동물의 의사소통
Animal communication_great-egret-8243-web_(324180866).jpg)
동물의 커뮤니케이션이란,[1] 수신자의 현재 또는 장래의 행동에 영향을 주는, 1개 또는 복수의 동물(송신자 또는 송신자)로부터, 1개 또는 복수의 다른 동물(수신자 또는 수신자)에게 정보를 전달하는 것입니다.정보는 구애 표시와 같이 의도적으로 전송될 수도 있고, 포식자에서 먹이로 냄새를 전달하는 것처럼 의도하지 않게 전송될 수도 있습니다.정보는 여러 [2]수신자의 "청취자"에게 전송될 수 있습니다.동물의 의사소통은 동물 행동, 사회학, 신경학, 동물 인지학을 포함한 분야에서 빠르게 성장하고 있는 연구 분야이다.상징적인 이름 사용, 감정 표현, 학습 그리고 성적 행동과 같은 동물 행동의 많은 측면들이 새로운 방식으로 이해되고 있다.
송신측으로부터의 정보가 수신측의 동작을 변경하는 경우, 그 정보를 「신호」라고 부릅니다.신호 전달 이론은 신호가 모집단에서 유지되기 위해, 송신자와 수신자 모두 보통 상호작용으로부터 어느 정도 혜택을 받아야 한다고 예측합니다.송신자에 의한 신호 생성과 수신자의 인식 및 후속 응답은 함께 [3]진화하는 것으로 생각됩니다.신호는 시각과 청각과 같은 여러 메커니즘을 수반하는 경우가 많으며, 신호가 송신자와 수신자 모두의 조정된 동작을 이해하려면 신중한 연구가 필요하다.
★★★

★★★
- 제스처:대부분의 동물들은 눈에 띄는 신체 부위나 신체 움직임을 시각적으로 보여줌으로써 의사소통을 이해한다.동물들은 특정 정보를 전달하기 위해 신체 부위를 드러내거나 강조합니다.어미 청어갈매기는 먹이를 가지고 둥지로 돌아왔을 때 자신의 밝은 노란색 부리를 병아리 옆에 바닥에 드러냅니다.병아리들은 청어갈매기의 부리 아랫턱에 있는 빨간 점을 두드리며 구걸하는 반응을 보입니다.이 신호는 부모가 음식을 역류하도록 자극하고 먹이 신호를 완성시킨다.이 의사소통에서 두드러지는 특징적인 형태학적 특징은 어미의 붉은 반점 부리인 반면, 땅을 향해 두드리는 것은 붉은 반점을 병아리가 볼 수 있게 하여 독특한 [4]움직임을 보여준다.Frans de Waal은 언어가 몸짓에 의해 진화했는지 이해하기 위해 보노보와 침팬지를 연구했다.그는 유인원과 인간 모두 의사 소통을 [5]위해 의도적인 제스처만을 사용한다는 것을 발견했다.
- 표정:동물의 의사소통에서 감정의 또 다른 중요한 신호는 얼굴 제스처이다.Blue와 Yellow Macaws는 그들이 친숙한 동물보호사와의 상호작용에 어떻게 반응하는지 이해하기 위해 연구되었다.연구에 따르면 Blue Macaws와 Yellow Macaws는 [6]관리인과 상호 교류하는 동안 자주 얼굴이 붉어지는 것을 보여주었다.또 다른 실험에서, 제프리 모길은 증가하는 통증에 대한 반응으로 생쥐의 얼굴 표정을 연구했다.그는 쥐들이 5가지 인식 가능한 얼굴 표정을 보인다는 것을 발견했다: 안와 조임, 코와 뺨의 팽만함, 그리고 귀와 수염의 [7]이동 변화.
