디프테라의 형태학
Morphology of Diptera디프테란 형태학은 곤충의 광범위한 형태학과는 몇 가지 중요한 면에서 다르다.디프테라는 매우 크고 다양한 목으로 대부분 작고 중간 크기의 곤충이다.그들은 이동식 머리에 눈에 띄는 겹눈과 복잡한 중흉부에 부착된 기능성 [1]막날개 한 쌍을 가지고 있습니다.두 번째 날개 쌍은 중흉부에 있으며, 반토막으로 감소합니다.이 목의 근본적인 특징은 날개 모양과 흉곽의 형태해부학적 적응에 있어 주목할 만한 전문화입니다. 즉, 비행 형태에 특별한 민첩성을 부여하는 특징입니다.필라형 더듬이, 양식 더듬이 또는 귀족 더듬이는 각각 [2]네마토세라, 브라키세라, 시클로라파 분류군과 관련이 있습니다.하위 분류군, 특히 속이나 종의 수준에서 상당한 형태학적 균일성을 보인다.날개 정맥과 마찬가지로, 관내 강모의 구성은 분류학에서 근본적으로 중요합니다.그것은 완전한 변성(알, 애벌레, 번데기, 성체)[2] 또는 홀로마볼라 [1]발생을 나타낸다.유충은 다리가 없고 선충에 하악골 입 부분이 있는 머리 캡슐을 가지고 있다.그러나 "높은 파리"의 애벌레는 머리가 없고 벌레처럼 생겼으며, 겨우 세 개의 등딱지벌레만 보인다.번데기는 선충류에서는 둔탁하고 [1][2]브라키세라에서는 공생한다.
어른
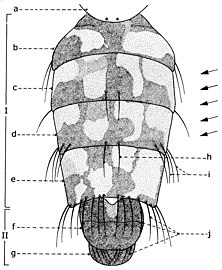
 | I: 머리, II: 흉부, III: 복부 1: 처방, 2: 전방 낙인, 3: 방추, 4: 기저골, 5: 발정골, 6: 흉골, 7: 후신경(추체), 8: ala, 9: 요석, 10: 반추, 11: 후위 낙인, 12: 대퇴골, 13: 정강이뼈, 14: 박차, 15: 발정골, 16: 전정골, 18: 전정골, 전정골: 전정골: 전정골, 6: 전정골: 전정골, 6: 전정골.: 상악 팔피; 26: 순음; 27: 순음; 28: 유사추; 29: 끝. |
어른은 작다(<2밀리미터(5µ64인치)에서 중형 곤충(-10밀리미터(25µ64인치) 미만).대형 디프테라는 희귀하며, 디프테라 미디다이와 판토프탈미다이의 특정 과만이 95-100밀리미터(3+3⁄4–4인치)의 날개폭에 이르며, 티푸라과의 열대종은 100밀리미터(4인치)가 넘는 것으로 기록되고 있다.칙칙하거나 밝은 색을 띠며 균일하거나 얼룩무늬가 있으며, 때로는 쥐과와 같이 흉내를 내기도 한다.분류학에서 가장 중요한 것은 부착된 관내 강모의 존재와 분포이다.
외부의
머리

머리는 흉곽과 구별되며, 목 부분이 눈에 띄게 좁아진다.하파리(Nematocera)에서는 예후(입이 앞쪽을 향하도록 머리를 수평으로 향하게 함)이고, 상파리(Brachycera)에서는 하후(head ognative)이다.두개골 캡슐의 모양도 다양합니다.선충에서, 머리 뒤쪽-복부 부분은 클라이푸스의 길이와 하위 속(하위 속)의 발달로 인해 눈에서 앞쪽으로 뻗어 있으며, 확장의 원위단은 '입 부분'이다.


