드로소필라메틀레리
Drosophila mettleri| 드로소필라메틀레리 | |
|---|---|
| 과학적 분류 | |
| 킹덤: | |
| 망울: | |
| 클래스: | |
| 순서: | |
| 패밀리: | |
| 속: | |
| 종: | D. 메틀레리 |
| 이항식 이름 | |
| 드로소필라 메틀러리 | |
흔히 소노란 사막의 파리로 알려진 드로소필라 메틀러리는 드로소필라 속에 있는 파리다. 이 종은 북아메리카에서 발견되며 캘리포니아의 남쪽 해안과 멕시코에 가장 집중되어 있다. D. 메틀레리는 식물 숙주, 즉 사구로와 카돈 선인장에 의존한다. 그러므로, 그들은 건조하고 사막적인 환경에서 가장 널리 퍼져있다.[1] 그것은 선인장 숙주의 썩어가는 액체에서 발견되는 화학물질을 해독할 수 있어, 그렇지 않으면 치명적인 토양을 보금자리로 사용할 수 있다.
D. 메틀레리는 1961년 10월 바하 캘리포니아주 산펠리페에서 여러 종의 드로소필라를 유인하기 위해 바나나를 사용한 실험 중에 발견되었다.[2] 소노란 사막의 파리들 사이의 물리적인 지리적 장벽 때문에, 유전자 흐름은 분화를 설명한다.
그것은 썩은 선인장에 의해 배설된 액체로 축축한 토양과 괴사체에서 번식한다. 썩은 선인장에서 발견된 이 액체는 다른 디프테란의 장에 독성이 있는 화학물질을 함유하고 있음에도 불구하고 D. 메틀레리 유충의 영양소 역할을 한다.[3] 그 구애를 노래하는 노래와 코퍼레이션 플러그와 같은 파리 종의 구애 행동의 몇 가지 측면은 이 파리 안에서 성적 선택 방법을 설명한다.
설명
D. 메틀레리와 D. 니그로스피라큘라의 차이점
소노란 사막파리종 중 D.메틀레리는 행동하고 번식하며 D. 니그로스피라큘라와 가장 유사하게 분포한다. 널리 퍼져있는 지리적 위치에 있는 선인장에서 D. 메틀레리가 채취되면 D. 니그로스피라큘라와 함께 격리된다. 두 종 모두 숙주 식물의 독성 수액을 착취한다. 그러나 D. nigrospiracula와 달리 D.metleri는 D. nigrospiracula에 의해 악용되지 않는 지역에 둥지를 틀 수 있는 보다 효율적인 해독 시스템을 가지고 있다.[1]
D. 메틀레리는 다친 선인장의 수액에 촉촉한 토양에서 번식하는 것을 선호하며, D. 니그로스피라큘라는 선인장 자체의 조직에서 발견되어 D. 메틀레리에 비해 덜 농축된 독소 농도를 받는다. 유전체 생물학적 저항성을 위한 유전자 인코딩의 상향 조절은 다른 종에 비해 D. 메틀레리의 전문적이고 고효율적인 해독 능력에 기인한다. 이것은 이 종들이 유충과 성인에게 치명적인 번식지를 사용할 수 있게 해준다.[1]
공급 차이
D. 니그로스피라큘라는 오로지 식물 자체에 선인장 수액을 착취한다(즉, 선인장의 조직에서). 그러나 파리는 주변 토양에서 발견되는 고농축 수액을 해독하는 특화된 능력을 가지고 있기 때문에 D. 메틀레리 성인은 선인장과 인근 토양에 모두 위치하고 있다. 암컷은 번식지(토양)에 더 많이 위치하는 반면, 수컷은 양쪽에 위치한다.[4]
신체 구분
표현적 차이와 유사점은 D. 메틀러리와 D. 니그로스피라큘라를 구별하기 위해 사용될 수 있다.[4] 이 두 종 사이의 유사점에는 커다란 몸체, 검은 사체, 그리고 길쭉한 눈과 극명한 대조를 이루는 가늘고 정의된 볼이 있다.[4] 이들 사이의 해독에 사용될 수 있는 이 두 종 사이의 차이점에는 D. 메틀레리가 가지고 있는 특징과 D. 니그로스피라큘라가 가지고 있지 않은 특징들이 있다: 생식기 부위의 발가락 확장, "v"자 모양으로 각진 전면 폴리노스, 덜 숙성된 생식체물리학, 그리고 앞면에만 뿔이 있다(뿔이 없다). 후부([4]후부)

분배
열로 인한 분배