- 시선 추적:인간이나 비인간이나 사회적 동물들은 다른 [8]포유동물들의 머리와 눈의 방향을 관찰함으로써 시선을 쫓는 것을 의사소통의 한 형태로 사용한다.유인원, 원숭이, 개, 새, 늑대, 거북이에 대한 연구는 두 가지 다른 작업에 초점을 맞췄습니다: "멀리 있는 공간을 향해 다른 사람의 시선을 따라가기"와 "예를 들어 시야를 [9]가리는 장벽에 직면했을 때 신호를 따르도록 그들 자신을 재배치함으로써 시각 장벽 주위에 기하학적으로 다른 사람의 시선을 따라가기"입니다.다양한 종류의 동물들이 후자를 나타내는 것으로 입증되었지만, 오직 유인원, 개, 늑대, 그리고 까마귀들만이 먼 우주로 다른 사람들의 시선을 따라갈 수 있었다.마모셋과 따오기는 "추종하는 기하학적 시선"을 보여줄 수 없었다.연구자들은 아직 따라오는 시선의 인지적 기초에 대한 명확한 그림을 가지고 있지 않지만, 발전적 증거는 따라오는 "단순한" 시선과 따라오는 "기하학적" 시선이 아마도 다른 인지 [8]메커니즘에 의존한다는 것을 보여준다.
- 색상 변경:색상의 변화는 성장과 발달 중에 일어나는 변화와 기분, 사회적 맥락, 온도와 같은 비생물적 요인에 의해 유발되는 변화로 나눌 수 있다.후자는 많은 분류군에서 볼 수 있다.문어와 갑오징어와 같은 일부 두족류는 그들의 [10]피부의 겉으로 보이는 색깔, 불투명도, 반사율을 바꿀 수 있는 특별한 피부 세포를 가지고 있습니다.위장용으로 사용할 뿐만 아니라, 사냥이나 구애 [11]의식에서도 피부색의 급격한 변화가 사용됩니다.오징어는 몸의 반대쪽에서 완전히 다른 두 개의 신호를 동시에 표시할 수 있다.수컷 오징어가 수컷이 있는 곳에서 암컷에게 구애할 때 수컷은 암컷을 향하고 암컷은 다른 [12]수컷을 속이기 위해 암컷은 외면하는 패턴을 보인다.일부 컬러 신호는 사이클로 발생합니다.예를 들어, 암컷 올리브 개코원숭이가 배란을 시작하면, 그녀의 선천적인 부위가 부풀어 오르고 밝은 빨간색/분홍색으로 변합니다.이것은 수컷들에게 그녀가 [13]짝짓기를 할 준비가 되었음을 알려준다.훔볼트 오징어는 생물발광성이기 때문에 어두운 바다 [14]환경에서 시각적으로 의사소통을 할 수 있습니다.
- 생물 발광 통신:빛의 생성에 의한 의사소통은 바다의 척추동물과 무척추동물, 특히 깊은 곳(예: 아귀 물고기)에서 흔히 발생한다.두 가지 잘 알려진 형태의 육상 생물발광은 반딧불이와 지렁이에게서 발생한다.다른 곤충들, 곤충 애벌레들, 고리형 동물들, 거미류 그리고 심지어 균류 종들도 생물 발광 능력을 가지고 있다.어떤 생물발광 동물들은 스스로 빛을 내는 반면, 다른 동물들은 생물발광 [15]박테리아와 공생 관계를 가지고 있다.동물들은 먹이를 유인하거나, 짝을 유인하거나, 잠재적인 [16]포식자들로부터 자신들을 보호하기 위해 생물 발광 빛을 보인다.(「」도 참조해 주세요.생물 발광 생물 목록)
청각


많은 동물들이 발성을 통해 의사소통을 한다.목소리 소통은 짝짓기 의식, 경고음, 음식 공급원의 위치 전달, 사회적 학습 등 많은 목적을 가지고 있습니다.많은 종에서 수컷은 짝짓기 의식 중에 다른 수컷과의 경쟁의 한 형태로서 암컷에게 신호를 보내기 위해 울부짖는다.개구리, 망치머리 박쥐, 붉은 사슴, 혹등고래, 코끼리 물개, 그리고 지저귀는 새를 예로 [18][19][20]들 수 있습니다.음성 의사소통의 다른 예로는 캠벨 [21]원숭이의 경보음, 긴팔원숭이의 영역음, 그리고 그룹을 [22]구별하기 위해 큰 창코박쥐의 주파수 사용이 있습니다.버벳 원숭이는 4개의 다른 포식자들에게 각각 뚜렷한 경보를 울리고, 다른 원숭이들의 반응은 그 울음소리에 따라 적절히 달라진다.예를 들어, "독수리" 알람은 원숭이들이 [23]땅 위의 은신처를 찾게 하는 반면, 만약 뱀에게 경보가 울리면, 원숭이들은 나무에 올라갑니다.프레리독은 또한 포식자의 차이를 알리는 복잡한 울음소리를 사용한다.Con Slobodchikoff와 다른 사람들에 따르면, 프레리독은 다가오는 [24][25][26][27]포식자의 종류, 크기, 속도를 전달한다.고래의 발성은 [28][29]지역에 따라 다른 방언을 가지고 있는 것으로 밝혀졌다.
