호수대사
Lake metabolism.jpg)
호수의 신진대사는 탄소 고정(총 1차 생산)과 생물학적 탄소 산화(생태계 호흡) 사이의 호수의 균형을 나타낸다.[1] 전혈대사에는 박테리아에서 물고기까지 호수 내 모든 유기체의 탄소 고정과 산화가 포함되며, 일반적으로 하루 종일 용해된 산소나 이산화탄소의 변화를 측정하여 추정한다.[2]
1차 총생산을 초과하는 생태계 호흡은 호수가 하천이나 지하수 유입이나 쓰레기와 같은 주변 유역으로부터 유기물을 공급받음을 나타낸다. 호수 신진대사는 종종 호수로의 이산화탄소 배출량을 조절하지만, 주변 유역에서 나오는 무기탄소의 투입이 호수 내 이산화탄소에도 영향을 미치기 때문에 모든 이산화탄소 역학을 설명하지는 않는다.[3][4]
개념
호수 신진대사의 추정치는 일반적으로 용존산소나 이산화탄소의 측정, 또는 유기탄소의 생산과 소비량을 추정하기 위한 탄소나 산소 추적기의 측정에 의존한다. 산소는 광합성을 통해 생산되고 이산화탄소는 소비되며 산소는 호흡으로 배출된다. 여기서 유기물은 포도당으로 상징되지만, 이러한 반응을 통해 생산되고 재생되는 화학 종은 매우 다양하다.

광합성: + 6 → g 6 + 6
호흡:
![{\displaystyle C_{6}H_{12}O_{6}+6O_{2}{\xrightarrow[{}]{}}6CO_{2}+6H_{2}O}](https://wikimedia.org/api/rest_v1/media/math/render/svg/619bc6294a1bbc343eac76ab799d767420c04e83)
광합성과 산소 생성은 빛이 존재하는 곳에서만 일어나는 반면, 호흡을 통한 산소의 소비는 빛의 존재와 부재 모두에서 일어난다. 호수 신진대사 용어에는 다음이 포함된다.
- GPP - 1차 총 생산량(예: 총 광합성)
- R - 총 호흡 h+ )
- - 이질성 호흡
- a - 자가영양호흡
- NEP - 순 생태계 생산 = GPP - R
- NPP - 순 1차 생산 = GPP -


측정 기법
호수의 신진대사를 추정하기 위해서는 호수의 유기체들에 의한 유기탄소의 생산과 소비에 영향을 미치는 근사적인 과정이 필요하다. 태양빛은 광합성과 새로운 탄소의 생산에만 이용되기 때문에 지구의 대부분의 호수에서 매일 주기적인 변화가 일어난다. 연구자들은 이 diel 패턴을 이용하여 탄소 자체의 변화율이나 이산화탄소나 산소와 같은 용해된 기체의 변화율을 일일 규모로 측정할 수 있다. 일일 신진대사 추정치가 가장 일반적이지만, 통기류 탄소 예산을 추정함으로써 계절적 또는 연간 비율과 같은 긴 기간에 걸쳐 통기류 신진대사를 통합할 수 있다. 다음 절에서는 다양한 시간적 및 공간적 척도에 걸쳐 호수 대사를 추정하는 가장 일반적인 방법을 강조하고 이러한 각 방법의 몇 가지 가정을 살펴본다.

자유수법
"자유수" 방법으로도 알려진 호수 내 용존 가스의 다이엘 변화 측정은 물에 용해된 산소와 이산화탄소를 측정하는 데 사용되는 자율 센서의 광범위한 채택 이후 호수 대사를 추정하는 가장 일반적인 방법이 되었다.[6][7][8] 이 자유수법은 호수대사에 대한 많은 일일 추정치를 비교적 저렴하게 수집할 수 있고 폭풍우 사건처럼 관찰하기 어려운 기간 동안 대사 체계에 대한 통찰력을 줄 수 있기 때문에 특히 인기가 있다. 호수 내 용존산소와 이산화탄소의 측정된 변화는 용존가스의 생화학 변화를 고려한 후 박테리아에서 물고기까지의 모든 유기체 대사의 합을 나타낸다. 용해된 기체의 생물학적 변화로는 대기와 호수 표면 사이의 용해된 기체의 교환, 다른 농도의 물의 수직 또는 수평적 유입(예: 호수의 열전선 아래의 저산소 물) 또는 유입되는 하천이나 호수 배출구에서 용해된 기체의 수출입이 포함된다. 용존가스의 아바이오틱스 변화는 호수의 대사율이 낮거나(예: 과두영양호수, 흐린 날), 또는 아바이오틱 인자가 생물학적 인자를 초과하는 큰 사건(예: 저산소화수의 혼합과 유입을 유발하는 바람 사건)이 있을 경우 용존가스의 변화를 지배할 수 있다. 용해된 가스의 생물학적 신호는 태양이 빛나고 광합성이 일어나 용해된 산소의 생산과 이산화탄소의 소비로 이어질 때 가장 뚜렷하게 나타난다. 태양 에너지를 화학 에너지로 변환하는 것을 총 1차 생산(GPP)이라고 하고, 생물학적 탄소 산화를 통한 에너지 소산을 생태계 호흡(ER)이라고 한다. 용존산소나 이산화탄소의 고주파(예: 10분 간격) 측정은 고주파 데이터를 호수대사 모델에 적합시킴으로써 GPP, ER 및 두 용어인 순생태계 생산(NEP)의 차이에 대한 추정치로 환산할 수 있다. 용존산소를 측정하는 상부 혼합층에 위치한 단일 센서에서 호수 대사를 추정하기 위한 지배 방정식은 다음과 같다.
DO/t = GPP-ER+F
여기서 F는 호수와 대기 사이의 기체 흐름이다. 호수에 대해 그러한 생물학적 유동성이 유의하다고 간주되는 경우(예: 혼합 사건, 하천 가스 유입) 추가적인 항을 추가할 수 있다. 대기 가스 교환(F)은 거의 직접 측정되지 않으며, 일반적으로 풍력 및 대류 혼합에서 발생하는 호수 표면 난류를 추정하여 모델링한다. 대부분의 경우 F는 풍속과 대기압의 측정으로 추정되며 F를 추정하기 위한 다른 모델은 연구 호수에 따라 호수 대사율의 추정치가 크게 다를 수 있다.[9] 1차 총생산은 낮거나 빛이 없어 야간 중 0으로 가정하여 용존산소의 생화학 변화를 고려한 용존산소(또는 이산화탄소)의 야간 변화로 ER을 추정할 수 있다. 일차 총 생산량은 ER이 주야간 동일하다고 가정하고 낮 동안 용존 산소 변화를 고려한다고 가정할 수 있지만, 이러한 가정은 모든 호수에서 유효하지 않을 수 있다.[10]
높은 신호 대 잡음 비를 추출하는 것은 자유수법으로부터 호수 대사의 좋은 추정치를 얻는 데 핵심이며, 정확한 추정치를 확보하기 위해 수집 데이터와 데이터 분석 중에 연구자가 선택해야 하는 선택사항이 있다. 용해된 가스 채집 위치(일반적으로 표면 혼합층), 수직 및 수평으로 센서 수,[11][12][13] 데이터 채집 빈도와 지속시간, 모델링 방법 등을 고려할 필요가 있다.[14]
자유수질대사 모델링 기법
자유수량 측정 기법은 고주파 용해 가스 측정에서 호수 대사 지표를 추정하기 위한 수학적 모델이 필요하다. 이러한 모델은 단순한 대수적 모델에서 보다 진보된 통계 기법을 사용한 깊이 통합 모델링에 이르기까지 복잡성이 다양하다. GPP, R 및 NEP 또는 이러한 신진대사 용어와 관련된 매개변수를 추정하기 위해 몇 가지 통계적 기법이 사용되어 왔다.
빛과 어둠의 병법
빛과 어둠의 병 방법은 신진대사의 비율을 추정하기 위해 자유수법과 같은 개념을 사용한다 - GPP는 빛이 있는 것과 없는 것 모두에서 ER이 발생하는 동안 태양 에너지를 가지고 낮에만 발생한다.[15] 이 방법은 호수의 물을 두 개의 분리된 병으로 배양하는데 하나는 맑고 자연광 또는 인공광체계에 노출되는 병이고 다른 하나는 호일, 페인트 또는 다른 방법으로 병을 포장하여 빛으로부터 막는다. 탄소 고정 또는 용존 가스의 변화는 특정 호수 깊이 또는 통합된 호수 물기둥에 대한 신진대사의 비율을 추정하기 위해 일정 기간(예: 몇 시간에서 하루)에 걸쳐 측정된다. 탄소 고정은 빛과 어두운 병에 방사성 탄소 동위원소 C를 주입하고 병을 샘플링하여 측정한다. - 샘플은 여과지에 걸러지고, 녹조(및 박테리아) 세포에 통합된 C의 양은 섬광 카운터에서 샘플을 측정하여 추정한다. 빛 병 C와 어두운 병 C의 차이는 일차 생산성의 비율로 볼 수 있다. 그러나2, CO의 비포토합성 흡수로 인해 어두운 병을 C 방법으로 사용해야 하는지 아니면 알조제 DCMU로 처리된 가벼운 병과 병만 사용해야 하는지에 대한 논쟁이 있다. 이산화탄소나 산소와 같은 용해된 가스의 변화율은 생산성과 호흡의 비율을 추정하기 위해 빛과 어둠의 병 모두를 필요로 한다.