"높은" 딥테라에서 헤드는 서브글로보스 형태를 가지며, 전두엽은 눈과 정점에 의해 상방으로 경계된 영역이다.Cyclorhapha Shuchophora에서 특히 중요한 형태학적 요소는 번데기에서 출현한 후 프틸리눔의 재흡착에 의해 형성된 프틸리날 봉합부의 존재이다.봉합사는 두 개의 영역을 분리한다. 1. 윗부분은 꼭대기, 안와부 및 제나와 연속성을 갖는 전두부, 2. 아랫부분인 얼굴 또는 클라이푸스는 더듬이의 삽입을 포함하고 윗입술로 이루어진 서간체 가장자리로 끝난다.
눈은 보통 매우 명백하지만, 브라키세라에서 주목할 만한 발달에 도달합니다.이 아목에서 눈은 눈에 띄게 볼록하고 머리 옆면 대부분을 차지하도록 성장했습니다.두 눈 사이의 공간은 때때로 후두부 앞쪽에서 이어지는 좁은 띠로 줄어들거나 눈 사이의 직접적인 접촉이나 가장자리로 인해 완전히 사라질 수 있습니다.겹눈의 형태학은 점액질에서 수천 개 정도의 상당한 수의 옴마티디아로 특징지어진다.주석은 있을 때 머리 꼭대기에 위치하며, 삼각형의 모서리에 배열되어 있으며, 스템매틱섬 또는 주상 삼각형이라고 불립니다.
계통학의 목적상, 머리털의 존재, 배치 및 배치는 중요하며 특정 용어를 가지고 있다.머리의 강모는: 때로는 하부 안와 강모라고 불리는 전두강모는 더듬이의 밑부분에서 정점으로 뻗어나가는 작은 골목과 비슷하고 중앙의 전두근을 가로로 엣지하는 전두강모이다.때때로 그것들은 더듬이 부착물 아래의 전경을 따라 더 낮거나 더 짧은 거리에 위치한다.안와 강모는 전두엽의 정점판에 위치하고 있으며 보통 상반부로 제한됩니다.이들은 안쪽 및 바깥쪽 안와부 강모라고 하는 세로줄로 배열될 수 있다.(전두엽의 정면 및 정점판은 전두엽과 안와엽의 배열에 기초하여 시각화할 수 있다.)주엽은 오셀리 사이의 정점에 위치한다.외측 및 내측 수직 강모는 정점과 후두엽 사이의 경계와 눈의 상부 모서리 근처에 위치한다.후수직 강모는s는 머리 중앙선 부근 후두부에 있는 오셀리 뒤에 위치한다.진동은 대개 구강 가장자리 부근의 원호 봉합사 안면부를 따라 작은 수로 배열된다.때로는 더 크거나 더 작은 거리에 걸쳐 봉합사를 따라 오르기도 하고 때로는 더듬이가 거의 붙어 있는 곳까지 오르기도 한다.가진동;구강 가장자리를 따라 이새낭을 배치한다.
때때로 용어가 모순될 수 있습니다.예를 들어, 아칼립트라과에는 보통 오셀라 삼각형의 뒤쪽 가장자리를 따라 다소 강한 두 개의 강모가 있습니다.이 털들은 19세기 이후 옛 문헌에서 "후수직 털"이라고 불리며, 최근 문헌에서는 가끔 이 용어가 사용된다.Steyskal(1976)은 Norticic Diptera 설명서(McAlpine, J.F., 1981년)와 Palaearic Diptera 설명서(Bernhard Merz, Jean-Paul Haenni, 2000년)에서 채택된 용어인 "포스트 해양 브리스털"을 제안했다.일부 아칼립트라과에서만 볼 수 있는 두 개의 다른 털은 주상 삼각형의 뒤쪽과 측면에 위치해 있고, 옛 문헌에서는 "내부 후두부"라고 불립니다.Steyskal(1976)은 "수직 강모"라는 이름을 사용하며 인용된 두 설명서의 기본 명명법에 동일한 이름이 사용됩니다.러시아어로 앞부분의 옆부분을 '오르빗'이라고 한다.영어에서 이 부분을 가장 일반적으로 '전두엽', '전두엽', 또는 '전두엽'이라고 부르는 반면, 단순 용어인 '오빗'은 복안경의 경계를 의미한다.아래쪽 머리 또는 얼굴의 중앙 부분은 종종 아래쪽 모서리에 '비브리새'라고 불리는 한 쌍의 큰 세타(또는 몇 쌍의)가 있고, 때로는 비브리새에서 위로 뻗은 능선을 따라 여러 개 또는 심지어 완전한 연속이 있다.후자의 세태는 러시아어 문헌에 얼굴털이 아닌 바이브리살 털이라고도 불린다.
더듬이는 두 아목의 구별의 기초가 되는 두 가지 기본적인 형태학적 유형으로 나뉜다.모든 파리 더듬이는 세 부분으로 구성되어 있다: 스케이프, 페디셀, 편모.내재된 근육은 경치와 [3]족저에서만 발견된다.네마토세라에서는 7~15개의 미분화 품목으로 이루어진 다립성, 실과 같은 깃털형이다.브라키세라에서 더듬이는 최대 6개의 세그먼트로 구성되어 있으며, 그 중 처음 3개는 잘 발달되어 있습니다.대부분의 Brachycera 계열에서 세 번째 세그먼트는 확대되고 끝이 뾰족한 세그먼트는 부속물로 축소됩니다. 즉, 단단할 때는 스타일러스, 강모처럼 생겼을 때는 아리스타라고 불립니다.
곤충의 입 부분은 조직적인 그룹에 따르면 다양한 형태를 보여준다.구강은 변형되어 흡인 주둥이로 결합되어 구조변화가 매우 심하다.조상들의 상태는 천공과 흡인형 주둥이로, 더 변형된 주둥이는 다양한 산줄기 또는 스폰지 액체를 형성합니다.음순은 그러한 스폰지 기관 중 하나이다.어떤 종들은 기능하지 않는 성체의 입 부분을 가지고 있다.