모든 동물이 청각적 의사소통의 수단으로 발성을 사용하는 것은 아니다.많은 절지동물들은 소리를 내기 위해 특수 신체 부위를 서로 비벼댄다.이것을 스트리디테이션이라고 합니다.귀뚜라미와 메뚜기는 이로 잘 알려져 있지만, 갑각류, 거미, 전갈, 말벌, 개미, 딱정벌레, 나비, 나방, 밀리페데스, 지네를 포함한 많은 다른 동물들도 스트리디션을 사용한다.청각적 의사소통의 또 다른 수단은 가시가 있는 물고기의 수영 방광의 진동이다.수영 방광과 부착된 음파 근육의 구조는 가시가 많은 어족에 따라 매우 다양하며, 그 [30]결과 다양한 소리를 냅니다.신체부위를 함께 타격하는 것도 청각신호를 발생시킬 수 있다.이에 대한 잘 알려진 예로는 방울뱀의 꼬리 끝 진동이 경고 신호입니다.다른 예로는 새의 부리 소리, 매너킨 구애 전시의 날개 박수, [31]고릴라의 가슴 박치기 등이 있다.

굴을 파고드는 동물들은 위협을 전달하기 위해 휘파람을 부는 것으로 알려져 있고, 때로는 기분을 느낍니다.고산 마못을 포함한 마못 종과 같은 종들은 이러한 특성을 보여줍니다.휘파람은 프레리독과 같은 동물들이 위협을 전달하기 위해 사용하는데, 프레리독은 동물 왕국에서 가장 복잡한 의사소통 시스템을 가지고 있습니다.프레리 도그는 동물의 속도, 모양, 크기, 종, 그리고 인간 고유의 복장, 그리고 인간이 [32]총을 소지하고 있는지 여부를 전달할 수 있다.이 의사소통 방법은 보통 보초병을 두 발로 세워 잠재적인 위협을 조사하는 방식으로 이루어지며, 나머지 무리는 음식을 찾는다.위협이 확인되면 보초병은 호루라기 경보를 울리고 (때로는 위협을 묘사하기도 한다) 이때 무리는 그들의 굴로 후퇴한다.위협의 강도는 보통 보초가 휘파람을 부는 시간에 따라 결정됩니다.보초병은 무리 전체가 안전한 곳으로 갈 때까지 계속해서 경보기를 울리고, 거기서 보초가 [33]굴로 돌아간다.
후각

가장 오래된 통신 방법임에도 불구하고, 화학 통신은 부분적으로 우리 환경에 화학 물질이 풍부하고 샘플 [31]내의 모든 화학 물질을 감지하고 측정하는 것이 어렵기 때문에 가장 잘 알려지지 않은 형태 중 하나입니다.환경 속의 화학 물질을 탐지하는 능력은 많은 기능을 하는데, 중요한 기능은 음식을 탐지하는 것인데,[31] 이것은 지구 생명체의 초기 시기에 바다에 사는 단세포 생물에서 처음 생겨난 기능이다.이 기능이 진화함에 따라, 유기체는 자원, 동종(동종, 즉 짝과 친족), 이종(다른 종, 즉 경쟁자와 포식자)[31]에서 나오는 화학 화합물을 구별하기 시작했다.