전리크 탄소 예산 방법
아마도 호수대사의 측정지표를 추정하는 가장 노동집약적인 방법은 한 계절이나 한 해에 걸쳐 호수에 대한 유기 탄소나 무기탄소의 모든 투입과 출력을 측정하는 것으로, 전유량 탄소 예산으로도 알려져 있다. 호수로 들어오고 나가는 탄소의 모든 입력과 출력을 측정하는 것은 순생태계 생산량(NEP)을 추정하는 데 사용될 수 있다.[16][17] NEP는 1차 총생산과 호흡(NEP = GPP - R)의 차이이므로 무기탄소를 유기탄소로 순 생물학적 변환(그리고 그 반대)하는 것으로 볼 수 있으며, 따라서 무기탄소 또는 유기탄소의 전체적 질량 균형을 통해 결정할 수 있다.[16] 무기물(IC) 또는 유기 탄소(OC)를 통해 평가된 NEP는 다음과 같이 추정할 수 있다.


OC의 하천 운송을 이용하여 대기 중에 하천 교통과 탄소 가스가 호수 표면 사이에(예를 들어 이산화 탄소, 메탄)교환까지 어디서 E는 수출과 IC, S는 호수 퇴적물과 IC에 OC와 물을 열에 대한 물 기둥, 그리고 강박과 IC의 하천, 주변 습지에서 내가 입력이 보관, 및 공중 경로(예를 들어 atmo.sp그녀의 퇴적, 쓰레기 투하 유역으로부터 OC를 하류로 수출하거나 물기둥과 퇴적물에 축적하는oc 것(Ioc > Eoc + S)보다 더 많이 받는 호수는 호수 내에서 OC를 IC로 순전환한 것이 있었음을 나타내며, 따라서 순 헤테로이드성(네페)이다. 마찬가지로 유역(Sicic + E > I)으로부터ic 받은 IC보다 더 많은 IC를 축적·수출하는 호수 역시 호수 내에서 OC의 IC로의 순전환(순전환)을 나타내며, 따라서 순 헤테로이드성(순 헤테로이드성)이다.
벤트닉 대사법
자유수법에는 약간의 벤트릭 대사신호가 포함되어 있을 가능성이 있지만, 전리크 신진대사에 대한 벤트닉 기여도를 격리하는 것은 벤트릭 특유의 방법이 필요하다. 위에서 설명한 빛과 암병 방법과 유사하게 호수 침전물 코어를 수집할 수 있으며 용존 산소 또는 탄소 고정의 변화를 사용하여 일차 생산성과 호흡의 비율을 추정할 수 있다. 비교적 새로운 방법은 투명한 돔으로 침전물-물 접점을 분리하고 용존 산소 in-situ의 변화를 측정하는 것으로, 자유수법과 연두암 병법의 혼합물이다.[18] 이러한 현장 벤트닉 챔버 방법은 벤트닉 신진대사를 비교적 쉽게 다일 추정할 수 있도록 하는데, 이는 연구자가 다양한 날씨 패턴과 호수 특성에 따라 벤트닉 대사가 어떻게 변화하는지 판단하는 데 도움이 된다.
가정
호수[19] 내에서 수직 및 수평으로 상당한 대사 변동성이 있을 수 있기 때문에 호수 전체에 대한 외삽 현장 또는 깊이별 측정은 문제가 될 수 있다(변동성 섹션 참조). 예를 들어, 많은 호수 신진대사 연구는 신진대사에 대한 하나의 경구적 추정치만 가지고 있지만, 이것은 혼합층 깊이 대 가벼운 멸종 깊이 비율에 따라 NEP와 같은 호수에 대한 대사 특성을 과대평가할 수 있다.[20][21] 장기간의 일일 신진대사 추정치를 평균화하면 이러한 단일 부위 외삽 문제를 극복하는 데 도움이 될 수 있지만,[19] 지나치게 외삽된 측정치가 아닌 신진대사 추정치의 영향을 신중하게 고려해야 한다.
구성 요소와의 관계
유기체 대사율, 즉 유기체가 에너지를 동화, 변형, 소모하는 속도는 빛, 영양소, 온도, 유기 물질 등 몇 가지 주요 성분에 의해 영향을 받는다. 이러한 성분들이 유기체의 신진대사에 미치는 영향은 궁극적으로 전유량에서 신진대사를 지배하며, 호수가 탄소배출원인지 아니면 싱크대인지를 지시할 수 있다. 다음 절에서는 이러한 핵심 성분과 유기체 및 생태계 수준의 신진대사 사이의 관계를 설명한다. 여기서 설명하는 유기체와 성분 간의 관계는 잘 확립되어 있지만, 유기체에서 호수 생태계에 이르는 대사율에 대한 성분들의 상호작용 효과는 호수를 가로지르는 혹은 호수의 신진대사의 변화를 시간적으로 예측하기 어렵게 만든다. 이러한 복잡한 상호작용 효과는 공간적 및 시간적 가변성 섹션에서 논의될 것이다.
온도

온도는 생화학적 반응률과 생물학적 활동을 통제하는 강력한 요인이다. 최적의 온도는 수생 유기체에 따라 달라지는데, 이는 일부 유기체가 더 추운 적응을 하는 반면 다른 유기체는 더 따뜻한 서식지를 선호하기 때문이다. 초절정 남극호수(예: 돈 후안 연못)나 온천(예: 플라이 간헐천)에서 극도의 내열성을 보이는 경우는 드물지만, 지구의 대부분의 호수 생물체는 섭씨 0도에서 40도의 온도에 서식한다. 대사율은 일반적으로 온도에 따라 기하급수적으로 확장되지만, 1차 생산성과 호흡에 대한 활성화 에너지는 종종 달라지는데, 광합성은 유산소 호흡보다 활성화 에너지가 낮다. 활성화 에너지의 이러한 차이는 기후가 온난화됨에 따라 호수 생태계 내의 순대사 균형에 영향을 미칠 수 있다. 예를 들어 샤펜베르거 외(2019년)[23]는 기후변화에 따른 수온 상승이 활성화 에너지의 차이로 인해 호수가 순 자폐성에서 이기성으로 바뀔 수 있다는 것을 보여주지만 호수가 전환되는 온도는 이용 가능한 영양소의 양에 따라 달라진다.
영양소
유기 세포에 동화되는 데 이용 가능한 물질의 양은 세포에서 호수 생태계에 이르는 신진대사의 비율을 조절한다. 호수에서 인과 질소는 일차 생산과 생태계 호흡의 가장 흔한 제한 영양소다. 인의 농도와 호수의 영적화 사이의 긍정적인 관계에 대한 기초적인 연구는 세탁 세제에서 인의 양을 제한하는 법률을 만들었는데, 그 중에서도 다른 규정들 중에서도.[24][25] 인은 호수 생태계 생산성과 과잉인 인의 예측 변수로 자주 이용되지만, 신진대사는 인과 질소 또는 질소만으로 공동 제한된다는 연구결과가 많다.[26] 생태학적 스토이치측정법으로 불리는 인, 질소, 그리고 다른 영양소들 사이의 균형은 생명-역사적 특성으로 매개되는 이러한 필수 영양소의 세포 요구 사항을 통해 유기체의 성장과 전유동 신진대사의 속도를 좌우할 수 있다. 예를 들어 급성장하는 클래도케란들은 주로 빠른 성장을 위해 사용되는 세포에서 인이 풍부한 RNA의 양이 많기 때문에 요각류보다 질소 대 인 비율(N:P)이 훨씬 낮다. 클라도케란 신체 단층계에 비해 N:P 비율이 높은 호수에 거주하는 클라도케란인은 성장과 신진대사가 제한돼 전혈구 신진대사에 영향을 미친다. 게다가, 음식 거미줄 조작으로 인한 계단식 효과는 영양소 계량법으로의 변화로부터 생산성의 변화를 일으킬 수 있다. 예를 들어, 피시보어를 첨가하면 빠르게 증가하는 낮은 N:P 클래도케란에 대한 포식 압력을 줄일 수 있으며, 이는 개체수를 빠르게 증가시키고, 인을 세포에 보존하며, 호수가 인으로 제한되어 결과적으로 전체 레이크 1차 생산성이 감소할 수 있다.
빛
태양 에너지는 이산화탄소와 물을 광합성이라고 알려진 유기 물질로 전환하는데 필요하다. 온도나 영양소의 경우와 마찬가지로, 어떤 해조류는 어두운 환경에 더 잘 적응하고 다른 해조류는 더 가벼운 조건에서 경쟁할 수 있기 때문에, 조류의 신진대사 반응 속도가 빛의 증가에 따라 다르고 또한 성장을 위한 최적의 빛 조건도 다르다. 빛은 또한 증가하는 빛에 대한 종 특유의 조류 생산성 반응에 영향을 주기 위해 영양소와 상호작용할 수 있다.[27] 유기체 수준에서 이러한 다른 반응은 생태계 수준에서 신진대사에 영향을 미치기 위해 전파된다.[28][29] 영양소가 1차 생산성의 제한 자원이 될 것으로 기대되는 저영양 호수에서도 빛은 여전히 제한 자원이 될 수 있으며, 이는 어류 생산성과 같은 영양성 높은 수준에 연쇄적으로 부정적인 영향을 미친다.[30] 다른 호수 지역과 호수 내에서 시간 경과에 따른 빛의 변동성은 공간적으로나 일시적으로 생산성에 패치를 초래한다.
일차 생산성을 조절하는 것 외에도, 햇빛은 유기 물질을 부분적으로 산화시켜 호흡 속도에 영향을 줄 수 있는데, 이것은 박테리아가 분해되어 이산화탄소로 전환되기 쉽게 할 수 있다. 이 부분적인 광산화 작용은 본질적으로 광물화에 이용 가능한 유기 물질의 양을 증가시킨다.[31] 일부 호수에서는 완전광산화 또는 부분광산화 등이 유기물질에서 무기물질로의 변환의 대부분을 차지할 수 있지만, 박테리아 호흡에 대한 비율은 호수마다 크게 다르다.