흉곽
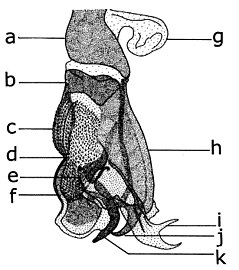
| 점액상 디페테란의 흉곽 형태학 오른쪽 측면(왼쪽), 등쪽(오른쪽) | |
 |  |
| 1: 중완골, 2: 상완골, 3: 중완골, 4: 중완골, 5: 후완골, 6: 중완골, 7: 중완골, 8: 중완골, 9: 중완골, 10: 날개 기반, 11: 중완골, 후완골, 12: 중완골, 중완골, 중완골, 중완골, 중완골, 중완골, 중완골, 중완골, 중완골, 중완골, 중완골, 중완골, 중완골, 중완골, 중완골, 중완골, 중완골, 중완골, 중완골, 중우튀르; 19: 중편모충 또는 아네피머론; 20: 복부 중편모충 또는 케이트피스테노; 21: 봉합 에피테노-프레콕살레; 22: 프로콕사; 23: 흉막 봉합; 24: 등편모충 또는 에피테노; 25: 프로플류론. 후두근(후두근): a: 3행시; dc: dorsocentral; ph: 사후; om: 상완골; ps: 전후두근; ia: 후두근(후두근), sa: 상완골(후두근), pa: 우편; pct: 후두근. | |
흉곽에 있는 분류학적으로 중요한 털

