-affinity2)가 각각 23, 1800 mM이기−1 때문에 중간이고 매우 높다.[11]이는 콩 Lb가 O-저장2 단백질 또는 –운송 단백질로 기능할 수 있으며, O에2 대한 이 단백질의 높은 친화력은 극히 낮은off k 상수에서 기인하기 때문에 쌀 Phytogb1(및 다른 육각형 Phytogbs)의 기능이 O-운송2 이외의 것이 될 수 있음을 나타낸다.[12]
합성 및 가정된 함수
다른 글로빈과 마찬가지로 펜타 조정 피토그스도 역행적으로 결합하여 O를2 운반한다.결절에서의 lbs의 기능은 1974년 비텐베르크, 애플비 등에 의해 해명되었다.[13]결절에서는 전체 수용성 단백질의 약 30%에 해당하기 때문에 Lbs의 농도가 매우 높다.결절에서 Lbs의 겉보기 기능은 질소 고정을 위해 O의2 재전송 박테로이드의 확산을 촉진하는 것이다.이와 동시에 Lb는 대기 질소를 고정하는 O 민감2 질소 효소의 불활성화를 피하기 위해 낮은 O-수준2(약 10nM)을 유지하는 데 기여한다.[14]
게다가, 피토그bs는 가장 두드러진 질소산화물(NO)인 다른 기체 리간드를 결합하고 NO dioxygenase 활동을 보인다.[15]힐과 협력자들에 의한 지난 15년 동안의 연구는 내생성 NO의 수준은 유전자 변형 옥수수와 알팔파에서의 피토그브스1의 농도에 따라 달라진다는 것을 보여주었다.[16]이러한 관찰에 기초하여, 이 저자들은 산소화된 피토그스의 함수는 NO 이산화질소 활동을 통해 NO의 수준을 변조하고 NO의 수준에 의해 변조되는 매우 다양한 셀 기능을 간접적으로 규제하는 것이라고 제안했다.질산염을 생성하기 위해 NO와 반응하는 산소화된 1등급 피토글로빈은 식물에서 NO를 파내는 주요 메커니즘을 나타낸다.질산염 환원효소, 질산염을 NO로 감소시키고, 피토글로빈에 의한 NO를 청소하는 사이클을 피토글로빈-NO 사이클로 정의했다.[17]저산소증 시 리독스 및 에너지 상태를 유지하며 에탄올과 젖산의 생산량 감소로 이어진다.[18]
피토그브스0, 1, 2, 3은 다양한 (배양 및 식물성) 식물 기관에서 매우 낮은 농도로 합성된다.[19][20][21]그러나 홍수와[22] 빛 제한과 같은 특정한 스트레스 조건에 따른 식물에서는 피토그스의 농도가 증가한다.[23]따라서, 일부 피토그는 식물의 응력에 반응하는 단백질로 여겨져 왔다.
진화
이 글의 일부분(Becana phylogeny와 관련된 것들은 Lbs와 Pytogb1의 SymPhytogbs에 대한 공통 기원을 거부한다)은 업데이트가 필요하다.(2020년 11월) |
아레돈도-피터, 비노그라도프, 협력자들에 의한 연구는 피토그스의 진화 동안에 일어날 수 있었던 주요 사건들을 해명하였다.[24][25][26]피토그bs는 진핵생물(euvacteria domain)의 기원 이전의 공통의 조상으로부터 진화하였지만, 식물에서는 피토그브에 대해 두 가지 진화 선, 즉 (3/3) 피토그b0, 1, 2와 SymPhytogb 및 Lb 혈통과 (2/2-) 피토그b3 혈통이 확인되었다.분명히 피토그브스0은 피토그브스1과 2의 조상이었고, 심피토그브스와 Lbs는 공생 질소 고정에 적응하기 위해 피토그브1의 조상으로부터 진화했다.Comparative analysis of moss Phytogb0 with rice Phytogb1 and soybean Lb structure revealed that the major evolutionary changes that probably occurred during the evolution of Phytogbs0, 1 and 2, SymPhytogbs and Lbs were (i) a hexa-coordinate to penta-coordinate transition at the heme group, (ii) a length decrease at the CD-loop and N- and C-termini그리고 단백질의 혼합을 입상 구조로 만든다.또한, Pytogbs3의 글로빈 영역의 구조는 박테리아 잘린 글로빈 구조와 매우 유사하여 Pytogbs3가 진화하는 동안 2/2 접이 보존되었음을 나타낸다.그러나 육지식물 피토그브스3의 글로빈 영역에는 아마도 조상으로부터 피토그브스3 육지식물에서 유래한 엑스트라 아미노산 시퀀스가 측면에 있다.피토그브스3 구조에서 엑스트라 아미노산 시퀀스의 기능적, 진화적 중요성은 여전히 알려져 있지 않다.