유기탄소
호수의 1차 소비자와 2차 소비자는 유기체 기능을 유지하기 위해 유기 물질(식물이나 동물로부터)을 필요로 한다. 나무 잎, 용해된 유기물, 해조류를 포함한 유기물은 이러한 소비자들에게 필수적인 자원을 제공하고 그 과정에서 유기물을 세포 성장과 유기체 유지로 전환하는 과정에서 호수 생태계의 호흡수를 증가시킨다. 유기 물질의 일부 원천은 다른 구성 요소의 가용성에 영향을 미칠 수 있다. 예를 들어, 용해된 유기물은 종종 호수 물을 어둡게 하여 호수에서 이용 가능한 빛의 양을 감소시켜 1차 생산량을 감소시킨다. 그러나 호수에 대한 유기물 하중의 증가는 유기물과 관련된 영양소를 증가시켜 1차 생산과 호흡을 자극할 수 있다. 용해된 유기물 하중의 증가는 빛의 제한 증가와 영양소의 제한으로부터의 방출 사이에서 절충을 일으킬 수 있다. 이러한 절충은 유기물질과 얼마나 많은 영양소가 연관되어 있는지 그리고 용해된 유기물질이 물기둥의 빛을 얼마나 빨리 차단하는지에 기초하여 호수 1차 생산과 용해된 유기물 하중 사이에 비선형 관계를 만들 수 있다.[32][33][34][35] 용해된 유기물 농도가 높아짐에 따라 낮은 용해 유기물 농도에서는 관련 영양소의 증가가 GPP를 강화시키기 때문이다.[32][33][34] 그러나 용해된 유기물이 계속 증가함에 따라, 호수 물이 어두워짐에 따른 빛의 감소는 빛이 일차 생산성의 제한 자원이 되면서 GPP를 억제한다.[32][33][34] DOC 부하 증가에 대응한 최대 GPP의 크기와 위치의 차이는 호수로 들어오는 영양소에 대한 DOC의 비율과 호수 빛 기후에 대한 DOC의 영향에 기초하여 발생한다고 가정한다.[34][36] 호수 물을 어둡게 하는 것은 보통 더 따뜻한 물이 호수 꼭대기에 남아 있고, 더 차가운 물이 바닥에 있다는 것을 의미하기 때문에 호수 내부의 열전도 바꿀 수 있다. 이러한 열 에너지 분포의 변화는 펠로직 및 벤트직 생산성 비율(위 온도 참조)에 영향을 미칠 수 있으며, 영양소의 수직 분포에 영향을 미치면서 물기둥 안정성을 변화시킬 수 있으며, 따라서 대사율의 수직 분포에 영향을 미칠 수 있다.

기타 성분
다른2 호수 성분들은 CO 농도, pH, 염도, 실리카를 포함한 호수 대사율에 영향을 미칠 수 있다. CO는2 1차 생산성을[38] 위한 제한적(또는 다른 영양소와 함께 공동 제한) 자원이 될 수 있으며, 보다 강렬한 식물성 플랑크톤 꽃을 촉진할 수 있다.[39] 번데기와 같은 일부 녹조 종은 탄소를 농축하는 메커니즘이나 광합성을 위한 무기탄소의 원천으로 중탄산을 사용하는 능력이 없을 수 있으므로, CO의2 상승은 그들의 광합성 속도를 증가시킬 수 있다. 녹조 중에 용해된 CO를2 증가시키면 CO가 급격히 증가하여 CO가2 고갈되고 pH가 증가하기 때문에 CO가2 성장을 위한 제한적인 자원이 되지 않는다. 일차 생산성의 급상승으로 인한 단기적 규모의 pH 변화(예: 서브데일리)는 박테리아 성장과 호흡의 단기적 감소를 야기할 수 있지만, 더 긴 시간에는 박테리아 공동체가 pH 상승에 적응할 수 있다.[40][41]
또한 염도는 개인의 대사율과 공동체 구성에 대한 염분 영향을 통해 호수의 대사율에 변화를 일으킬 수 있다.[42][43][44] 호수 대사율은 홍조율이나 가뭄과 같은 생태계 신진대사의 다른 동인들과의 염분 상호작용에 의해 염분과 긍정적이거나 부정적으로 상관될 수 있다.[45] 예를 들어, Moreira-Turcq(2000년)[46]는 증발보다 과도한 강수량이 해안 석호의 염분 감소, 영양소 부하 증가, 펠라직 1차 생산성 증가를 유발한다는 것을 발견했다. 일차 생산성과 염분 사이의 긍정적인 관계는 유입량 증가에 따른 영양소 가용성 변화를 나타내는 지표일 수 있다. 그러나 도로 염분으로부터의[47] 염분 증가는 일부 호수 유기체에 독성을 유발할 수 있으며,[48] 염분 증가는 호수의 혼합을 제한하여 호수 물기둥 전체의 신진대사율 분포를 변화시킬 수 있다.
공간 및 시간적 변동성
호수와 저수지의 대사율은 빛과 영양소의 가용성, 온도, 물기둥 혼합 방식과 같은 많은 환경적 요인에 의해 제어된다. 따라서 이러한 요소들의 공간적, 시간적 변화는 대사율의 공간적, 시간적 변동성을 야기하며, 그러한 요소들은 각기 다른 공간적, 시간적 척도로 신진대사에 영향을 미친다.
호수내 공간변동향
다른 호수 지역(즉, 연막, 림프, 벤트닉)에서 전체 호수 대사에 대한 가변적 기여는 대부분 녹조 및 박테리아 바이오매스의 패치성 및 빛과 영양소의 가용성에 의존한다. 이들 각 구역에서 신진대사에 기여하는 유기체의 측면에서 림프성 대사는 식물성 플랑크톤, 동물성 플랑크톤, 박테리아성 대사에 의해 지배되고 있으며, 인식체 및 생선의 기여도는 낮다. 벤트의 신진대사는 마크로피테스, 매크로와 미세조류, 무척추동물, 박테리아로부터 많은 기여를 받을 수 있다. 벤트의 신진대사는 보통 얕은 연근대 또는 맑은 물이 얕은 얕은 호수에서 가장 높으며, 빛이 호수 바닥에 도달하여 1차 생성을 자극한다. 어둡거나 탁한 깊은 호수에서는 일차생산을 얕은 물로 제한할 수 있으며, 음산성 깊은 구역의 형성으로 인해 깊은 수역에서는 유산소호흡이 감소하거나 존재하지 않을 수 있다.
호수 내 대사율의 공간적 이질성의 정도는 호수 형태측정법, 유역 특성(예: 유역 및 하천으로부터의 유입에 따른 토지 이용의 차이) 및 유역학적 프로세스에 따라 달라진다. 예를 들어, 강한 수직 및 횡방향 혼합과 같이 보다 강한 유체역학 과정을 가진 호수는 고도로 변형된 호수보다 신진대사율과 관련하여 횡방향과 수직방향으로 균일하다. 반면 연돌 부위가 발달한 호수는 보다 원형적인 형태와 얕은 연돌 부위의 비율이 낮은 호수보다 횡방향으로 대사 이질성이 더 크다.
물기둥 전체에서 발생하는 빛 감쇠는 열 및 화학 성층화, 바람 또는 대류성 난류와 결합하여 물기둥 내 영양소와 유기체의 수직적 분포에 기여한다. 층화된 호수에서는 유기물과 영양소가 더 깊은 층에 집중되는 경향이 있는 반면, 얕은 층에서는 빛이 더 많이 이용 가능하다. 1차 생산의 수직적 분포는 빛과 영양소 가용성 사이의 균형에 반응하는 반면, 호흡은 빛과 영양소로부터 더 독립적으로, 그리고 심도와 함께 더 균등하게 일어난다.[49] 이로 인해 호수 표면층에서는 GPP(총 1차 생산)와 생태계 호흡(ER)이 강하게 결합되지만 더 깊은 곳에서는 결합이 약해지는 경우가 많다. 즉 ER 속도는 1차 생산에 크게 의존하는 반면, 보다 깊은 층에서는 육지 공급원의 유기 물질과 조류 입자의 침전 및 샬로우 층에서 생성된 유기 물질의 혼합에 더 의존하게 된다. 지표수의 영양소 농도가 낮고 혼합층 이하의 빛 침투가 있는 호수에서는 광합성을 위한 충분한 빛이 있는 중간 깊이에서 1차 생산량이 더 높다.[49] 반면, 투명도가 낮은 다극성 호수는 지표면에 가까운 층에서 1차 생산량이 높으며, 보통 1차 생산과 호흡 사이에 순 자생 균형(GPP > ER)이 있다.[12]
횡방향으로 호수 내 이질성은 개방수 림프구 내 대사율의 차이와 보다 구부러진 주도의 연두구간에 의해 추진된다. 연돌 지역은 대개 육지계와의 근접성 때문에 더 복잡하고 이질적이지만, 또한 낮은 수량과 높은 침전물 대 수량의 비율 때문에 그러하기도 한다. 따라서 연립구(littoral zone)는 온도 변화, 경관 및 하천 유입으로부터 발생하는 영양소 및 유기물 투입, 윈드시어 혼합 및 파동 작용, 지상 식물에서 음영 처리, 퇴적물의 재충전 등에 더 취약하다(그림 1). 또한 연근위 지역은 대게 많은 유기체의 쉼터, 보육원, 먹이 장소 역할을 하는 대식세포의 존재로 인해 서식지의 복잡성이 더 크다. 결과적으로, 연골 부위의 대사율은 대개 높은 단기적 변동성을 가지고 있으며 일반적으로 림프성 대사율보다 크다.[50][11]
호수에 따른 공간적 변화
호수 내 공간적 변동성 외에도, 전리크 대사율과 그 추진력은 호수마다 다르다. 각 호수는 형태계, 유역 특성, 수문학적 특성에 따라 고유한 특성 집합을 가진다. 이러한 특징들은 호수 대사에 직간접적인 영향을 미치며 수채색, 온도, 영양소, 유기 물질, 빛 감쇠, 수직 및 수평 혼합과 같은 호수 조건에 영향을 미친다.