- 스쿠텀의 중앙 세로 축에 인접한 (a) 강모.불규칙하거나 두 개 이상의 행에 정렬할 수 있습니다.행 수, 각 행의 세트 수, 크기 및 두께가 유의합니다.많은 그룹에서, 삼행시 세태는 세툴레나 털로 대체된다.
- 다른 삼행시보다 발달된 두 개의 삼행시 강모(psc)가 방광 봉합사 앞에 삽입된다.
- dorsocentral(dc) 이러한 강모는 2개의 열을 따라 3행과 바깥쪽에 정렬된다.
- 사후(ph) 강모는 경막 전 부위에 정렬되고 상완골에서 방추골을 분리하는 봉합사와 평행합니다.
- 상완골(hm)은 상완골(hm)에 있다.
- 수술 전(ps)
- 노토플론 위의 노토플(np) 강모
- (ia) 도르소센트럴 시리즈 근처에 일정하게 정렬된 강모.위치가 잘 정의되어 있지 않다.
- 상완골(supral, 관능)(sa) 이러한 세태는 수에 제한이 있으며, 상완골 전 굳은살에서 상완골 영역까지 위치한다.
- 우편물(pa)의 수에 제한이 있으며, 날개 삽입 뒤쪽의 스쿠툼 측면 가장자리 근처의 우편물 굳은살 위에 위치한다.
- 스커텔럼에 있는 스커텔러(psct) 털.그것들은 가장자리이거나 스쿠텔룸의 등쪽에 있을 수 있습니다(디스크라고 불립니다).
흉막의 융기 또한 분류학적으로 중요하다.고려되는 문자는 유무, 수, 세태의 위치 및 머리카락 그룹입니다.
- 회충 또는 중두엽 – 회충 또는 중두엽 강모
- 강모 또는 흉골강모 – 강모 또는 흉골강모
- 프로에피넘 및 프로에피머론– 프로에피탈 및 프로에피메랄 강모 또는 프로페럴 강모
- 아네피메론(프테로플론)– 아네피메랄 또는 프테로플론 강모
- 메론(뇌중풍) – 메론 또는 뇌하수체 강모.
디프테라의 근본적인 특징은 날개의 모양과 흉곽의 형태 해부학적 적응으로 이루어진 놀라운 진화적 특성입니다.드물게 날개가 없는 형태를 제외하고, 디프테라는 보통 날개가 있고, 이동의 주요 수단으로 날개를 사용합니다.
날개
전문화 수준(해부학적, 기능적, 형태학적)은 일반적으로 이러한 곤충들이 날 수 있는 수준이며, 특히 민첩성을 언급할 때 예외적으로 잘 날 수 있습니다.모든 Diptera는 중흉부(전면)에 있는 한 쌍의 기능 날개만 갖추고 있습니다.중흉부에 있는 날개는 팔뚝 또는 로커 암으로 변환됩니다.이 특징에서 이 목의 이름은 "두 개의 날개"를 뜻하는 그리스어 딥테로스로부터 유래되었다.이 형태학적 구조의 결과 중흉부는 발달과 복잡성이 큰 세그먼트를 나타내며 전흉부와 중흉부는 현저하게 감소한다.
이 홀테어는 날아다니는 곤충의 균형을 잡기 위해 사용되는 곤봉 모양의 기관으로, 기계 감각 기관과 연결된 근위부로 구성되어 있습니다.날개와 구렁이 사이의 상동성은 초파리 드로소필라 멜라노가스터의 네 날개 돌연변이에 의해 증명된다.할테르의 발육은 조직군에 따라 다르다.할테라과는 얇지만 길고 또렷하게 보이지만 대부분의 다른 그룹에서는 보통 날개에 가려져 있다.가장 진보된 Diptera를 포함하는 Calyptratae에서, 할테르는 Calyptrae(할테어 위의 작은 막)에 의해 보호된다.
중흉부 날개는 완전히 막질이고, 완전히 투명하고 무색하거나, 인식에 유용한 영역 색소침착을 수반한다.그것의 표면은 세 개의 영역으로 나뉘어져 있다: 가장 발달된 것은 튼튼한 날개 정맥에 의해 지탱되는 경적(주 비행) 영역; 뒤쪽은 항문 영역; 그리고 마지막으로, 후방-근위부에는 확장된 소엽상 알룰라가 있다.겨드랑이 잎이라고도 불리는 알룰라는 날개 줄기 근위 후연에 있는 넓은 잎이다.그것은 상부 칼립터와 연속적이며, (통상적으로) 척골 절개라고 불리는 움푹 패인 곳에 의해 날개에서 분리된다.알룰리는 디프테라(Hennig, 1973년)의 새로운 특징이며, 알룰리는 보통 네마토세라(Anisopodidae 제외)에는 없거나 잘 발달하지 않지만 브라키세라에는 존재하며 비교적 크다.알룰라와 흉부 사이의 높은 Diptera는 상부 쇄골과 테굴라이다.칼립트라는 흉곽과의 날개 접합부 바로 아래에 있고 일부 디프테라 날개의 액와막의 일부입니다 – 두 개의 기본 엽은 칼립테레라고 불립니다.근위엽은 하부 칼립터라고 불립니다.그것은 좁은 막 인대로서 스쿠텔룸과 포스트노툼 사이의 고랑에서 발생하며, 상부 칼립터(또는 원위형 또는 비늘형 알라리)라고 불리는 더 원위엽이 그 위에서 급격히 접히는 끝부분(칼립트 주름)이다.상부의 칼립터는 보통 하부의 칼립터보다 크지만, 일부 그룹(Tabanidae, Acroceridae, 많은 Calyptratae)에서는 하부의 칼립터가 상부의 칼립터보다 크다.편두엽 가장자리는 각 편두엽의 뒤쪽 가장자리를 따라 난 털의 가장자리이다.테굴라(여기에 표시된 [1])는 늑연 하부에 있는 가장 가까운 판(늑골판 또는 견갑골판이라고도 함)이다.그 옆(멀리)은 기본형입니다.
정맥 체계는 단순하지만, 곤충의 날개 형태학의 용어를 정확하게 정의하기 위해 19세기 후반에 고안된 컴스톡-니덤 시스템을 대표한다.디프테라에는 코스타, 서브코스타, 레이디얼, 중앙, 큐비탈이 있다.게다가, 두 개의 항문 정맥이 있는데, 두 번째 정맥은 액와라고도 불리며, 항문 부위를 알룰라로부터 분리한다.날개 정맥, 가로 정맥 및 세포의 모양은 종 수준에서 분류군을 결정하는 데 중요한 특징이다.