기술 응용 프로그램
피토그는 잠재적으로 많은 생명공학 용도에 유용하다.예를 들어 쌀 Phytogbs1 발견 직후 의도된 적용은 이러한 단백질을 전자기기에서 O센서로 사용하는2 것이었다.또한, 피토그브스의 과표현은 초기 홍수 신호 에틸렌을 식물의 낮은 산소 반응에 결합시켜 농작물 범람과 같은 특정 스트레스 조건에 대한 농작물 내성을 높이기 위한 전략으로 제안되었다.[27][28][29][30]최근, 피토그브는 혈액 대체물을 개발하고 야채버거에 첨가제를 첨가한 후보지로 여겨졌다.
메모들
참조
- ^ 쿠보 H, 우버 하모프로틴 아우스 덴 우르젤나놀첸 폰 레기미노센, 액타 피토힘.(도쿄), 11 (1939년) 195-200.
- ^ 배니슈테인 B. K. 하루티우냔 E. H. 쿠라노바 I. P. 보리스노프 V.V.N.I.소스페노프, 파블로프스키 A. G. 그레벤코 A.I, Konareva N. V, 5 A 분해능에서 루핀 루트 결절에서 레게모글로빈 구조, 네이처, 254 (1975) 163-164
- ^ a b c d e f Becana, Manuel; Yruela, Inmaculada; Sarath, Gautam; Catalán, Pilar; Hargrove, Mark S. (September 2020). "Plant hemoglobins: a journey from unicellular green algae to vascular plants". New Phytologist. 227 (6): 1618–1635. doi:10.1111/nph.16444. PMID 31960995.
- ^ a b c d Hill R., Hargrove M. S., Arredondo-Peter R (2016). "Phytoglobin: a novel nomenclature for plant globins accepted by the globin community at the 2014 XVIII conference on Oxygen-Binding and Sensing Proteins". F1000Research. 5: 212. doi:10.12688/f1000research.8133.1. PMC 4792203. PMID 26998237.
{{cite journal}}: CS1 maint: 작성자 매개변수 사용(링크) - ^ 마르티네즈-오캄포 F, 바스케스-리몬 C, 아르레돈도-피터 R, 검출, PCR-증화 및 특성화, 글로벌 J. 생화학, 3 (2012) 12 이끼코미트렐라 패틴스로부터 4 intron 함유 비생체 헤모글로빈 유전자
- ^ Hardison R. C, 헤모글로빈에 대한 간략한 역사: 식물, 동물, 양성자, 박테리아, Proc.나틀. 아카드.미국 과학, 93 (1996) 5675-5679
- ^ Go M, 네이처, 291 (1981) 90-92의 헤모글로빈 내 단백질 구조 단위와 DNA 외전 부위의 상관 관계.
- ^ Hyldig-Nielsen J. J. Jensen E. Paludan K, Wiborg O, Garret R, Joergersen P. L., Marker K. A. 콩, 핵산 레그헤모글로빈 유전자의 1차 구조, 핵산 레그 헤모글로빈 유전자의 1차 구조, 10 (1982) 689-701.
- ^ Hargrove M, Brucker E. A., Stec B, Sarath G, Arredondo-Peter R, Klucas R. V, Olson J. S. PhilipsJr.G. N, 비생체 헤모글로빈의 결정구조, Structure, 8(2000) 1005-1014.
- ^ 리드 B. J.Hough M. A., 아라비도피스 탈리아나의 3등급 비생균 식물 헤모글로빈의 구조는 새로운 N단자 나선 연장선인 액타 결정체르, D70 (2014) 1411-1418을 보여준다.
- ^ Arredondo-Peter R, Hargrove M. S, Moran J. F, Sarath G, Klucas R. V, 식물 헤모글로빈, 식물 물리, 118 (1998) 1121-1126.
- ^ Arredondo-Peter R., Hargrove M. S., Sarath G., Moran J. F., Lohrman J., Olson J. S.,Klucas R. V., Rice hemoglobins: gene cloning, analysis and oxygen-binding kinetics of a recombinant protein synthesized in Escherichia coli., Plant Physiol., 115 (1997) 1259-1266
- ^ 비텐베르크 J. B. 버거센 F.J. 애플비 C. A.터너 G. L., 촉진 산소 확산: 콩 뿌리 결절에서 격리된 박테로이드에 의한 질소 고정에서 레그헤모글로빈의 역할, J. 비올.화학, 249 (1974년) 4057-4066
- ^ 애플비 C.A, 레게모글로빈과 리조비움 호흡, 안누.식물 물리학자, 35 (1984) 443-478
- ^ Smagghe B. J. T. Hargrove M. S. Hargrove M. S. Hargrove M. S. NO dioxygenase 활성은 시험관 내 어디에나 있지만, 체내 감소에 의해 제한된다. PLoS One, 3(2008) doi: 10.1371/journal.pone.0002039.