호수는 성분(예: 빛, 영양소, 온도, 유기물)의 상태가 다르기 때문에 호수 간 대사율의 규모와 변동성에 새로운 차이가 나타나고 있다. 이전 섹션(구성요소와의 관계)에서는 이러한 영향력 있는 성분의 변동성에 대응한 대사율의 예상 패턴을 논의하였다. 여기서는 호수 형태 측정, 유역 특성 및 수거 시간의 차이로 매개되는 이들 성분의 차이로 인해 호수 전체에 걸쳐 전유동 신진대사가 어떻게 변화하는지 논의한다.
호수 형태측정법(예: 호수 크기 및 모양)과 유역 특성(예: 토지 이용, 배수 지역, 기후 및 지질 특성)은 호수의 수적 단위당 유기물질과 영양소의 외부 입력의 유동성을 결정한다. 유역 규모와 호수의 용적률(배수율)의 비율이 높아짐에 따라 주변의 육지풍경에서 나오는 영양소와 유기물의 유동성이 일반적으로 증가하게 된다.[51] 즉, 유역이 비교적 큰 작은 호수는 유역이 비교적 적은 큰 호수보다 호수 부피 단위당 외부 영양소 및 유기물 투입량이 많아 1차 생산량과 호흡수가 모두 향상된다. 배수 비율이 작은 호수(즉, 유역 면적과 관련된 상대적인 큰 호수 표면적)에서는 대사 과정이 주변 유역에서 오는 외부 입력에 덜 의존할 것으로 예상된다. 또한, 작은 호수는 바람에 의한 혼합에 덜 노출되고 일반적으로 지상 유기 물질 입력이 더 높아져서 종종 얕은 혼합 깊이와 강화된 빛 감쇄를 초래하여 1차 생산량을 작은 호수의 상위 부분으로 제한한다. 이후 더 높은 호흡률 더 높은 타지성 유기물에 의해 1차 생산 wh를 상회하는 시스템에 들어가(즉 배수 지역 이내에 합성된 외부의 물 몸)된 유역 특성을 가지고 호수를 생각하면, 작은 호수 일반적으로 더 많은 종속 영양(범용 플랫폼<>ER)의 큰 호수보다 순이익 있다.ich는 얕은 호수 층으로 제한된다.

토지 커버, 토지 이용, 지질학적 특성 등 유역 특성은 풍속 노출뿐만 아니라 호수에 유입되는 유기 물질과 영양소의 질에 미치는 영향을 통해 호수 신진대사에 영향을 미친다. 유기 물질 품질은 빛 감쇠에 영향을 줄 수 있으며, 바람 노출과 함께 수호 기둥 전체의 열과 빛 분포에 영향을 줄 수 있다. 농업이 지배하는 경관의 호수는 배수율은 비슷하지만 숲이 지배하는 경관의 호수에 비해 영양소 투입량이 높고 유기물 투입량이 낮다. 따라서 농업이 지배적인 경관의 호수는 산림 지배적인 경관의 호수에 비해 1차 생산률이 높고, 녹조가 많으며, 과도한 대식세포 바이오매스가 예상된다(그림). 그러나 유역 크기와 유역 유형의 효과는 복잡하고 상호 작용적이다. 상대적으로 숲이 우거진 호수는 그늘이 더 많고 바람의 노출로부터 보호되며 또한 많은 양의 알록달록한 유기 물질을 받는다. 따라서 숲이 우거진 작은 호수는 일반적으로 얕은 혼합 층과 빛의 침투가 감소하여 더 유머러스하다. (호수 밖에서 생산되는) 알로크톤성 유기물질의 높은 투입은 박테리아, 동물성 플랑크톤, 어류 등 이질생성 공동체를 자극해 전리호흡률을 높인다. 따라서 숲이 우거진 작은 호수는 순 이질성 호수가 될 가능성이 더 높으며, ER 비율이 호수의 1차 생산률을 초과한다. 반면, 배수율이 낮은 숲이 우거진 호수는 상대적으로 적은 영양소와 유기물을 공급받으며, 일반적으로 맑은 물이 흐르는 호수로 GPP와 ER 비율(표)이 낮다.

호수 대사의 가변성에 영향을 미치는 호수들 간의 또 다른 중요한 차이점은 특히 인간이 집중적으로 관리하는 호수들 사이에서 시스템의 물의 거주시간이다. 호수 수위 및 플러싱률의 변화는 영양소와 유기물 농도, 유기체의 풍부함, 색소 유기물의 광분해와 같은 생태학적 처리율에 영향을 미치며 대사율의 크기와 변동성에 영향을 미친다. 중간 수압 거주 시간(HRT)이 있는 내측호수 또는 호수는 일반적으로 시스템 내 영양소와 유기물질의 유지시간이 높으며, 이는 1차 생산자의 성장과 유기물질의 박테리아 저하를 선호한다.[52] 따라서 이러한 유형의 호수는 같은 영양 상태에 있는 거주 시간이 낮은 호수보다 상대적으로 높고 가변성이 적은 GPP와 ER 비율을 유지할 것으로 예상된다. 반면 HRT가 긴 호수는 호수에 대한 영양소 및 유기물 투입량이 적어 대사량이 줄어들 것으로 예상된다. 마지막으로, 수위 변화가 빈번하고 심한 렌즈콩 시스템과 플러싱 속도가 가속화된 렌즈콩 시스템은 강력한 플러싱 이벤트 중 영양소, 유기물, 해조류가 시스템 밖으로 플러싱되기 때문에 대개 GPP와 ER 비율이 낮은 로틱 시스템에 동적 근접성을 가진다.
일별척도변동
일 단위로 보면 GPP 비율은 광합성 활성 방사선의 다이엘 주기에 가장 큰 영향을 받는 반면 ER은 수온의 변화에 크게 영향을 받는다.[53] 또한 ER 속도는 야간 호흡 패턴에 대한 연구(예: Sadro et al[10] 2014)에서 알 수 있듯이 유기 기질의 양 또는 질과 자가영양 및 이종영양 호흡의 상대적 기여와도 관련이 있다. 예를 들어, 박테리아 소플랑크톤 호흡은 식물성 플랑크톤에 의해 생성된 미생물 용해 유기물질의 높은 가용성 때문에 낮과 밤의 첫 시간에 더 높을 수 있다. 해가 뜨면서 호수에서 1차 생산량이 급격히 증가하여 자주 자가영양(NEP > 0)이 되고, 밤중에 발생한 탄소광물화에서 생성된 용해된 CO2가 감소한다. 이러한 행동은 NEP에서 정점에 도달할 때까지 계속되며, 일반적으로 최대 광도 가용성에 가깝다. 그 후 NEP는 다음날 해가 뜰 때까지 최대 광선 이용 시간 사이에 꾸준히 하락하는 경향이 있다.
구름 덮개와 폭풍과 같은 날씨 차이로 인해 들어오는 빛과 온도의 일상적인 차이는 일차 생산 속도에 영향을 미치고, 호흡은 더 적게 영향을 미친다.[54] 이러한 날씨 변화는 또한 혼합층 깊이의 단기적 변동성을 야기하며, 이는 수직적, 수평적 가스 교환뿐만 아니라 영양소, 유기 물질, 빛의 가용성에 영향을 미친다. 딥믹스는 빛의 이용가능성을 감소시키지만 상층부의 영양소와 유기물 이용가능성을 증가시킨다. 따라서 혼합층 깊이의 단기 변동성이 총 1차 생산량(GPP)에 미치는 영향은 특정 기간 동안 각 호수에 대한 제한 요인에 따라 달라질 것이다. 따라서 더 깊은 혼합 층은 광합성의 영양소와 빛 한계 사이의 균형에 따라 GPP 비율을 증가시키거나 감소시킬 수 있다(그림).
대사율의 반응은 호수에서 일어나는 물리적, 화학적 과정만큼 역동적이지만, 녹조 바이오매스의 변화는 더 긴 기간에 걸쳐 성장과 손실을 수반하는 변동성이 적다. 높은 빛과 영양소의 가용성은 호수의 녹조 발생과 관련이 있다; 이러한 꽃 피우는 동안 GPP 비율은 매우 높고, ER 비율은 대개 GPP 비율만큼 증가하며, GPP와 ER의 균형은 1에 가깝다. 개화 직후부터 GPP는 감소하기 시작하지만 ER 속도는 미분기 유기물질의 높은 이용가능성으로 인해 계속 높아져 물기둥의 용존산소 농도가 빠르게 고갈되어 어류 살처분으로 이어질 수 있다.
연간척도변동
신진대사의 계절적 변화는 온도, 얼음 덮개, 강우, 혼합 및 층화 역학, 지역사회 계승(예: 동물성 플랑크톤에[55] 의한 식물성 플랑크톤 조절)의 계절적 변화에 의해 추진될 수 있다. 호수의 신진대사의 계절적 변화는 계절이 영양소와 유기물의 투입을 어떻게 변화시키는지, 그리고 광의 이용가능성을 어떻게 변화시키는지, 그리고 각 호수의 대사율을 제한하는 요인에 따라 달라질 것이다.
빛은 호수대사의 주요 원인이므로, 빛의 계절성은 호수대사의 계절적 변화를 일으키는 중요한 요인이다. 따라서 광도가 높고 낮이 긴 봄과 여름과 같은 계절에는 GPP 비율이 더 뚜렷해질 것으로 예상된다. 이는 특히 GPP가 가벼운 호수에 대해, 예를 들어, 더 탁하거나 얼룩진 호수에 대해 두드러진다. 광도의 계절성은 또한 ER 비율에도 영향을 미친다. 생태계 호흡수는 대개 GPP 비율과 결합되기 때문에 GPP가 높은 계절은 호수 내에서 생성되는 유기물 증가와 관련된 ER 비율도 더 높게 나타난다. 더욱이 광도가 높은 계절에는 유기물의 광분해 현상이 더욱 뚜렷하게 나타나 미생물 저하를 자극하여 이질성 호흡수를 향상시킨다.