분류학적으로 중요한 날개 정맥 용어
- 코스타(C), 늑하(Sc), 요골(R), 매체(M), 입방정맥(Cu), 항문정맥(A)이 그것이다.디프테라에서 늑골하부 뒤쪽 정맥의 동질성에 대해서는 많은 의견 차이가 있어 왔고, 일부 저자들은 단지 앞부분에서 뒤쪽으로 세로 정맥의 숫자를 세었을 뿐이다.종정맥 외에도 세 개의 주요 횡정맥이 있는데, 보통 크로스베인이라고 불립니다.이것들은 내측 또는 전측 크로스베인, 후측 또는 원반 크로스베인, 항문 크로스베인입니다.분류에 사용되는 날개의 주요 세포는 원반세포, 항문세포, 그리고 제2기세포이다.
Diptera 식별 키에서 가장 많이 사용되는 용어는 다음과 같습니다.–
- 셀컵후입방세포라고도 불리며 종종 항문세포라고도 불린다.셀컵의 형태는 중요한 특징입니다.
- 늑골 골절이것들은 특히 정신분열증에서 자주 발생한다.그것들은 코스타의 약점이고 그 수는 1에서 3까지이다.일반적으로 이러한 절단은 늑골하 삽입에 근접합니다.가장 빈번하게 발생하는 브레이크는 서브코스타가 코스타에 합류하는 지점 또는 완료 시 코스타에 합류하는 상상 지점입니다.거의 같은 빈도로 발생하는 유사한 균열은 상완골 크로스베인(크로스베인 h)과 약간 원위쪽에 위치한다.세 번째, 훨씬 덜 빈번한 파손은 코스타(세귀의 늑골)의 밑부분에서 짧고 굵고 강한 털이 있는 단면을 나타내는 크로스베인 h에 약간 가까운 곳에서 발견됩니다.이 브레이크는 늑골, 상완골, 늑골하 브레이크라고 불립니다.늑골 골절은 네마토세라와 브라키세라 하부에서 드물다.그것들은 비행하는 동안 날개를 구부리는 점으로 생각되며 번데기 날개의 주름과도 관련이 있다.
- 서브코스타두 번째 세로 날개 정맥, 코스타 뒤에 있습니다.코스타에 도달하거나 코스타에 도달하기 전에 사라지거나 R1에 합류할 수 있습니다.참고 항목 [2](= 많은 저자의 보조 정맥)
- 원반 세포날개 중앙의 닫힌 세포(d, dm)는 내측 정맥과 경계를 이루며 많은 네마토세라에서 크로스 베인 M-M에 의해 닫힙니다.정맥 CuA1에 둘러싸여 크로스 베인 DM-Cu에 의해 닫혀 있는 경우(대부분의 Lower Brachycera 및 모든 상위 Diptera와 같이), 엄밀하게 원반-중간세포(그러나 여전히 원반세포라고 불린다)
- 기저부 또는 첫 번째 기저부(br.이 세포는 요골의 기저부와 요골 섹터에 의해 전방으로 구분되며, 원위부는 요골-중간 크로스베인에 의해 구분됩니다.
- 기저 내측 또는 두 번째 기저(bm) 세포.이 세포는 후배지의 기저부에 의해 전방으로, 원위부에는 내측-경부 크로스베인에 의해 구분된다.
- 서브아피칼 셀정맥 R4+5, M1+2, rm과 꼭대기 날개 가장자리 사이의 세포.M1+2가 날개 가장자리에서 종료되면 아첨면 셀이 열립니다.M1+2가 R4+5를 향해 커브되어 날개 여백 전에 R4+5를 결합하면 셀이 닫힙니다.
- 항문 정맥 날개 끝에 도달하거나 도달하지 않는 하나 또는 두 개의 항문 정맥이 있을 수 있습니다.
스쿠텔룸은 거의 항상 뚜렷하지만 중턱 바로 뒤에 있는 것보다 훨씬 작습니다.매크로채테는 분류학에서 중요하다.
.svg)