이 종은 특히 멕시코와 애리조나에서 북아메리카 사막에서 가장 흔하게 발견된다. 이 종은 또한 가시 돋친 배에서 서식하는 캘리포니아 남부 해안에서도 발견된다.[5] 소노란 사막에서 발견된 드로소필라 4종 중 D. 메틀레리는 내열성이 가장 낮다.[6] D. 메틀레리는 사계절 내내 풍부하지만 여름 기간에는 덜 만연한다. 사막의 건조한 환경에 있는 선인장의 괴사 조직은 강렬한 열 조건을 특징으로 하기 때문에, D. 메틀레리의 번식 목적으로 선인장의 인근 토양을 이용할 수 있는 능력은 이 종의 번데기가 다른 드로필라 사막종보다 생존율이 높은 선택적 이점을 제공한다.[7] 드로소필라의 다른 종들은 극심한 조건 때문에 소노란 사막의 열기에 덜 성공한다. 그들이 땅을 파고들지 못하는 것은 번데기 생존을 증가시키는데 해로운 것으로 판명되었지만, D. 메틀러리는 번데기 생존을 위한 틈새 환경을 이용하는 능력에 있어서 진화적인 이점을 가지고 있다. 선인장의 괴사조직이 다른 드로소필라 종의 소노란 사막의 토양과 유사한 보호환경 역할을 한다는 가설은 반증되었다. 다른 드로소필라는 열이 높은 선인장의 괴사 조직에서 번식지 때문에 열 허용오차가 높은 반면, D. 메틀레리는 그러한 열 허용오차(선인장 토양의 열 노출이 적은 영역을 이용할 수 있는 능력)에 진화적 이점이 없기 때문에 열 허용오차가 낮다.[8]

지리적 장벽의 영향
소노란 사막이 포함하는 지역의 지형은 괴사성 부패를 먹고 사는 사막 드로소필라 종들 사이에서 유전적 분화를 일으킨다. 소노란 사막은 바하 캘리포니아의 반도 지역과 캘리포니아의 본토 지역을 통합한다. 반도가 형성된 후, 반도와 본토 사이에 놓여 있는 캘리포니아 만은 곤충과 육지 포유류에게 기회를 분산시키는 장벽 역할을 했다. 또 다른 물리적 지리적 장벽은 한반도와 본토 사이에 있는 큰 모래언덕인 그란 데시오르 드 알테르가 있다. 이러한 물리적인 지리적 장벽은 소노란 사막 파리들 사이에서 유전자 흐름의 원인이다. D. 메틀레리와 D. 니그로스피라큘라의 알로지에 대한 연구는 소노란 사막 지역들 간의 차이를 다르게 시사하는 핵 표지의 차이를 보여준다. D. 니그로스피라큘라에는, 그들이 사막의 반도와 본토 지역들 사이에 불균형하게 위치해 있을 때, 그 종들 내에서의 차이를 보여주는 표지가 있었다. 그러나 D. mettleri에서는 한반도에서 발견된 종족과 본토에서 발견된 종족 간에 그러한 핵 표식 차이는 없었다. 이러한 차이는 디 메틀러리의 편차가 디딤돌 가설을 통해 뒷받침될 수 있음을 보여준다.[9]
유전학적으로 가장 가까운 D. 메틀레리의 친척들은 각각 소노란 사막 지역에 서식하지 않기 때문에 소노란 사막 지역에 서식하는 4종의 드로소필라는 사막 내의 분화 대신 평행 진화를 경험했다. D. 메틀레리 이외의 소노란 지역에 고유하게 분포하는 종은 D. 니그로스피라큘라, D. 모하벤시스, D. 멜라노가스터 등이다.[10]

해비타트
숙주식물
주요 숙주 식물로는 사구로(카네기아 기간테아), 카돈(파키체레우스 프링글리), 세니타(로프호체레우스 쇼티이) 선인장 등이 있다.
D. 메틀레리는 사구로와 카돈 선인장에서 가장 흔하게 발견되며, 부분적으로 이러한 선인장의 더 큰 크기와 시원한 내부 환경을 위해 사용된다.[11] 게다가, 선인장의 이 종들은 더 많은 비옥한 물질을 토양에 방출한다; 이 농축된 토양은 보금자리 역할을 한다. 이들 선인장의 괴사조직은 먹이와 보금자리 장소로도 이용될 수 있지만, 이러한 선인장의 썩은 패치의 착취는 다른 사막의 드로필라 종에 비해 D. 메틀레리에서 덜 흔하다.[12]
회전 패치에 의한 호스트 플랜트 선택
소노란 사막 드로소필라 각 종과 관련된 특징은 숙주 선인장 선택이다. 이 파리들은 각각 다른 숙주 공장을 자원(생종, 먹이)으로 이용하기로 선택한다. 숙주 선인장의 선택은 이들 각 종이 이용할 수 있는 자원의 양에 중요하며 종들의 성장과 발전에 역할을 한다.