세계의 대부분의 호수는 겨울 동안 얼어서 얼음과 눈이 물기둥의 빛의 침투를 제한하는 저반사성 기간이다.[56] 빛의 제한은 주로 얼음이 아닌 눈 덮개에 의해 발생하며, 이는 1차 생산으로 인해 호수에서 눈 덮개에 매우 민감하게 만든다.[57] 가벼운 제한 외에도 얼음 아래의 낮은 온도는 대사율을 감소시키지만 대사 과정을 멈추기에는 충분하지 않다. 따라서 대부분의 빙수기에는 대사 균형이 대개 음성이어서 용존 산소 고갈로 이어진다. 건조한 기후의 얕은 호수는 겨울 동안 눈이 전혀 오지 않거나 거의 내리지 않기 때문에 송씨 등이 몽골 호수에서 보고한[57] 바와 같이 얼음 아래를 유지하는 일차 생산량은 용해된 산소 고갈을 방지하기에 충분하다. 겨울철에 얼어붙는 세계 호수의 높은 비율에도 불구하고, 주로 시료 채취 기술상의 어려움으로 인해 호수 신진대사에 대한 연구는 거의 이루어지지 않았다.[56][58][57] 적도에 가까운 호수는 높은 위도(온도 및 극지방)의 호수보다 광도와 일조 시간에 관한 계절성이 적다. 따라서, 높은 위도의 호수는 저조도 계절(겨울과 가을)에 일차 생산의 가벼운 제한을 경험할 가능성이 더 높다. 열대지방에서도 계절적인 기온 차이는 높은 위도의 호수에 있어서 그다지 중요하지 않다. 따라서 기온 계절적 변화가 대사율에 미치는 직접적인 영향은 열대호(그림)보다 위도가 높은 호수에서 더 중요하다. 이에 따라 열대호와 아열대호는 위도가 높은 호수에 비해 천문 또는 기상 4계절(봄, 여름, 가을, 겨울)보다는 성층화 및 혼합 역학, 강우(습기, 건기)에 따른 계절적 편차가 더 심하다.

기온과 강우량의 계절적 변화는 물기둥 안정성에 계절적 변화를 가져온다. 낮은 물기둥 안정성 기간 동안 더 깊은 혼합층(호수에 따라 물기둥의 전체 또는 부분 혼합)은 더 깊은 층으로부터 그리고 침전물 재충전을 통해 영양소와 유기 물질의 투입을 증가시켜 빛 이용 가능성을 감소시킨다. 반대로 물기둥 안정성이 강한 시기에는 물기둥에 영양소, 유기물, 관련세균의 내부 하중을 억제하는 한편 침하로 인한 녹조손실은 강화된다. 또한 이 기간 동안의 빛 가용성은 광 블리딩, 낮은 퇴적물 재충전, 낮은 혼합 깊이 등으로 인해 더 높으며, 이는 식물성 플랑크톤을 더 밝은 환경에 노출시킨다. 이 유기물질과 관련된 유기물 이용가능성과 박테리아 바이오매스 증가의 결과로 저수기둥 안정성 기간 동안의 높은 ER 비율은 전 세계 많은 호수에서 보고되었다.[59][60][61] 그러나 이러한 계절적 변화에 대한 일차 생산률은 호수마다 다른 행동을 보였다. 앞에서 말한 바와 같이, 그러한 변화에 대한 대사율의 반응은 각 호수의 1차 생산 제한 요인에 따라 달라질 것이다(그림). 낮은 물기둥 안정성 기간 동안, 영양분이 풍부한 물의 공급은 일부 열대 호수에서 관찰된 것처럼 더 높은 피질 GPP 비율을 초래할 수 있다.[62][63] 반대로, 낮은 물기둥 안정성 기간 동안 GPP 속도는 일부 온대호 및 아열대호에서 관찰된 것처럼 낮은 광 이용가능성에 의해 제한될 수 있다.[64][65] 순대사량 균형은 일반적으로 잘 섞인 계절이 가장 생산적인 시기인 호수에서 조차 탈지위기에 더 부정적이다. 이러한 시스템에서 높은 GPP와 관계없이 ER 속도는 퇴적물과 깊은 수역의 유기 물질 재고 가용성이 증가함에 따라 향상된다.
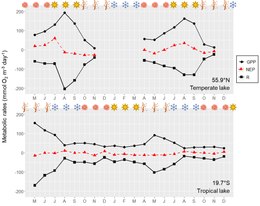
강우량의 계절적 차이도 대사율에 영향을 미친다. 강수량의 증가는 영양소의 증가와 낮은 광 이용성의 균형에 따라 ER 비율을 자극하고 GPP 비율을 자극하거나 억제할 수 있는 호수의 유기 물질과 영양소의 유입을 촉진한다. 반면 강수량이 적으면 수심이 낮아져 임질학적 상태에 영향을 미치며 영양소와 엽록소의 농도를 높이고 수생 환경의 열 안정성을 변화시킨다. 이러한 변화는 또한 ER과 GPP 비율을 향상시킬 수 있다. 따라서, 강우량의 계절적 변화에 대한 대사율의 응답 정도는 호수 형태 측정, 유역 특성, 강우 사건의 강도 및 지속 기간에 따라 달라질 것이다. 북서태평양의 태풍 지역처럼 강한 폭풍에 자주 노출되는 호수는 며칠 동안 지속될 수 있는 강한 강우 이벤트를 받는다.[66] 이러한 폭풍 시즌에는 햇빛을 줄이고 물과 유기체의 홍수로 대사율의 감소가 예상된다. 이러한 감소는 ER 속도보다 GPP에서 더 뚜렷하게 나타나서 보다 이질적인 NEP(GPP < ER)가 될 것으로 예상된다. 예를 들어, 대만의 아열대 호수에서는 태풍 기간 동안 유기 물질 풀(호수 내에서 생성되는 유기 물질)이 자동 치환 기반에서 알록치 기반(호수 밖에서 생성되는 유기 물질)으로 바뀐 후 GPP와 ER 비율의 디커플링이 관찰되었다.[67] 이는 ER 비율이 GPP 비율보다 태풍 교란에 대한 저항성이 높았음을 시사한다.
연차변동
대사율의 연간 변동성은 유역의 광범위한 변화 또는 엘니뇨 남부 진동(ENSO)과 관련된 사건과 같은 방향 및 주기적 기후 변화 및 기후 장애에 의해 주도될 수 있다. 연도 간 유역, 공기 온도 및 강수량의 변화는 호수에 대한 영양소 및 유기 물질 투입물 변경, 가벼운 감쇠, 혼합 역학 및 대사 과정의 직접적인 온도 의존성에 의해 대사율에 영향을 미친다.
강수량의 증가는 호수의 유기물, 영양소, 퇴적물의 외부 부하를 증가시킨다. 게다가 강우량 증가에 의해 촉진된 배출량 증가는 혼합 역학을 변화시키고 유기체의 물리적 플러싱을 유발할 수 있다. 높은 증발률과 관련된 강수량이 낮아지면 수심을 감소시켜 영양소와 엽록소의 농도를 증가시키고 수생 환경의 열 안정성을 변화시킴으로써 림프학적 조건에도 영향을 미친다. 따뜻한 해 동안, 더 강한 물기둥 안정성은 영양소와 유기 물질의 입력을 광자 영역으로 제한한다. 이와는 대조적으로, 추운 기간 동안, 안정적이지 않은 물 기둥이 퇴적물의 재충전 및 깊은 물에서 나오는 영양소와 유기 물질의 투입을 강화시킨다. 이것은 빛 이용 가능성을 낮추는 반면, 영양소와 유기 물질 이용성은 향상시킨다. 따라서, 대사율의 년 간 강수량과 온도 차이의 영향은 이러한 변화의 강도와 지속 기간에 따라 달라질 것이며, 또한 각 수역의 GPP와 ER을 제한하는 요인에 따라 달라질 것이다.
GPP와 ER의 영양소와 유기 물질 제한이 있는 호수에서, 습윤 연도는 풍경으로부터 더 높은 영양소와 유기 물질의 투입으로 인해 GPP와 ER 비율을 향상시킬 수 있다. 이는 지상 투입물이 1차 생산자와 이질적 공동체를 위해 즉시 이용 가능한지 또는 대사 과정이 매우 낮거나 존재하지 않는 더 깊은 물을 통해 호수로 유입되는지에 따라 달라질 것이다. 이 경우 입력은 다음 물 칼럼 혼합 이벤트에서만 사용할 수 있다. 따라서 강우량에 의한 대사율의 증가는 또한 호수의 층화와 혼합 역학, 수문학, 형태계에 따라 달라진다. 반면에, 건조한 해는 낮은 수위와 함께 GPP와 ER 비율을 증가시킬 수 있고, 이것은 더 높은 영양소와 유기 물질 농도로 이어질 것이다. 낮은 수위는 물기둥의 안정성이 떨어지고 퇴적물과의 근접성이 높아져 더 깊은 물에서 나오는 영양소와 유기물의 투입량이 증가한다. 또한 수분 증발을 통한 수위 감소는 농도 효과로 이어진다. 반대로 따뜻한 해에는 물기둥의 안정성이 더 높고, 혼합층의 깊이는 얕아져 있어 혼합층에 대한 영양소와 유기물의 내부 투입을 감소시킨다. 이 시나리오에서 대사율은 상위 혼합층에서 더 낮아질 것이다. 혼합층보다 더 깊은 광자대를 가진 호수에서, 깊은 엽록소와 일치하여 중간 깊이의 대사율이 더 높을 것이다.