비교적 얇은 다리는 치밀하게 배치된 강모를 가지고 있으며, 강모는 또한 카에토갤럭시에서도 기능합니다.대퇴골과 정강이뼈는 등, 흉골, 후흉골, 복부, 장골, 후퇴골의 강모의 조합을 가질 수 있다.이러한 털의 위치, 수, 크기 및 기울기는 고등 파리 분류에서 중요합니다.다리는 등쪽 복측면에서 구부러집니다(대퇴골의 경골).정강이뼈(특히)와 대퇴골의 배면은 종종 매우 작은 강모의 이중선으로 식별된다.또 다른 중요한 강모는 정강이뼈의 전정골이다(가족 차원에서 유무는 중요하다.
복부
복부의 형태학은 생식기능으로서 남녀 모두 형태소 원자적응에 의해 실질적으로 결정된다.일반적으로 10개의 소변기(복부 또는 복부 후 세그먼트 중 하나)는 첫 번째 소변기와 마지막 소변기의 구조변경에 의해 더 적은 수의 소변기로 감소한다.일반적으로 첫 번째 요산염의 위축과 2° 및 3° 요산염의 결합이 있다.터가이트와 흉골은 서로 잘 구별될 수 있지만 종종 터가이트가 흉골과 겹치는 차이가 있습니다. 극단적인 경우는 터가이트의 확장이 복부에서 합쳐져 튜브 구조나 고리를 형성하는 것입니다.암컷의 경우 마지막 소변이 얇아지고 늘어지면서 신축성 있는 신축성 있는 난소판이 형성됩니다.이러한 형태학적 적응은 종종 말기 8번째 요산의 경화작용을 동반한다. 따라서 난소기는 알과 유충을 수용하는 유기체의 조직을 통해 침투할 수 있다.수컷의 경우 마지막 소변은 생식기와 통합된 장치를 형성하기 위해 복잡한 변형을 겪습니다.구조 변화의 정도와 성질은 계통 그룹에 따라 다르지만, 보통 9번째 요로테르가이트의 엽이 겸자(에판듐)와 IX 요로스테르노(하이판듐)로 발달하는 것을 포함한다.때때로 복부의 축을 따라 뒤틀림이 있어 상완부와 하완부의 위치가 역전된다.
사코파가 카르나리아의 복부

칼리포라의 복부
아프리디갈리아 아드리안폰티 수컷 생식기



내부의
소화기 계통
포겟
전두엽의 P4부 수축은 전두엽 자체에 외생적인 세로토닌에 의해 조절되며, 대신 용혈에서 도달한다.특히 세로토닌은 수축률을 높인다.Liscia et al 2012는 이 세로토닌이 흉부-복부 신간절의 세로토닌 신경총에서 유래한 것일 수 있음을 입증했다.작물이 가득 차 세로토닌이 P4에 도달하면 P4가 수축하여 P2로 밀어넣고, 그 때 괄약근 활성화에 따라 P2가 중간관에 밀어넣어 소화를 시키거나 [4]공허로 역류시킨다.
유충
디프테라 유충의 대부분은 수생 환경, 부패하는 유기 기질, 그리고 다른 유기체(풍기, 동물, 식물)에서 삽니다.그러므로 그들의 형태학적 구조는 상당히 단순하다.
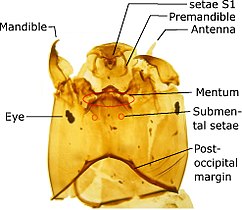
디프테라 유충은 (다리가 없는) 수생 유충이지만, 때때로 가성 유충과 유사한 부속지를 가지고 있다.머리에는 보통 눈이 없고, 씹는 입 부분, 최대 6개의 세그먼트가 있는 변형된 더듬이가 있으며, 어느 정도 발달하거나 유두 모양으로 축소됩니다.머리는 흉부(뇌수유충)와 명확하게 구별될 수 있고, 몸의 다른 부분과 구별되지 않거나(소두증), 또는 흉부에 움푹 패여 있을 수 있습니다(암호두증).