호스트 선택으로 이어지는 기타 요인
각각의 파리 종은 영양분을 얻기 위해 선인장의 괴사성 로트 패치를 이용한다. 가장 큰 괴사 패치 크기는 카돈, 사과로, 오르간파이프, 세니타 순으로 정렬되어 있다. 선인장의 크기와 팔 길이는 괴사체의 크기에 영향을 미치지 않았다. 회전 패치 크기가 공간 요인에 의해 규제되는 대신, 숙주 공장의 성장의 계절적 차이가 얼마나 크고 얼마나 많은 괴사 패치가 있는가를 결정하였다. 또한, 일시적 요인들은 괴사성 패치가 지속되는 시간을 결정했다. 봄철에는 패치가 가장 적게, 여름에는 패치가 가장 많이 발생한다. 4개의 소노란 사막 드로소필라의 다른 모든 종들은 숙주의 괴사체의 자원 풍부함과 인구 규모 사이에 상관관계를 보였지만, D. 메틀레리에 대해서는 이렇다 할 뚜렷한 상관관계가 발견되지 않았다. D. mettleri는 괴사성 패치와 썩어가는 선인장 근처의 흙을 모두 착취한다; 게다가, 이 종은 다양한 숙주에서 번성할 수 있기 때문에 주어진 공간 지역 아래에서 이 파리들을 더 많이 분산시킬 수 있다. 숙주 선인장의 전반적인 발달 기간과 이용가능성은 유충의 성장과 성인의 생식 성공에 큰 영향을 미친다. 유충의 경우 최적의 숙주 조건은 유충이 더 크고 건강한 성체로 자랄 수 있는 충분한 시간을 주기 때문에 장수 괴사성 플롯을 수반한다. 성인의 경우, 성인은 알을 낳을 수 있는 정기적인 장소를 갖게 되기 때문에 더 규칙적으로 성장하는 숙주가 적절한 조건에 포함된다.[6]
방어 메커니즘
숙주 선인장에는 독성 화학물질과 다른 방법으로 파리가 선인장을 먹는 것을 막는 화합물이 포함되어 있다. D. 메틸러리는 이 독성 화합물을 해독할 수 있게 하는 유전자를 포함하고 있어 선인장의 영양분을 공급한다. 이 유전자들은 이질 생물학적 저항의 진화와 연관되어 있다.
인생사
선인장 주변 토양에 존재하는 고농도의 알칼로이드를 썩은 점으로 해독할 수 있는 D메틀러의 독특한 능력은 성인 암컷이 알을 낳고 같은 보금자리에 유충을 지탱할 수 있게 한다. 같은 지리적 지역에 서식하는 다른 사막의 드로소필라 종은 이러한 능력이 부족하여 암컷들이 둥지 터를 찾는데 있어서 직면하는 경쟁의 양을 줄인다. 1종(D. mazavensis)은 선인장 토양에서 약간만 예민한 유충과 달리 높은 알칼로이드 농도를 견디지 못하는 성인으로 구성되어 있다. 이것은 토양에서 유충 퇴적물을 형성하도록 장려할 수 있지만, 이러한 토양 조건에서는 성인 암컷이 생존할 수 없기 때문에 그들이 토양에 둥지를 틀지 못하게 하고, 따라서 적절한 보금자리를 찾는데 있어서 D. 메틀레리가 직면하는 경쟁의 양을 감소시킨다.[13]
식량자원
로트 소재
드로소필라의 다른 사막종과 마찬가지로 D. 메틀레리는 선인장의 풍부한 육즙을 먹이 기질로서 의존한다. 선인장의 괴사 조직에서 발견되는 즙은 토양 내 배출량이 적거나 희박해지면 먹이 기질로도 쓰인다.[14]
효모와 박테리아
D. 메틀러리가 착취하는 선인장의 영역(토양에서 발출되지 않은 배출물)은 '회전 주머니'이다. 이 주머니들은 선인장이 다쳤을 때 형성된다; 상처 부위 근처의 표면에 있는 박테리아와 효모 복합체가 모여 주머니 모양의 움푹 패이기 시작한다. 이 주머니 속의 효모와 박테리아의 발효는 D. 메틀레리의 "신호" 역할을 하여 그 부지가 먹이로 이용될 수 있게 한다.[6]