주로 광 가용성에 의해 제한된 일차 생산의 호수에서 강우량 증가는 용해된 유기 물질 및 총 부유 물질 증가와 관련하여 낮은 광 가용성으로 이어질 수 있다. 결과적으로, 강우량 증가는 낮은 수준의 GPP와 연관될 것이며, 이것은 자동 치환 생산과 관련된 호흡수를 감소시켜 GPP와 ER 비율을 디커플링하게 될 것이다.[68] 또한 습윤기 동안 모든 색소성 유기 물질 가용성이 증가하면 ER이 높아질 수 있고, 결과적으로 대사 균형이 음성으로 이어질 수 있다(NEP <0).[52]
연간 강수량의 변화는 호수 내 대사율의 공간적 변화에도 영향을 미칠 수 있다. 예를 들어 [52]Williamson과 협력자들은 북아메리카의 초영양 저수지에서 GPP와 ER 비율의 상대적인 공간적 변동성이 습한 것에 비해 건조한 해에 더 높다는 것을 발견했다. 이러한 결과는 내부 부하, 영양 섭취, 침전 및 재부유와 같은 내부 프로세스의 건조 기간 동안의 대사 속도에 대한 높은 관련성을 시사한다.
참고 항목
참조
- ^ Winslow, Luke A.; Zwart, Jacob A.; Batt, Ryan D.; Dugan, Hilary A.; Woolway, R. Iestyn; Corman, Jessica R.; Hanson, Paul C.; Read, Jordan S. (January 2016). "LakeMetabolizer: an R package for estimating lake metabolism from free-water oxygen using diverse statistical models". Inland Waters. 6 (4): 622–636. doi:10.1080/iw-6.4.883. ISSN 2044-2041. S2CID 54811593.
- ^ Hanson, Paul C.; Bade, Darren L.; Carpenter, Stephen R.; Kratz, Timothy K. (2003). "Lake metabolism: Relationships with dissolved organic carbon and phosphorus". Limnology and Oceanography. 48 (3): 1112–1119. Bibcode:2003LimOc..48.1112H. doi:10.4319/lo.2003.48.3.1112. ISSN 1939-5590.
- ^ Bogard, Matthew J.; Giorgio, Paul A. del (2016). "The role of metabolism in modulating CO2 fluxes in boreal lakes". Global Biogeochemical Cycles. 30 (10): 1509–1525. Bibcode:2016GBioC..30.1509B. doi:10.1002/2016GB005463. ISSN 1944-9224.
- ^ Wilkinson, Grace M.; Buelo, Cal D.; Cole, Jonathan J.; Pace, Michael L. (2016). "Exogenously produced CO2 doubles the CO2 efflux from three north temperate lakes". Geophysical Research Letters. 43 (5): 1996–2003. Bibcode:2016GeoRL..43.1996W. doi:10.1002/2016GL067732. ISSN 1944-8007.
- ^ Yvon-Durocher, Gabriel; Caffrey, Jane M.; Cescatti, Alessandro; Dossena, Matteo; Giorgio, Paul del; Gasol, Josep M.; Montoya, José M.; Pumpanen, Jukka; Staehr, Peter A.; Trimmer, Mark; Woodward, Guy (2012). "Reconciling the temperature dependence of respiration across timescales and ecosystem types". Nature. 487 (7408): 472–476. Bibcode:2012Natur.487..472Y. doi:10.1038/nature11205. ISSN 1476-4687. PMID 22722862. S2CID 4422427.
- ^ Porter, John H.; Hanson, Paul C.; Lin, Chau-Chin (2012-02-01). "Staying afloat in the sensor data deluge". Trends in Ecology & Evolution. 27 (2): 121–129. doi:10.1016/j.tree.2011.11.009. ISSN 0169-5347. PMID 22206661.
- ^ Cole, Jonathan J.; Pace, Michael L.; Carpenter, Stephen R.; Kitchell, James F. (December 2000). "Persistence of net heterotrophy in lakes during nutrient addition and food web manipulations". Limnology and Oceanography. 45 (8): 1718–1730. Bibcode:2000LimOc..45.1718C. doi:10.4319/lo.2000.45.8.1718. ISSN 0024-3590.
- ^ Hofmann, Hilmar; Encinas-Fernández, Jorge; Tengberg, Anders; Atamanchuk, Dariia; Peeters, Frank (2016-12-21). "Lake Metabolism: Comparison of Lake Metabolic Rates Estimated from a Diel CO2- and the Common Diel O2-Technique". PLOS ONE. 11 (12): e0168393. Bibcode:2016PLoSO..1168393P. doi:10.1371/journal.pone.0168393. ISSN 1932-6203. PMC 5176309. PMID 28002477.
- ^ Dugan, Hilary A.; Woolway, R. Iestyn; Santoso, Arianto B.; Corman, Jessica R.; Jaimes, Aline; Nodine, Emily R.; Patil, Vijay P.; Zwart, Jacob A.; Brentrup, Jennifer A.; Hetherington, Amy L.; Oliver, Samantha K. (2016-01-01). "Consequences of gas flux model choice on the interpretation of metabolic balance across 15 lakes". Inland Waters. 6 (4): 581–592. doi:10.1080/IW-6.4.836. ISSN 2044-2041.
- ^ a b Sadro, Steven; Holtgrieve, Gordon W.; Solomon, Christopher T.; Koch, Gregory R. (2014). "Widespread variability in overnight patterns of ecosystem respiration linked to gradients in dissolved organic matter, residence time, and productivity in a global set of lakes". Limnology and Oceanography. 59 (5): 1666–1678. Bibcode:2014LimOc..59.1666S. doi:10.4319/lo.2014.59.5.1666. ISSN 1939-5590.
- ^ a b Bogert, Matthew C. Van de; Bade, Darren L.; Carpenter, Stephen R.; Cole, Jonathan J.; Pace, Michael L.; Hanson, Paul C.; Langman, Owen C. (2012). "Spatial heterogeneity strongly affects estimates of ecosystem metabolism in two north temperate lakes". Limnology and Oceanography. 57 (6): 1689–1700. Bibcode:2012LimOc..57.1689V. doi:10.4319/lo.2012.57.6.1689. ISSN 1939-5590.
- ^ a b Obrador, Biel; Staehr, Peter A.; Christensen, Jesper P. C. (2014). "Vertical patterns of metabolism in three contrasting stratified lakes". Limnology and Oceanography. 59 (4): 1228–1240. Bibcode:2014LimOc..59.1228O. doi:10.4319/lo.2014.59.4.1228. ISSN 1939-5590.
- ^ Giling, Darren P.; Staehr, Peter A.; Grossart, Hans Peter; Andersen, Mikkel René; Boehrer, Bertram; Escot, Carmelo; Evrendilek, Fatih; Gómez‐Gener, Lluís; Honti, Mark; Jones, Ian D.; Karakaya, Nusret (2017). "Delving deeper: Metabolic processes in the metalimnion of stratified lakes" (PDF). Limnology and Oceanography. 62 (3): 1288–1306. Bibcode:2017LimOc..62.1288G. doi:10.1002/lno.10504. ISSN 1939-5590.
- ^ Staehr, Peter A.; Bade, Darren; Bogert, Matthew C. Van de; Koch, Gregory R.; Williamson, Craig; Hanson, Paul; Cole, Jonathan J.; Kratz, Tim (2010). "Lake metabolism and the diel oxygen technique: State of the science". Limnology and Oceanography: Methods. 8 (11): 628–644. doi:10.4319/lom.2010.8.0628. ISSN 1541-5856.
- ^ Bender, Michael; Grande, Karen; Johnson, Kenneth; Marra, John; Williams, Peter J. LeB.; Sieburth, John; Pilson, Michael; Langdon, Chris; Hitchcock, Gary (September 1987). "A comparison of four methods for determining planktonic community production1". Limnology and Oceanography. 32 (5): 1085–1098. Bibcode:1987LimOc..32.1085B. doi:10.4319/lo.1987.32.5.1085. ISSN 0024-3590.
- ^ a b Stets, Edward G.; Striegl, Robert G.; Aiken, George R.; Rosenberry, Donald O.; Winter, Thomas C. (2009). "Hydrologic support of carbon dioxide flux revealed by whole-lake carbon budgets". Journal of Geophysical Research: Biogeosciences. 114 (G1): G01008. Bibcode:2009JGRG..114.1008S. doi:10.1029/2008JG000783. ISSN 2156-2202.
- ^ Lovett, Gary M.; Cole, Jonathan J.; Pace, Michael L. (2006-02-01). "Is Net Ecosystem Production Equal to Ecosystem Carbon Accumulation?". Ecosystems. 9 (1): 152–155. doi:10.1007/s10021-005-0036-3. ISSN 1435-0629. S2CID 5890190.
- ^ Godwin, Sean C.; Jones, Stuart E.; Weidel, Brian C.; Solomon, Christopher T. (2014). "Dissolved organic carbon concentration controls benthic primary production: Results from in situ chambers in north-temperate lakes". Limnology and Oceanography. 59 (6): 2112–2120. Bibcode:2014LimOc..59.2112G. doi:10.4319/lo.2014.59.6.2112. ISSN 1939-5590.
- ^ a b Bogert, Matthew C. Van de; Bade, Darren L.; Carpenter, Stephen R.; Cole, Jonathan J.; Pace, Michael L.; Hanson, Paul C.; Langman, Owen C. (2012). "Spatial heterogeneity strongly affects estimates of ecosystem metabolism in two north temperate lakes". Limnology and Oceanography. 57 (6): 1689–1700. Bibcode:2012LimOc..57.1689V. doi:10.4319/lo.2012.57.6.1689. ISSN 1939-5590.
- ^ Obrador, Biel; Staehr, Peter A.; Christensen, Jesper P. C. (2014). "Vertical patterns of metabolism in three contrasting stratified lakes". Limnology and Oceanography. 59 (4): 1228–1240. Bibcode:2014LimOc..59.1228O. doi:10.4319/lo.2014.59.4.1228. ISSN 1939-5590.