Molophilus, Tipulidae.지렁이 같은 애벌레는 앞다리나 부위가 없지만, 뒷부분에 있는 잎과 나선형(2개)을 통해 디프테라로 식별된다.

아노펠레스 유충(두유충)

츄렉스 레스튜언스 유충

애벌레과(소두증)
오소클라디아과 머리
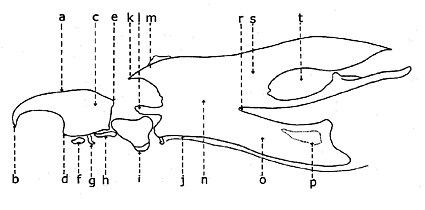
머리 인두 장치

호흡 사이펀을 가진 에리스탈리스 애벌레








기관나선의 수와 위치에 따라 다음과 같은 종류의 호흡장치를 구별할 수 있다.
- 스티그마(영혼)가 없는 무신경성
- 성흔이 한 쌍밖에 없는 메타뉴스트입니다.이건 복부에 있어요.
- 두 쌍의 스티그마가 있는 양친매제입니다전흉부에 한 쌍씩, 복부에 한 쌍씩.
- 흉부 스티그마 두 쌍과 복부 여덟 쌍이 있는 완전 신경성.
브라키세라에서 발견되는 가장 흔한 유형은 양지신생인 반면, 다른 종류는 주로 수생 애벌레에서 나타난다.Brachycera Cyclorrhapha의 애벌레는 지렁이처럼 생겼으며, 일반적으로 벌레라고 불리울 정도로 신체 부위(머리, 흉부, 복부)가 거의 구별되지 않습니다.이 유충들은 원통형 원추형이고 복부가 넓어요다른 씹는 입과는 달리 복잡한 두인두 장치에 형성되는 갈고리 모양의 2개의 턱과 일련의 내두경화물로 대표되는 간이 입부는 수직면을 따른 이동을 갖춘다.
특별한 형태학적 적응은 수중 환경 또는 내생충으로서 적응된 애벌레에서 관찰된다: 예를 들어, 긴 호흡 사이펀을 가진 Eristalis의 부생성 수생 애벌레는 슬러시 또는 부패한 물에서 살 수 있는 반면, Tachinidae의 애벌레는 기관으로 이끄는 호흡관을 가지고 있다.호스트 또는 호스트 몸 밖으로 이동합니다.
번데기
Diptera 번데기는 둔부, 엑사레이트 또는 코아크테이트일 수 있다.둔갑 번데기는 날개와 다리의 윤곽을 볼 수 있지만 전체가 하나의 큐티클에 의해 싸여 있기 때문에 몸의 다른 부분과 가깝게 눌려 있다. 엑스트라 번데기는 자신의 큐티클에 둘러싸인 부속지를 가지고 있기 때문에 곤충의 나머지 부분과 분리할 수 있다.유충의 피부 안에서 코아크테이트 번데기가 발달한다.
시클로라파 번데기 아목 브라키세라 번데기는 번데기(유충 피부의 경화에 의해 형성되는 경우)에 마지막 유충 단계의 형태학적 및 생화학적 추출물의 수정에 의해 형성되는 코크타이트 번데기를 가지고 있다.번데기의 개구부가 성충이 되었을 때 두 개의 큰 조직군인 아스키자와 시조포라를 구별하는 방식입니다.
오뚝이 번데기는 일반적으로 자유롭고 보호되지 않는다. 단, 비단 파편과 함께 만들어진 보졸레티에 의해 보호된다.
그들의 삶의 마지막 단계에서, 디프테라의 번데기는 움직일 수 있게 된다.