D. 메틀러리의 숙주 종에 있는 효모 집단은 격리되어 숙주 선인장을 분해하는 역할을 하고 있다. 짝짓기를 하는 동안 파리의 양쪽 성별은 서로에게 효모와 박테리아를 준다. 그리고 나서 이 혼합물은 번식지에 놓이게 되고, 나중에 유충들이 그들의 건강을 증진시키는데 필요한 천연 꽃에 노출된다.[15]
짝짓기
구애행동
에레모필라 콤플렉스의 사막 드로소필라의 구애 행태는 비슷하다. 이 콤플렉스에 속하는 D. 메틀레리는 첫 번째 성간 접촉이 있는 순간 구애하기 시작한다. 일단 수컷이 암컷에게 빗질을 하면, 구애는 시작되고 수컷은 암컷을 따라오기 시작한다. 수컷은 날개 진동으로 구애 노래를 시작한다. 수컷은 암컷의 한 쪽을 따라가거나 암컷의 뒤를 따라가는 쪽을 택할 것이다. 수컷이 점령하기로 결정한 장소에 따라 수컷의 날개의 진동 주파수가 결정된다. 수컷이 암컷에게 멀리 있을수록 날개 펄스는 짧아진다. 암컷이 움직임을 멈추면 진동 주파수가 증가한다. 이 구애 노래 중에 수컷은 암컷의 성기를 핥는 것을 선택할 수도 있다. 만약 암컷이 수컷의 구애 노래 중에 수컷을 받아들이기로 선택한다면 암컷의 날개가 넓어지고 수컷이 수컷의 노래를 종료하게 된다. 파리들이 곧 교미하기 시작할 것이다.[16] 에레모필라 콤플렉스의 다른 종에 비해 D. 메틀러리의 수컷은 암컷과 교미를 강요할 가능성이 더 높다.
구애곡
D. mettleri의 구애곡은 뚜렷한 진동 펄스와 주파수가 특징이다. 암컷이 움직일수록 수컷의 진동 펄스는 불규칙해진다. 구애의 노래를 시작하려면 수컷이 날개를 78도 펼치며 몇 개의 짧은 펄스를 발산한다. 시작 과정이 끝난 후, 더 많은 규칙적인 펄스가 수컷에 의해 방출된다.[16]
남녀 조정기
D. 메틀러리에서 수컷이 짝에게 정자(즉, 사정)를 삽입한 후 질벽이 부풀어 오른다. 이것은 다른 수컷들이 암컷과 짝짓기를 하고 새끼를 낳는 것에 대한 방어 역할을 하는 결합 플러그 역할을 할 수 있다. 그리고 나서 수컷은 자신의 친자식을 보장받을 수 있게 된다. 이것은 D. 메틀러리의 수컷들이 각색한 것으로, 이 종의 암컷들이 여러 번 다시 번식하고 선택적으로 알을 수정하게 된다. 이 결합 플러그는 아버지의 친자 관계를 보장하는 것으로 밝혀졌지만, 이 플러그가 어떤 선택 목적에도 도움이 된다는 증거는 없다.
수정 과정이 일어난 후 6~8시간 후, 암컷은 짝에 의해 과잉 정자와 결합 플러그로 사용되었던 물질을 제거하게 된다. 이는 관계에 있는 남성과 여성 모두 이성의 성적 행동과 결과에 대한 통제력을 가지고 있음을 보여준다.[17]
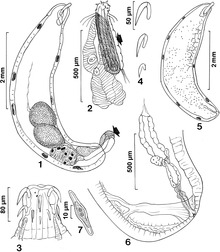
사정 소재
수컷의 사정 물질은 교배 플러그 역할을 하는 것보다 더 큰 목적을 제공한다. 이 액체는 영양소의 원천 역할을 할 수 있으며, 정자 활성화가 일어나는 데 걸리는 시간뿐만 아니라 난소 시간에도 영향을 미칠 수 있다. 이 액체에서 발견되는 영양소는 달걀의 크기를 증가시키고 암컷이 낳은 달걀의 양을 증가시키는 것으로 나타났다(액체의 영양소 밀도에 따라 다름). 따라서 이 액체의 질은 암컷의 생식 성공을 제한하거나 증가시킨다. 수컷이 짝짓기를 하는 동안 기여할 수 있는 액체의 양은 소노란 사막 드로소필라의 종에 따라 달라진다. D. 메틀러리에서 수컷은 짝짓기를 할 때 큰 사정투자를 하며, 따라서 수컷의 유체질도 유충의 발달에 큰 역할을 한다.