- ^ Giling, Darren P.; Staehr, Peter A.; Grossart, Hans Peter; Andersen, Mikkel René; Boehrer, Bertram; Escot, Carmelo; Evrendilek, Fatih; Gómez‐Gener, Lluís; Honti, Mark; Jones, Ian D.; Karakaya, Nusret (2017). "Delving deeper: Metabolic processes in the metalimnion of stratified lakes" (PDF). Limnology and Oceanography. 62 (3): 1288–1306. Bibcode:2017LimOc..62.1288G. doi:10.1002/lno.10504. ISSN 1939-5590.
- ^ Yvon-Durocher, Gabriel; Caffrey, Jane M.; Cescatti, Alessandro; Dossena, Matteo; Giorgio, Paul del; Gasol, Josep M.; Montoya, José M.; Pumpanen, Jukka; Staehr, Peter A.; Trimmer, Mark; Woodward, Guy (2012). "Reconciling the temperature dependence of respiration across timescales and ecosystem types". Nature. 487 (7408): 472–476. Bibcode:2012Natur.487..472Y. doi:10.1038/nature11205. ISSN 1476-4687. PMID 22722862. S2CID 4422427.
- ^ Scharfenberger, Ulrike; Jeppesen, Erik; Beklioğlu, Meryem; Søndergaard, Martin; Angeler, David G.; Çakıroğlu, Ayşe İdil; Drakare, Stina; Hejzlar, Josef; Mahdy, Aldoushy; Papastergiadou, Eva; Šorf, Michal (2019). "Effects of trophic status, water level, and temperature on shallow lake metabolism and metabolic balance: A standardized pan-European mesocosm experiment". Limnology and Oceanography. 64 (2): 616–631. Bibcode:2019LimOc..64..616S. doi:10.1002/lno.11064. ISSN 1939-5590.
- ^ "USGS WRI99-4007 Review of Phosphorus Control Measures in the United States and Their Effects on Water Quality". pubs.usgs.gov. Retrieved 2020-06-28.
- ^ Kogawa, Ana Carolina; Cernic, Beatriz Gamberini; do Couto, Leandro Giovanni Domingos; Salgado, Hérida Regina Nunes (2017). "Synthetic detergents: 100years of history". Saudi Pharmaceutical Journal. 25 (6): 934–938. doi:10.1016/j.jsps.2017.02.006. ISSN 1319-0164. PMC 5605839. PMID 28951681.
- ^ Elser, James J.; Bracken, Matthew E. S.; Cleland, Elsa E.; Gruner, Daniel S.; Harpole, W. Stanley; Hillebrand, Helmut; Ngai, Jacqueline T.; Seabloom, Eric W.; Shurin, Jonathan B.; Smith, Jennifer E. (2007). "Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems". Ecology Letters. 10 (12): 1135–1142. doi:10.1111/j.1461-0248.2007.01113.x. hdl:1903/7447. ISSN 1461-0248. PMID 17922835.
- ^ Litchman, Elena; Steiner, Daniel; Bossard, Peter (2003). "Photosynthetic and growth responses of three freshwater algae to phosphorus limitation and daylength". Freshwater Biology. 48 (12): 2141–2148. doi:10.1046/j.1365-2427.2003.01157.x. ISSN 1365-2427.
- ^ Staehr, Peter A.; Brighenti, Ludmila S.; Honti, Mark; Christensen, Jesper; Rose, Kevin C. (2016). "Global patterns of light saturation and photoinhibition of lake primary production". Inland Waters. 6 (4): 593–607. doi:10.1080/IW-6.4.888. ISSN 2044-2041. S2CID 54945617.
- ^ Zwart, Jacob A.; Solomon, Christopher T.; Jones, Stuart E. (2015). "Phytoplankton traits predict ecosystem function in a global set of lakes". Ecology. 96 (8): 2257–2264. doi:10.1890/14-2102.1. ISSN 1939-9170. PMID 26405750.
- ^ Karlsson, Jan; Byström, Pär; Ask, Jenny; Ask, Per; Persson, Lennart; Jansson, Mats (2009). "Light limitation of nutrient-poor lake ecosystems". Nature. 460 (7254): 506–509. Bibcode:2009Natur.460..506K. doi:10.1038/nature08179. ISSN 1476-4687. PMID 19626113. S2CID 4360994.
- ^ Cory, Rose M.; Ward, Collin P.; Crump, Byron C.; Kling, George W. (2014). "Sunlight controls water column processing of carbon in arctic fresh waters". Science. 345 (6199): 925–928. Bibcode:2014Sci...345..925C. doi:10.1126/science.1253119. ISSN 0036-8075. PMID 25146289. S2CID 206556629.
- ^ a b c Seekell, David A.; Lapierre, Jean-François; Ask, Jenny; Bergström, Ann-Kristin; Deininger, Anne; Rodríguez, Patricia; Karlsson, Jan (2015). "The influence of dissolved organic carbon on primary production in northern lakes". Limnology and Oceanography. 60 (4): 1276–1285. doi:10.1002/lno.10096. ISSN 1939-5590.
- ^ a b c Seekell, David A.; Lapierre, Jean-François; Karlsson, Jan (2015-07-14). "Trade-offs between light and nutrient availability across gradients of dissolved organic carbon concentration in Swedish lakes: implications for patterns in primary production". Canadian Journal of Fisheries and Aquatic Sciences. doi:10.1139/cjfas-2015-0187. hdl:1807/69824.
- ^ a b c d Seekell, David A.; Lapierre, Jean-François; Cheruvelil, Kendra Spence (2018). "A geography of lake carbon cycling". Limnology and Oceanography Letters. 3 (3): 49–56. doi:10.1002/lol2.10078. ISSN 2378-2242.
- ^ Kelly, Patrick T.; Solomon, Christopher T.; Zwart, Jacob A.; Jones, Stuart E. (2018). "A Framework for Understanding Variation in Pelagic Gross Primary Production of Lake Ecosystems". Ecosystems. 21 (7): 1364–1376. doi:10.1007/s10021-018-0226-4. ISSN 1435-0629. S2CID 31266760.
- ^ Bergström, Ann-Kristin; Karlsson, Jan (2019). "Light and nutrient control phytoplankton biomass responses to global change in northern lakes". Global Change Biology. 25 (6): 2021–2029. doi:10.1111/gcb.14623. ISSN 1365-2486.
- ^ Kelly, Patrick T.; Solomon, Christopher T.; Zwart, Jacob A.; Jones, Stuart E. (2018-11-01). "A Framework for Understanding Variation in Pelagic Gross Primary Production of Lake Ecosystems". Ecosystems. 21 (7): 1364–1376. doi:10.1007/s10021-018-0226-4. ISSN 1435-0629. S2CID 31266760.
- ^ Jansson, Mats; Karlsson, Jan; Jonsson, Anders (2012). "Carbon dioxide supersaturation promotes primary production in lakes". Ecology Letters. 15 (6): 527–532. doi:10.1111/j.1461-0248.2012.01762.x. ISSN 1461-0248. PMID 22420750.
- ^ Verspagen, Jolanda M. H.; Waal, Dedmer B. Van de; Finke, Jan F.; Visser, Petra M.; Donk, Ellen Van; Huisman, Jef (2014-08-13). "Rising CO2 Levels Will Intensify Phytoplankton Blooms in Eutrophic and Hypertrophic Lakes". PLOS ONE. 9 (8): e104325. Bibcode:2014PLoSO...9j4325V. doi:10.1371/journal.pone.0104325. ISSN 1932-6203. PMC 4132121. PMID 25119996.
- ^ Tank, Suzanne E.; Lesack, Lance F. W.; McQueen, Donald J. (2009). "Elevated pH regulates bacterial carbon cycling in lakes with high photosynthetic activity". Ecology. 90 (7): 1910–1922. doi:10.1890/08-1010.1. ISSN 1939-9170. PMID 19694139.
- ^ Christensen, Jesper P. A.; Sand‐Jensen, Kaj; Staehr, Peter A. (2013). "Fluctuating water levels control water chemistry and metabolism of a charophyte-dominated pond". Freshwater Biology. 58 (7): 1353–1365. doi:10.1111/fwb.12132. ISSN 1365-2427.
- ^ Comte, Jérôme; Langenheder, Silke; Berga, Mercè; Lindström, Eva S. (2017). "Contribution of different dispersal sources to the metabolic response of lake bacterioplankton following a salinity change". Environmental Microbiology. 19 (1): 251–260. doi:10.1111/1462-2920.13593. ISSN 1462-2920. PMID 27871136. S2CID 13011521.
- ^ Jiang, Hongchen; Dong, Hailiang; Yu, Bingsong; Liu, Xinqi; Li, Yiliang; Ji, Shanshan; Zhang, Chuanlun L. (2007). "Microbial response to salinity change in Lake Chaka, a hypersaline lake on Tibetan plateau". Environmental Microbiology. 9 (10): 2603–2621. doi:10.1111/j.1462-2920.2007.01377.x. ISSN 1462-2920. PMID 17803783.
- ^ Toepfer, Conrad; Barton, Michael (1992-09-01). "Influence of salinity on the rates of oxygen consumption in two species of freshwater fishes, Phoxinus erythrogaster (family Cyprinidae), and Fundulus catenatus (family Fundulidae)". Hydrobiologia. 242 (3): 149–154. doi:10.1007/BF00019963. ISSN 1573-5117. S2CID 21293418.
- ^ Alfonso, María Belén; Brendel, Andrea Soledad; Vitale, Alejandro José; Seitz, Carina; Piccolo, María Cintia; Perillo, Gerardo Miguel Eduardo (2018). "Drivers of Ecosystem Metabolism in Two Managed Shallow Lakes with Different Salinity and Trophic Conditions: The Sauce Grande and La Salada Lakes (Argentina)". Water. 10 (9): 1136. doi:10.3390/w10091136.