- ^ 힐 R. D. 비생체성 헤모글로빈-질소산화물 청소 외에 무슨 일이 일어나고 있는가?AoB 공장, Pls004(2012) doi: 10.1093/aobpla/pls1004
- ^ Igamberdiev, A.U.; Baron, K.; Manac'h-Little, N.; Stoimenova, M.; Hill, R.D. (2005). "The Haemoglobin/Nitric Oxide Cycle: Involvement in Flooding Stress and Effects on Hormone Signalling". Annals of Botany. 96 (4): 557–564. doi:10.1093/aob/mci210. ISSN 0305-7364. PMC 4247025. PMID 16027133.
- ^ Igamberdiev, A.U.; Hill, R.D. (2004). "Nitrate, NO and haemoglobin in plant adaptation to hypoxia: an alternative to classic fermentation pathways". Journal of Experimental Botany. 55 (408): 2473–2482. doi:10.1093/jxb/erh272. ISSN 0022-0957. PMID 15448180.
- ^ 가루로초-빌레가스 V, 아르레돈도-피터 R, 이끼의 분자 복제와 특성화(Ceratodon paipureus) 비생체 헤모글로빈(Mol. Biol) 식물의 초기 진화에 대한 통찰력을 제공한다.Evol, 25 (2008) 1482-1487.
- ^ Ross E. J. H, Shearman L, Mathiesen M, Jou J, Arredondo-Peter R, Sarath G, Klucas R. V, 비생체 헤모글로빈은 발아 중과 차별화 세포 유형인 프로토플라즈마, 218 (2001) 125-133에서 합성된다.
- ^ Watts R. A, Hunt P. W, Hvited A. N, Hargrove M. S, Pocket W. J, Dennis E. S, 미생물의 잘린 헤모글로빈에 이르는 식물에서 나오는 헤모글로빈, Proc.나틀. 아카드.미국 과학, 98 (2001) 10119-10124.
- ^ Taylor E. R, Nie X. Z, MacGregor A. W, Hill R. D, 곡물 헤모글로빈 유전자는 혐기성 조건인 Plant Mol에서 씨앗과 뿌리 조직으로 표현된다.비올, 24 (1994년) 853-862
- ^ 리라루안 V, 사라스 G, 클루카스 R. V, 아르레돈도-피터 R, 쌀 속의 헤모글로빈 합성(오리자 사티바 바)잭슨) 정상 및 스트레스 조건에서 자라는 식물, 플랜트 Sci, 161 (2001) 279-287.
- ^ Vazquez-Limon C, Hoogewijs D, Vinogradov S. N, Arredondo-Peter R, 육지 식물 헤모글로빈의 진화, 식물 과학, 191-192 (2012) 71-81.
- ^ 비노그라도프 S. N, 페르난데스 I, 후게위즈 D, 아르레돈도-피터 R, 고골플라스티다 게놈의 3/3 및 2/2 헤모글로빈과 박테리아 및 기타 진카리오테 헤모글로빈, 몰. 플랜트, 4(2011) 42-58.
- ^ 비노그라도프 S. N., 후게위즈 D., 아르레돈도-피터 R., 식물 헤모글로빈의 기원과 기원은 무엇인가?, 코뮌.통합.비올, 4 (2011) 443-445.
- ^ Andrzejczak, OA; Havelund, JF; Wang, WQ; Kovalchuk, S; Hagensen, CE; Hasler-Sheetal, H; Jensen, ON; Rogowska-Wrzesinska, A; Møller, IM; Hebelstrup, KH (24 February 2020). "The Hypoxic Proteome and Metabolome of Barley (Hordeum vulgare L.) with and without Phytoglobin Priming". International Journal of Molecular Sciences. 21 (4): 1546. doi:10.3390/ijms21041546. PMC 7073221. PMID 32102473.
- ^ Mira, Mohamed M.; Hill, Robert D.; Stasolla, Claudio (November 2016). "Phytoglobins Improve Hypoxic Root Growth by Alleviating Apical Meristem Cell Death". Plant Physiology. 172 (3): 2044–2056. doi:10.1104/pp.16.01150. PMC 5100795. PMID 27702845.
- ^ Hartman, Johannes Gerardus Wilhelmus (2020-01-22). The early flooding signal ethylene acclimates plants to survive low-oxygen stress. Universiteit Utrecht. hdl:1874/387763. ISBN 978-90-393-7231-9.
- ^ Hartman, S; Liu, Z; van Veen, H; Vicente, J; Reinen, E; Martopawiro, S; Zhang, H; van Dongen, N; Bosman, F; Bassel, GW; Visser, EJW; Bailey-Serres, J; Theodoulou, FL; Hebelstrup, KH; Gibbs, DJ; Holdsworth, MJ; Sasidharan, R; Voesenek, LACJ (5 September 2019). "Ethylene-mediated nitric oxide depletion pre-adapts plants to hypoxia stress". Nature Communications. 10 (1): 4020. Bibcode:2019NatCo..10.4020H. doi:10.1038/s41467-019-12045-4. PMC 6728379. PMID 31488841.