일부일처제에 있어서 사정 영양소의 역할
플러그의 효과로 인해 생식 파트너를 하나만 갖는 비용은 사정 물질에 의해 얻어지는 영양적 이점보다 더 커야 한다. 절충을 위한 비용은 자원 가용성에 따라 달라지며 썩은 물질의 분배와 성장에 의해 크게 규제된다. D. Mettleri에서는 남성들이 최상의 품질과 양의 액체를 기증하는 것에 대해 더 큰 생식 성공을 향한 진화적 추진이 있었다. 무선 라벨링은 여성의 난자 생산에 통합된 액체의 양을 측정하기 위해 사용되었다(6~8시간 후에는 몸에서 제거되지 않음). 그들은 가장 큰 질 부음을 가진 여성들이 더 작은 질 부음을 가진 여성들보다 단일 남성 파트너에서 나온 가장 많은 양의 정자를 포함한다는 것을 발견했다. 수정 과정을 겪고 있는 암컷을 갖지 않은 다른 디프테란들도 관찰되었다; 이들 암컷의 알에서 라디오 라벨이 부착된 물질은 발견되지 않았으며, D. 메틀러리와 다른 사막의 드로소필라의 수정 과정 특성이 남성 육아 보장에 필수적이라는 것을 보여주었다.[18]
모든 것이 일부일처제는 아니다.
2012년에 발표된 한 연구는 D. 메틀레리의 생식 생물학에 대한 광범위한 연구를 수행했으며, 정보의 생식 행동과 정보를 연관시켰다. 수컷 파리들은 암컷 파리들보다 더 빨리 성숙하지만, 이 연구는 이것이 왜 많은 야생 암컷들이 정자가 부족하거나 종양이 되면 너무 빨리 정자원을 착취하는 중요한 요소가 아니라는 것을 보여주었다. 연구진은 암컷에서 정자를 빠르게 순환시키는 것이 왜 이 종에서 연구실과 자연 모두에서 빈번하게 재생을 하는지를 설명해 준다고 결론지었다. 이 연구는 또한 다른 여성들보다 더 많이 재생된 여성의 수명에 어떠한 영향도 끼치지 않는다는 것을 발견했다. 사실, 암컷의 다중 교미에는 여성의 정자에 대한 더 큰 여성 후기 조절과 자손의 더 큰 성공이 있다.[19]
부모양육
난원
괴사체로서 이 파리들은 죽어가는 선인장의 썩어가는 쓰레기통에 알을 낳는 것으로 알려져 있다. 사막의 다른 괴사체에 비해 D. 메틀러리는 이러한 육즙이 함유된 선인장의 조직에서 번식하기보다는 이러한 썩어가는 선인장의 액체에 고착된 토양에서 번식하는 것을 선호한다. 이러한 행위의 의의는 사막의 건조한 상태가 선인장의 극심한 수분 손실을 초래하고, 그 결과 인근 토양으로 유입되는 액체 독소가 다른 과즙이 해독할 수 없는 정도로 집중된다는 것이다. 따라서 D. 메틸러리의 해독 능력은 다른 종에 의해 일반적으로 이용되지 않는 틈새 환경에서 번식할 수 있게 한다.[20]
숙주-식물 교대가 일반적인 숙주 선인장(세니타, 사구아로)과는 다른 화학적 구성을 가진 선인장을 착취하는 파리의 미래 자손에게 유전적 수준의 변화를 가져올 수 있는지 알아보기 위해 D. 메틀러의 전문 해독 능력을 시험했다.[21] 미래 세대는 숙주-작티(host-cacti)의 변종 화학적 환경을 해독하기 위해 더 잘 갖춰진 서로 다른 대사 경로를 촉발하는 유전자의 전사적 변화를 보여주었다.