- ^ Moreira-Turcq, Patricia F. (2000-06-01). "Impact of a low salinity year on the metabolism of a hypersaline coastal lagoon (Brazil)". Hydrobiologia. 429 (1): 133–140. doi:10.1023/A:1004037624787. ISSN 1573-5117. S2CID 20968144.
- ^ Dugan, Hilary A.; Bartlett, Sarah L.; Burke, Samantha M.; Doubek, Jonathan P.; Krivak-Tetley, Flora E.; Skaff, Nicholas K.; Summers, Jamie C.; Farrell, Kaitlin J.; McCullough, Ian M.; Morales-Williams, Ana M.; Roberts, Derek C. (2017-04-25). "Salting our freshwater lakes". Proceedings of the National Academy of Sciences. 114 (17): 4453–4458. Bibcode:2017PNAS..114.4453D. doi:10.1073/pnas.1620211114. ISSN 0027-8424. PMC 5410852. PMID 28396392.
- ^ Corsi, Steven R.; Graczyk, David J.; Geis, Steven W.; Booth, Nathaniel L.; Richards, Kevin D. (2010-10-01). "A Fresh Look at Road Salt: Aquatic Toxicity and Water-Quality Impacts on Local, Regional, and National Scales". Environmental Science & Technology. 44 (19): 7376–7382. Bibcode:2010EnST...44.7376C. doi:10.1021/es101333u. ISSN 0013-936X. PMC 2947309. PMID 20806974.
- ^ a b Coloso, James J; Cole, Jonathan J; Hanson, Paul C; Pace, Michael L (2008). "Depth-integrated, continuous estimates of metabolism in a clear-water lake". Canadian Journal of Fisheries and Aquatic Sciences. 65 (4): 712–722. doi:10.1139/f08-006. ISSN 0706-652X.
- ^ Cavalcanti, J. Rafael; da Motta-Marques, David; Fragoso, Carlos Ruberto (2016). "Process-based modeling of shallow lake metabolism: Spatio-temporal variability and relative importance of individual processes". Ecological Modelling. 323: 28–40. doi:10.1016/j.ecolmodel.2015.11.010. ISSN 0304-3800.
- ^ Jones, Stuart E.; Zwart, Jacob A.; Kelly, Patrick T.; Solomon, Christopher T. (2018). "Hydrologic setting constrains lake heterotrophy and terrestrial carbon fate". Limnology and Oceanography Letters. 3 (3): 256–264. doi:10.1002/lol2.10054. ISSN 2378-2242.
- ^ a b c Williamson, Tanner J.; Vanni, Michael J.; Renwick, William H. (2020). "Spatial and Temporal Variability of Nutrient Dynamics and Ecosystem Metabolism in a Hyper-eutrophic Reservoir Differ Between a Wet and Dry Year". Ecosystems. 24: 68–88. doi:10.1007/s10021-020-00505-8. ISSN 1435-0629. S2CID 216559711.
- ^ Yvon-Durocher, Gabriel; Caffrey, Jane M.; Cescatti, Alessandro; Dossena, Matteo; Giorgio, Paul del; Gasol, Josep M.; Montoya, José M.; Pumpanen, Jukka; Staehr, Peter A.; Trimmer, Mark; Woodward, Guy (2012). "Reconciling the temperature dependence of respiration across timescales and ecosystem types". Nature. 487 (7408): 472–476. Bibcode:2012Natur.487..472Y. doi:10.1038/nature11205. ISSN 1476-4687. PMID 22722862. S2CID 4422427.
- ^ Rose, Kevin C.; Winslow, Luke A.; Read, Jordan S.; Read, Emily K.; Solomon, Christopher T.; Adrian, Rita; Hanson, Paul C. (2014). "Improving the precision of lake ecosystem metabolism estimates by identifying predictors of model uncertainty". Limnology and Oceanography: Methods. 12 (5): 303–312. doi:10.4319/lom.2014.12.303. ISSN 1541-5856.
- ^ Lampert, Winfried; Fleckner, Walter; Rai, Hakumat; Taylor, Barbara E. (1986). "Phytoplankton control by grazing zooplankton: A study on the spring clear-water phase1". Limnology and Oceanography. 31 (3): 478–490. Bibcode:1986LimOc..31..478L. doi:10.4319/lo.1986.31.3.0478. ISSN 1939-5590.
- ^ a b Obertegger, Ulrike; Obrador, Biel; Flaim, Giovanna (2017). "Dissolved oxygen dynamics under ice: Three winters of high-frequency data from Lake Tovel, Italy". Water Resources Research. 53 (8): 7234–7246. Bibcode:2017WRR....53.7234O. doi:10.1002/2017WR020599. hdl:2445/123119. ISSN 1944-7973.
- ^ a b c Song, Shuang; Li, Changyou; Shi, Xiaohong; Zhao, Shengnan; Tian, Weidong; Li, Zhijun; Bai, Yila; Cao, Xiaowei; Wang, Qingkai; Huotari, Jussi; Tulonen, Tiina (2019). "Under-ice metabolism in a shallow lake in a cold and arid climate". Freshwater Biology. 64 (10): 1710–1720. doi:10.1111/fwb.13363. hdl:10138/327111. ISSN 1365-2427.
- ^ Block, Benjamin D.; Denfeld, Blaize A.; Stockwell, Jason D.; Flaim, Giovanna; Grossart, Hans-Peter F.; Knoll, Lesley B.; Maier, Dominique B.; North, Rebecca L.; Rautio, Milla; Rusak, James A.; Sadro, Steve (2019). "The unique methodological challenges of winter limnology". Limnology and Oceanography: Methods. 17 (1): 42–57. doi:10.1002/lom3.10295. ISSN 1541-5856.
- ^ Biddanda, Bopaiah A.; Cotner, James B. (2002). "Love Handles in Aquatic Ecosystems: The Role of Dissolved Organic Carbon Drawdown, Resuspended Sediments, and Terrigenous Inputs in the Carbon Balance of Lake Michigan". Ecosystems. 5 (5): 431–445. doi:10.1007/s10021-002-0163-z. ISSN 1432-9840. S2CID 25449277.
- ^ a b Staehr, Peter A.; Sand-Jensen, Kaj (January 2007). "Temporal dynamics and regulation of lake metabolism". Limnology and Oceanography. 52 (1): 108–120. Bibcode:2007LimOc..52..108S. doi:10.4319/lo.2007.52.1.0108. ISSN 1939-5590.
- ^ a b Brighenti, Ludmila Silva; Staehr, Peter Anton; Gagliardi, Laura Martins; Brandão, Luciana Pena Mello; Elias, Eliane Côrrea; de Mello, Nelson Azevedo Santos Teixeira; Barbosa, Francisco Antônio Rodrigues; Bezerra-Neto, José Fernandes (June 2015). "Seasonal Changes in Metabolic Rates of Two Tropical Lakes in the Atlantic Forest of Brazil". Ecosystems. 18 (4): 589–604. doi:10.1007/s10021-015-9851-3. ISSN 1435-0629. S2CID 16135518.
- ^ Brighenti, Ludmila S.; Staehr, Peter A.; Brandão, Luciana P. M.; Barbosa, Francisco A. R.; Bezerra‐Neto, José Fernandes (2018). "Importance of nutrients, organic matter and light availability on epilimnetic metabolic rates in a mesotrophic tropical lake". Freshwater Biology. 63 (9): 1143–1160. doi:10.1111/fwb.13123. ISSN 1365-2427.
- ^ Gagliardi, Laura Martins; Brighenti, Ludmila Silva; Staehr, Peter Anton; Barbosa, Francisco Antônio Rodrigues; Bezerra-Neto, José Fernandes (2019). "Reduced Rainfall Increases Metabolic Rates in Upper Mixed Layers of Tropical Lakes". Ecosystems. 22 (6): 1406–1423. doi:10.1007/s10021-019-00346-0. ISSN 1435-0629. S2CID 71146859.
- ^ Staehr, Peter A.; Sand-Jensen, Kaj; Raun, Ane L.; Nilsson, Bertel; Kidmose, Jacob (2010). "Drivers of metabolism and net heterotrophy in contrasting lakes". Limnology and Oceanography. 55 (2): 817–830. Bibcode:2010LimOc..55..817S. doi:10.4319/lo.2010.55.2.0817. ISSN 1939-5590.
- ^ Tonetta, Denise; Staehr, Peter Anton; Schmitt, Rafael; Petrucio, Mauricio Mello (2016). "Physical conditions driving the spatial and temporal variability in aquatic metabolism of a subtropical coastal lake". Limnologica. 58: 30–40. doi:10.1016/j.limno.2016.01.006. ISSN 0075-9511.
- ^ Jones, Stuart E.; Kratz, Timothy K.; Chiu, Chih-Yu; McMAHON, Katherine D. (2009). "Influence of typhoons on annual CO2 flux from a subtropical, humic lake". Global Change Biology. 15 (1): 243–254. Bibcode:2009GCBio..15..243J. doi:10.1111/j.1365-2486.2008.01723.x. ISSN 1365-2486.
- ^ Tsai, Jeng-Wei; Kratz, Timothy K.; Hanson, Paul C.; Kimura, Nobuaki; Liu, Wen-Cheng; Lin, Fang-Pan; Chou, Hsiu-Mei; Wu, Jiunn-Tzong; Chiu, Chih-Yu (2011). "Metabolic changes and the resistance and resilience of a subtropical heterotrophic lake to typhoon disturbance". Canadian Journal of Fisheries and Aquatic Sciences. 68 (5): 768–780. doi:10.1139/f2011-024. ISSN 0706-652X.
- ^ Richardson, David C.; Carey, Cayelan C.; Bruesewitz, Denise A.; Weathers, Kathleen C. (2017). "Intra- and inter-annual variability in metabolism in an oligotrophic lake". Aquatic Sciences. 79 (2): 319–333. doi:10.1007/s00027-016-0499-7. ISSN 1420-9055.