예를 들어, 작은 민어는 북방 [34]갯장어와 같은 포식자와 관련된 화학적 신호 농도를 감지하여 서식지를 피하는 것이 좋습니다.육식동물이 눈에 띄기 전에 육식동물의 존재를 인지한 후 적응행동(숨는 등)으로 대응하는 능력을 가진 소형동물은 살아남아 [35]번식할 가능성이 높아집니다.대서양 연어는 포식자의 신호를 감지하는 것 보다 한 걸음 더 나아가서, 포식자에 의해 개체가 피해를 입었을 때,[36] 동종에게 화학 신호를 방출합니다.다른 종에서도 관찰된 바와 같이 산성화와 pH의 변화는 물리적으로 이러한 화학적 신호를 교란시키고, 이는 동물의 [36][37]행동에 다양한 영향을 미친다.
후각 표시와 후각 문지르는 [38][39]것은 포유류에서 흔한 후각 소통의 형태이다.동물에 의해 문지르는 냄새의 예는 곰에게서 볼 수 있는데, 곰은 영역을 표시하거나 다른 사람들에게 그들이 그곳에 있다는 것을 알리고 [40]멀리하기 위한 방법으로 이것을 한다.
전기
전기 통신은 동물에게는 드문 형태의 통신이다.그것은 주로 수생 동물에서 볼 수 있지만, 일부 육상 포유류, 특히 오리너구리와 에키드나는 의사소통에 사용될 [41]수 있는 전기장을 감지합니다.
약한 전기 물고기는 전기 위치와 함께 전기 통신의 예를 제공합니다.이 물고기들은 전기 기관을 사용하여 전기장을 발생시키고, 전기장은 전기 수용체에 의해 감지된다.파형의 차이와 필드의 변화 빈도는 종, 성별, 정체성에 대한 정보를 전달한다.이러한 전기 신호는 호르몬, 일주기 리듬, 그리고 다른 물고기와의 상호작용에 반응하여 생성될 수 있습니다.그들은 또한 사회적 [42]질서를 가진 종들 사이에서 사회적 위계질서를 조정하는 역할을 할 수 있다.상어나 가오리와 같은 일부 포식자들은 수동적인 전기 [43]수신을 통해 이러한 전기 유전적인 물고기들을 엿들을 수 있다.
만지다
- 터치 메커니즘에 대한 자세한 내용은 Somatosensory 시스템 및 기계 수용기를 참조하십시오.
접촉은 많은 사회적 상호작용에서 중요한 요소이다.다음은 몇 가지 예입니다.
- 화이팅:싸움에서 터치는 상대에게 도전하고 싸우는 동안 움직임을 조정하기 위해 사용될 수 있습니다.패자가 투항을 [44]나타내는 데 사용할 수도 있다.
- 짝짓기: 포유류는 종종 몸치장을 하거나, 쓰다듬거나, 서로 문지르면서 짝짓기를 시작합니다.이를 통해 화학적 신호를 적용하고 잠재적 배우자에 의해 배출된 신호를 평가할 수 있는 기회를 제공합니다.터치는 수컷 캥거루가 암컷의 꼬리를 잡는 것처럼 수컷이 암컷에 올라타려는 의도를 알릴 수도 있다.짝짓기 동안 접촉 자극은 쌍 위치 결정, 조정 및 생식기 [45]자극에 중요합니다.
- 소셜 통합:터치는 사회적 통합을 위해 널리 쓰이고 있는데, 이는 동물의 사회적 손질에 의해 특징지어지는 것이다.사회적 손질은 여러 가지 기능을 가지고 있습니다; 그것은 손질된 동물에서 기생충과 잔해들을 제거하고, 동물들 사이의 사회적 유대감이나 위계적 관계를 재확인하고, 손질된 개인에 대한 후각적 신호들을 검사할 수 있는 기회를 줍니다, 아마도 추가될 수 있습니다.이러한 행동은 사회적 곤충, 조류 및 [46]포유류에서 관찰되었습니다.