적
기생충
족유전 혈통인 D. 메틀레리 등을 포함한 드로소필라 종은 사막 선인장의 괴사성 썩는 곳에 사는 외타라스틱 진드기에 의해 포식적인 압력을 보인다. D. 메틀레리가 이 지역에서 이런 로트나 품종을 먹이면, 이 진드기에 의해 기생하게 되는데, 이것은 그들의 수명을 줄이고 번식 능력을 방해한다.[22]
유전학

P450 유전자 계열과 CYP28A1 유전자의 상향조절
죽어가는 선인장의 썩어가는 액체 쓰레기통을 이용하기 위해 D. 메틀러리는 진화적 적응을 유도하는 돌연변이를 포함하고 있는 것으로 밝혀졌다. 이러한 돌연변이가 파리의 유전생물학적 저항의 원인으로 생각된다.[3] P450s, 에스테라제, 글루타티온 s-전달을 포함한 세 종류의 유전자는 곤충의 해독 능력을 조절하는 것으로 알려진 특정 유전자를 포함하고 있다.[23] CYP28A1 유전자는 메틀러리 혈통의 딥테란 계통에서 분리되어 이들 파리에서는 상향 조절된다. 이 유전자의 상향 조정은 숙주 공장에서 독성 알칼로이드에 대한 노출에 대한 적응적 반응이다.[24] 마찬가지로 Cyp 28A1 유전자 발현 역시 D. mettleri에서 조절되어 숙주식물의 알칼로이드에 더 많이 노출된다. P450 유전자의 집단은 거의 돌연변이를 일으키지 않고 염기서열 변화로 고도로 변형될 수 있는 단백질 구조를 가지고 있다. Cyp 28A1 유전자의 한 부위는 무극에서 극성 아미노산으로 단일한 변화를 가지고 있다. 이러한 변화는 물리적 단백질 구조에 중대한 변화를 야기하는 유전자 발현의 상향 조절을 유도하기에 충분하다; 그 결과는 실험된 D. 메틀러리 내 숙주 식물에서 독소의 해독을 증가시킨다.[3]
종간 유전적 고립
캘리포니아 해안의 카탈리나 제도에서 선인장충 디프테라(D. mettleri and D. mojavensis) 종 사이의 유전적 고립을 연구하는 새로운 모델이 나왔다. 이들 종의 파리들은 숙주식물을 이 섬들의 가시 돋친 배 선인장으로 바꾸었고, 이 파리들에서도 유전적 차이들이 고립되었다. 카탈리나 섬에서 이 종들의 유전자형을 가시 배 선인장을 숙주로 사용하는 다른 파리들과 비교한 결과 D. 메틀레리와 D. 모하벤시스 사이의 하플라타입 관계가 발견되었다. 결론적으로, 이 연구는 카탈리나 제도 사이에서 발견된 산페드로 해협의 지리적 장벽이 유전적 고립을 야기시켰고, 카탈리나 섬에서 가시 돋친 배를 먹고 사는 D. 메틀레리와 D. 모하벤시스 파리에 유전적 차이를 일으키는 생태학적 요인이 있다는 것을 발견했다.[25]
생리학
체온조절
드로소필라의 다른 종들은 다른 종류의 선인장을 이용할 것이다; D. 메틀러리가 규칙적으로 서식하는 숙주 종들은 지리적 분산에도 불구하고 이 파리 종의 열경화성 차이를 결정하는 것으로 나타났다.[26] 카돈이나 사구로 선인장과 같이 크기가 더 크고 더 많은 물을 저장하는 선인장의 종은 극심한 더위 기간 동안 더 차가운 내부 온도를 유지할 수 있다. 세니타와 같은 종은 더 작고 물을 덜 운반한다; 건조한 기후에서 더 높은 온도에 도달하기 쉽다.[11] D. 메틀레리는 사구아로와 세니타 선인장 근처에서 가장 흔하게 발견되는데, 다른 사막의 드로필라 종보다 열 내성 수준이 낮은 특징을 가지고 있다.[27] 숙주 선인장들 사이의 종 분포의 차이는 다양한 선인장 종에서 자연적으로 발견되는 독소의 해독 효율성에 기초하여 파리의 생리학적 차이와 관련이 있다.[28] D. mettleri는 사구로와 카돈 선인장의 더 시원하고 온화한 환경을 해독하는 데 필요한 전사적 변화를 포함한다. 시간이 지나면서 더 뜨거운 선인장에 살 필요가 줄어들어 내열성을 높이기 위해 유전자를 선택하게 되었다.[29]
상호주의
호스트 회전 플롯 공격
드로소필라 사막의 다른 종들은 사막 선인장 종인 사구로와 카르돈의 기판을 이용할 수 있다. D. 메틀레리는 세니타 선인장의 즙을 식품으로 사용할 수 있는 9종류의 2종 중 하나이다. 선인장의 알칼로이드는 대부분의 드로소필라와 애벌레의 발달에 매우 독성이 강하다. 사막 선인장의 독성으로 이어지는 성분으로는 중지방산 체인, 다이드록시 스테롤, 세니타 알칼로이드 등이 있다.[30] 드로소필라의 어떤 종들은 번식할 때 선인장 안에서 이러한 화학적 조건을 견딜 수 있지만 유충의 생존의 결과물이다. D. 메틀레리는 선인장(특히 세니타)의 독성이 유충과 번데기의 생존율을 떨어뜨리지 않는 두 종 중 하나로, 산모의 생존에는 거의 영향을 미치지 않는다.[31]
내포 지역 선택에서 상호주의 역할
사막 선인장의 괴사 조직과 썩어가는 선인장의 배설물로 촉촉한 토양 모두에서 성공적으로 번식할 수 있는 D. 메틀레리의 뚜렷한 능력에도 불구하고, 이 종은 조건이 더 시원하고 포식 압력이 덜한 토양 서식지에서 번식하는 것을 선호하는 행동적 선호를 가지고 있다.[32]
참조
- ^ a b c Castrezana S, Bono JM (2012). "Host plant adaptation in Drosophila mettleri populations". PLOS ONE. 7 (4): e34008. Bibcode:2012PLoSO...734008C. doi:10.1371/journal.pone.0034008. PMC 3320901. PMID 22493678.