참고 자료 및 추가 자료
이 기사는 주로 Diptera의 이탈리아어 페이지 번역에 기초하고 있다.
- Brown, B.V., Borkent, A., Cumming, J.M., Wood, D.M., Woodley, N.E. 및 Zumbado, M. (편집자) 2009 중앙아메리카 딥테라 매뉴얼제1권 NRC 연구 프레스, 오타와 ISBN978-0-660-19833-0
- Capinera, John L. (편집), 2008 Entomology Springer Verlag 백과사전.ISBN 978-1-4020-6242-1.
- 콜리스, D.H. & McAlpine, D.K. 1991 Diptera(파리), 페이지 717-786.인: 곤충학 부문.호주의 곤충 캔버라(스폰서)의 영연방과학산업연구기구(Commonwealth Science and Industrial Research Organization.멜버른 대학교프레스, 멜버른
- 그리피스, G.C.D.디프테라 사이클로라하파의 계통학적 분류로, 특히 수컷 포스트아브도멘의 구조에 관한 것이다.경, 제8호, 340쪽 [닥터 W. 정크, 노스 V. 헤이그](1972년)
- 빌리 헤닉 다이 라벤포르멘 데 디프테렌 3세테일, 아카드- 베를린, 발락.185 페이지, 3 페이지, 1948년
- McAlpine, David K., 1958 Acalptrate Diptera (Incecta) 호주 가족의 핵심 호주 박물관 24(12) 183-190 PDF 전문 및 수치
- McAlpine, J.F. 1981 형태학 및 용어집 입력: McAlpine, J.P. 등 (ed.) : Nortic Diptera vol.1 Ottawa:캐나다 농업 연구소, 모노그래프 27.ISBN 0660107317 PDF 다운로드 매뉴얼
- Bernhard Merz, Jean-Paul Haenni, 2000 성인 Diptera의 형태학 및 용어(말기아 제외).인: Laszlo Papp, Béla Darvas가 Palaic Diptera 매뉴얼에 기여.제1권: 일반 및 응용 딥테롤로지.부다페스트, 사이언스 헤럴드, 2000: 22-51.ISBN 963-04-8839-6.
- K. G. V. Smith, 1989 영국 파리들의 미성숙 단계에 대한 소개.알, 번데기, 번데기에 대한 기록이 있는 쌍꺼풀 애벌레.영국 곤충 식별을 위한 핸드북 Vol 10. pdf 다운로드 매뉴얼 (2부 주요 텍스트 및 그림 색인)
- Hongfu, Zhu, 1949 미성숙한 곤충을 아는 방법; H. F. Chu에 의한 수집, 사육, 연구 제안과 함께 많은 미성숙한 곤충의 목과 과를 식별하는 삽화 키.W.C. Brown Co., 아이오와주, 두부케의 주요 자연 시리즈 사진.온라인 전문은 이쪽
레퍼런스
- ^ a b c Capinera, John L. (2008). Encyclopedia of Entomology (2 ed.). Springer Science & Business Media. p. 1481. ISBN 9781402062421.
- ^ a b c Meyer, John R. (28 March 2016). "Diptera: True Flies / Mosquitoes / Gnats / Midges". General Entomology. NC State University. Retrieved 5 February 2018.
- ^ Dessi, Giancarlo (28 May 2019). "Morphology and anatomy of adults: antennae". Flies [Insecta: Diptera]. Retrieved 27 August 2020.
- ^ Stoffolano, John G.; Haselton, Aaron T. (2013-01-07). "The Adult Dipteran Crop: A Unique and Overlooked Organ". Annual Review of Entomology. Annual Reviews. 58 (1): 205–225. doi:10.1146/annurev-ento-120811-153653. ISSN 0066-4170.
외부 링크
![]() 위키미디어 커먼스의 디프테라 해부학 관련 매체
위키미디어 커먼스의 디프테라 해부학 관련 매체
- 파리 해부학.
- 흰꼬리원숭이과 용어집
- 지안카를로 데시
- 드로잉 윙 벤션
- 몽골의 수생 디프테라
- 호주 MDFRC 수생 딥테라
- La Monde des Penges 삽화 기사 (프랑스어)



{kind=link}