- 먹이찾기: 어떤 개미 종들은 먼저 더듬이와 앞다리로 먹이를 두드리며 새로운 먹이를 찾기 위해 동료 일원을 모집합니다. 그리고 나서 신체적인 접촉을 유지하면서 그들을 먹이 공급원으로 인도합니다."순찰자"는 둥지를 떠나 근처에 위험이 있는지 확인하고 신체 [47]접촉을 통해 "식량자"를 모집한다.이것의 또 다른 예는 꿀벌의 [31]흔들림 춤이다.
- 옹기종기: 장시간 신체 접촉 또는 옹기종기 하는 것도 사회적 통합에 도움이 됩니다.허들링은 후각 [48]또는 촉각 정보의 전달과 함께 열 교환을 촉진합니다.어떤 유기체들은 군체 안에서 끊임없이 접촉하며 산다. 예를 들어 군체 산호와 같다.이러한 방식으로 개체들이 밀접하게 연결되어 있을 때, 전체 군락은 소수의 [49]개체들에 의한 혐오 또는 경보 움직임에 반응할 수 있습니다.몇몇 초식성 곤충의 님프와 애벌레에서는 장기간 접촉하는 집단이 그룹 협동에 중요한 역할을 합니다.이러한 집계는 행렬 또는 로제트의 [50]형태를 취할 수 있습니다.
지진
지진 통신은 토양, 물, 거미줄, 식물 줄기 또는 풀잎과 같은 기질을 통해 전달되는 자기 발생 진동 신호를 사용하여 정보를 교환하는 것입니다.이러한 형태의 통신에는 몇 가지 장점이 있습니다. 예를 들어 빛과 소음 수준에 관계없이 전송될 수 있으며, 일반적으로 짧은 범위와 짧은 지속성을 가지고 있어 포식자에 의한 탐지 위험을 줄일 수 있습니다.지진 통신의 사용은 개구리, 캥거루 쥐, 두더지 쥐, 벌, 선충 등을 포함한 많은 분류군에서 발견된다.네발동물은 보통 수신기의 [51]천골에 의해 감지되는 신호인 신체 부분으로 땅을 두드려서 지진파를 만든다.천골은 내이에 있는 기관으로 균형을 잡기 위해 사용되는 막낭을 포함하고 있지만, 이러한 형태의 통신을 사용하는 동물에게서도 지진파를 감지할 수 있다.진동은 다른 종류의 [52]통신과 결합될 수 있습니다.
온도

많은 다른 뱀들이 적외선(IR) 열방사를 감지할 수 있는 능력을 가지고 있는데, 이것은 이 파충류들이 5에서 30 μm 사이의 파장에서 포식자나 먹이가 방출하는 복사열로부터 열방사를 얻을 수 있게 해줍니다.이러한 감각의 정확성은 맹목적인 방울뱀이 먹이 [53]동물의 취약한 신체 부위를 공격 대상으로 삼을 수 있다는 것이다.이전에는 구덩이의 장기가 주로 먹이 탐지기로 진화했다고 생각되었지만, 이제는 체온을 [54]조절하는 데 사용될 수도 있다고 믿어진다.