- ^ Barker, J.S.; Starmer, William T.; Heed, W.B. (1982). "Origin of Drosophila in the Sonoran Desert.". Ecological genetics and evolution: the cactus-yeast-Drosophila model system.
- ^ a b c Bono JM, Matzkin LM, Castrezana S, Markow TA (July 2008). "Molecular evolution and population genetics of two Drosophila mettleri cytochrome P450 genes involved in host plant utilization". Molecular Ecology. 17 (13): 3211–21. doi:10.1111/j.1365-294X.2008.03823.x. PMC 2702709. PMID 18510584.
- ^ a b c d Heed WB (1977). "A new cactus feeding but soil breeding species of Drosophila (Diptera: Drosophilidae)". Proceedings of the Entomological Society of Washington. 79: 649–654.
- ^ Castrezana SJ, Markow TA (July 2008). "Sexual isolation and mating propensity among allopatric Drosophila mettleri populations". Behavior Genetics. 38 (4): 437–45. doi:10.1007/s10519-008-9209-0. PMID 18561017. S2CID 17423597.
- ^ a b c Breitmeyer CM, Markow TA (February 1998). "Resource availability and population size in cactophilic Drosophila". Functional Ecology. 12 (1): 14–21. doi:10.1046/j.1365-2435.1998.00152.x.
- ^ Gibbs AG, Perkins MC, Markow TA (July 2003). "No place to hide: microclimates of Sonoran Desert Drosophila". Journal of Thermal Biology. 28 (5): 353–62. doi:10.1016/S0306-4565(03)00011-1.
- ^ Stratman R, Markow TA (December 1998). "Resistance to thermal stress in desert Drosophila". Functional Ecology. 12 (6): 965–70. doi:10.1046/j.1365-2435.1998.00270.x.
- ^ Pfeiler E, Markow TA (May 2011). "Phylogeography of the Cactophilic Drosophila and Other Arthropods Associated with Cactus Necroses in the Sonoran Desert". Insects. 2 (2): 218–31. doi:10.3390/insects2020218. PMC 4553460. PMID 26467624.
- ^ Heed WB (1978). Brussard PF (ed.). "Ecology and Genetics of Sonoran Desert Drosophila". Ecological Genetics: The Interface. Proceedings in Life Sciences. New York: Springer: 109–126. doi:10.1007/978-1-4612-6330-2_6. ISBN 9781461263302.
- ^ a b Nobel P (1988). Environmental Biology of Agaves and Cacti. New York: Cambridge University Press.
- ^ Barker, JS; Starmer, William T.; Kircher, HW (1982). "Chemical composition of cacti and its relationship to Sonoran Desert Drosophila.". Ecological Genetics and Evolution: The Cactus–Yeast–Drosophila Model System. Sydney: Academic Press. pp. 143–158.
- ^ Biennial Report, National Institute of General Medical Sciences (U.S.). U.S. Department of Health and Human Services, Public Health Service, National Institute of Health. 1987.
- ^ Barker, JS; Starmer, William T. (1999). "Environmental effects and the genetics of oviposition site preference for natural yeast substrates in Drosophila buzzatii". Hereditas. 130 (2): 145–75. doi:10.1111/j.1601-5223.1999.00145.x. PMID 10479998.
- ^ Bakula M (November 1969). "The persistence of a microbial flora during postembryogenesis of Drosophila melanogaster". Journal of Invertebrate Pathology. 14 (3): 365–74. doi:10.1016/0022-2011(69)90163-3. PMID 4904970.
- ^ a b Alonso-Pimentel H, Spangler HG, Heed WB (January 1995). "Courtship sounds and behaviour of the two saguaro-breeding Drosophila and their relatives". Animal Behaviour. 50 (4): 1031–9. doi:10.1016/0003-3472(95)80103-0. S2CID 53176991.