체온조절을 가능하게 하는 얼굴의 구멍은 피트바이퍼와 일부 보아와 버마에서 평행한 진화를 거쳤으며, 피트바이퍼에서 한 번, 보아와 [55]버마에서 여러 번 진화했다.구조의 전기생리학은 혈통 간 유사하지만 전체 구조 해부학에서는 다릅니다.가장 표면적으로 볼 때, 핏바이퍼는 눈과 콧구멍 사이에 머리 양쪽에 하나의 큰 핏 기관을 가지고 있는 반면, 보아와 비단뱀은 비늘 안이나 사이에 윗입술과 때로는 아랫입술 사이에 세 개 이상의 비교적 작은 핏을 가지고 있다.피트바이퍼들은 단순한 피트 구조와 달리 현탁된 감각막을 가지고 있어 더 발달되어 있다.독수리과에서 독수리속은 독수리과(Crotalinae)에서만 볼 수 있다.IR 방사선의 검출에도 불구하고, 광수용체의 IR 메커니즘은 광화학 반응을 통해 빛을 검출하는 반면, 뱀의 얼굴 구덩이에 있는 단백질은 온도에 민감한 이온 채널입니다.빛에 [56]대한 화학적 반응보다는 피트 기관의 난방을 수반하는 메커니즘을 통해 적외선 신호를 감지합니다.이는 유입되는 적외선 방사선이 주어진 이온 채널을 빠르고 정확하게 가열하고 신경 자극을 트리거할 수 있는 얇은 피트막과 일치하며, 피트막을 혈관화하여 이온 채널을 원래의 "휴식" 또는 "비활성" [56]온도로 빠르게 냉각할 수 있습니다.
일반적인 뱀파이어 박쥐(Desmodus rotundus)는 코 잎에 [57]특수 적외선 센서를 가지고 있습니다.흡혈박쥐는 오로지 피를 먹고 사는 유일한 포유동물이다.데스모두스는 적외선 감지 기능을 통해 소와 말과 같은 동질적인 동물을 약 10~15cm 범위 내에서 위치시킬 수 있습니다.이 적외선 인식은 표적 먹잇감에서 최대 혈류 영역을 감지하는 데 사용될 수 있습니다.
자동 통신
자동 통신은, 송신자와 수신자가 같은 개인인 통신의 일종입니다.송신자는 환경에 의해 변경된 신호를 송신하고, 최종적으로 같은 개인에 의해서 수신됩니다.변경된 신호는 먹이, 포식자 또는 동종을 나타낼 수 있는 정보를 제공합니다.송신기와 수신기는 같은 동물이기 때문에 선택압력은 신호효율을 최대화한다.즉, 전파왜곡과 노이즈에도 불구하고 방사신호가 수신기에 의해 정확하게 식별되는 정도를 최대화한다.태평양 청어와 같은 몇몇 종들은 포식자들로부터 이러한 메시지를 가로채도록 진화했습니다.그들은 그것을 조기 경고 신호로 사용할 수 있고 [58]방어적으로 대응할 수 있다.자동 통신에는 두 가지 유형이 있습니다.첫 번째는 활성 전장입니다. 유기체는 전기 기관을 통해 전기 펄스를 방출하고 물체의 투영된 기하학적 특성을 감지합니다.이것은 전기 물고기 [59]Gymnotiformes와 Mormyridae에서 발견됩니다.두 번째 유형의 자동 통신은 박쥐와 이빨고래에서 발견되는 반향 위치 확인입니다.반향 위치 확인은 소리를 내고 [60]물체에서 돌아오는 진동을 해석하는 것을 포함한다.박쥐에서, 반향 정위는 또한 그들의 환경을 지도화하는 목적도 가지고 있다.그들은 가시광선 없이 이전에 있었던 공간을 인식할 수 있다. 왜냐하면 그들은 반향 [61]위치로부터 얻은 피드백의 패턴을 기억할 수 있기 때문이다.
기능들
동물의 의사소통에는 많은 기능이 있다.하지만, 어떤 것들은 다른 것들보다 더 자세히 연구되었다.여기에는 다음이 포함됩니다.
- 대회 중 커뮤니케이션:동물의 의사소통은 자원을 둘러싼 경쟁의 승자를 결정하는데 중요한 역할을 한다.많은 종들은 공격성이나 공격 의지를 나타내는 뚜렷한 신호나 먹이, 영토 또는 [62]짝을 둘러싼 경쟁 중에 후퇴를 전달하는 신호를 가지고 있다.