- ^ Alonso-Pimentel H, Tolbert LP, Heed WB (March 1994). "Ultrastructural examination of the insemination reaction in Drosophila". Cell and Tissue Research. 275 (3): 467–79. doi:10.1007/BF00318816. PMID 8137397. S2CID 29013736.
- ^ Pitnick S, Markow TA, Riedy MF (May 1991). "Transfer of Ejaculate and Incorporation of Male-Derived Substances by Females in the Nannoptera Species Group (Diptera: Drosophilidae)". Evolution; International Journal of Organic Evolution. 45 (3): 774–780. doi:10.1111/j.1558-5646.1991.tb04347.x. PMID 28568832. S2CID 26489538.
- ^ Markow, Therese A.; Beall, Stephen; Castrezana, Sergio (2012-04-01). "The wild side of life". Fly. 6 (2): 98–101. doi:10.4161/fly.19552. ISSN 1933-6934. PMID 22627903.
- ^ Fogleman JC, Heed WB, Kircher HW (January 1982). "Drosophila mettleri and senita cactus alkaloids: fitness measurements and their ecological significance". Comparative Biochemistry and Physiology Part A: Physiology. 71 (3): 413–7. doi:10.1016/0300-9629(82)90427-3.
- ^ Hoang K, Matzkin LM, Bono JM (October 2015). "Transcriptional variation associated with cactus host plant adaptation in Drosophila mettleri populations". Molecular Ecology. 24 (20): 5186–99. doi:10.1111/mec.13388. PMID 26384860.
- ^ Perez-Leanos A, Loustalot-Laclette MR, Nazario-Yepiz N, Markow TA (January 2017). "Ectoparasitic mites and their Drosophila hosts". Fly. 11 (1): 10–18. doi:10.1080/19336934.2016.1222998. PMC 5354228. PMID 27540774.
- ^ Després L, David JP, Gallet C (June 2007). "The evolutionary ecology of insect resistance to plant chemicals". Trends in Ecology & Evolution. 22 (6): 298–307. doi:10.1016/j.tree.2007.02.010. PMID 17324485.
- ^ Danielson PB, Macintyre RJ, Fogleman JC (1995). "Isolation of novel cytochrome-P450 genes from alkaloid-tolerant cactophilic Drosophila and their phylogenetic relatives". Journal of Cellular Biochemistry: 196.
- ^ Richmond MP, Johnson S, Haselkorn TS, Lam M, Reed LK, Markow TA (December 2012). "Genetic differentiation of island populations: geographical barrier or a host switch?". Biological Journal of the Linnean Society. 108 (1): 68–78. doi:10.1111/j.1095-8312.2012.01986.x.
- ^ Morrison WW, Milkman R (May 1978). "Modification of heat resistance in Drosophila by selection". Nature. 273 (5657): 49–50. Bibcode:1978Natur.273...49M. doi:10.1038/273049b0. PMID 99662. S2CID 4162235.
- ^ Fogleman JC (1982). "Temperature effects in relation to the patterns of distribution and abundance of three species in the Drosophila affinis subgroup". Ecological Entomology. 12 (2): 139–148. doi:10.1111/j.1365-2311.1982.tb00652.x. S2CID 86542409.
- ^ Heed WB, Mangan RL (1986). "Community ecology of the Sonoran desert Drosophila.". In Ashburner M, Carson HL, Thompson Jr JN (eds.). The Genetics and Biology of Drosophila. London: Academic Press. pp. 311–345.
- ^ Hoffmann AA, Watson M (July 1993). "Geographical variation in the acclimation responses of Drosophila to temperature extremes" (PDF). The American Naturalist. 142 Suppl 1: S93–113. doi:10.1086/285525. PMID 19425954.
- ^ Fogleman JC, Kircher HW (January 1986). "Differential effects of fatty acid chain length on the viability of two species of cactophilic Drosophila". Comparative Biochemistry and Physiology Part A: Physiology. 83 (4): 761–4. doi:10.1016/0300-9629(86)90724-3.
- ^ Fogleman JC (1984). "The ability of cactophilic Drosophila to utilize soaked soil as larval substrates". Droso. Inf. Ser. 60: 105–7.
- ^ Meyer JM, Fogleman JC (November 1987). "Significance of saguaro cactus alkaloids in ecology of Drosophila mettleri, a soil-breeding, cactophilic drosophilid". Journal of Chemical Ecology. 13 (11): 2069–81. doi:10.1007/BF01012872. PMID 24301541. S2CID 28046204.