- 짝짓기 의식:동물들은 가능한 짝의 관심을 끌거나 짝을 이루는 결합을 굳히기 위해 신호를 만든다.이러한 신호에는 신체 부위나 자세의 표시가 수반되는 경우가 많습니다.예를 들어, 가젤은 짝짓기를 시작하기 위해 특징적인 자세를 취합니다.짝짓기 신호에는 후각 신호 또는 종 고유의 짝짓기 신호도 포함될 수 있습니다.지속적인 쌍결합을 형성하는 동물들은 종종 서로 대칭적인 표시를 한다.유명한 예로는 줄리안 헉슬리가 연구한 큰 볏이 있는 큰 멧돼지의 상호 전시, 둥지에서 많은 종의 거위들과 펭귄들이 보여주는 승리 전시, 그리고 [citation needed]낙원의 새들이 보여주는 화려한 구애 전시 등이 있다.포유류의 "번식 울음소리"는 암컷의 번식 상태를 나타내거나 다른 [63]짝을 유혹할 수 있다.
- 소유권/지역:영토, 음식 또는 짝을 요구하거나 방어하는 데 사용되는 신호입니다.일부다처 도마뱀(Anolis carolinensis)은 영역이나 짝을 방어할 때 암컷 간보다 수컷 간 거리가 더 먼 곳에서 공격적 징후를 보입니다.수컷은 [64]암컷에 비해 생식력이 높아 서로 거리를 두도록 진화한 것으로 알려져 있다.
- 리다이렉트된 공격:자극을 두려워하는 몇몇 동물들은 그들 [65]근처에 있는 누군가를 공격할 것이다.
- 식품 관련 신호:많은 동물들은 짝짓기, 자손, 또는 사회적 집단의 다른 구성원을 먹이 공급원으로 끌어들이기 위해 "음식 호출"을 한다.아마도 가장 정교한 음식과 관련된 신호는 Karl von Frisch에 의해 연구된 꿀벌의 Waggle 춤일 것이다.움켜쥐거나 새끼를 낳아 구걸하는 것으로 잘 알려진 한 예는 고산성 지저귀는 새이다.어린 까마귀는 새로운 음식이나 테스트되지 않은 음식을 만나면 나이든 까마귀에게 신호를 보냅니다.레서스 마카크들은 벌을 피하기 위해 다른 원숭이들에게 먹이 공급원을 알리기 위해 먹이 전화를 보낼 것이다.페로몬은 사회의 다른 구성원들을 식량 공급원으로 이끌기 위해 많은 사회적 곤충들에 의해 방출된다.예를 들어, 개미는 땅 위에 페로몬 흔적을 남기는데, 다른 개미들이 그들을 식량 공급원으로 인도하기 위해 따라갈 수 있다.
- 경보 호출: 경보 호출은 포식자의 위협을 전달합니다.이것은 사회 집단의 모든 구성원(때로는 다른 종)이 그에 따라 반응할 수 있도록 합니다.공격 [66]위험을 줄이기 위해 은신처로 도망치거나, 움직이지 못하게 하거나, 그룹으로 모이는 것을 포함할 수 있습니다.알람 신호가 항상 발성이 되는 것은 아닙니다.뭉개미는 더 많은 개미를 유인하기 위해 경보 페로몬을 방출하고 공격 상태로 [67]보낸다.
- 메타 통신:후속 신호의 의미를 변경하는 신호.한 가지 예는 개들의 '놀이 얼굴'인데, 이것은 이후의 공격적인 신호가 심각한 공격적 에피소드가 아닌 놀이 싸움의 일부라는 것을 나타냅니다.
동물 행동 해석
위에서 설명한 것처럼, 많은 동물들의 몸짓, 자세, 그리고 소리는 근처의 동물들에게 의미를 전달한다.이러한 신호는 해석보다 설명하기가 더 쉬운 경우가 많습니다.특히 길들여진 동물들과 유인원들에게 의인화하는 것은 유혹적이지만, 이것은 꽤 오해를 불러일으킬 수 있다; 예를 들어, 원숭이의 "미소"는 종종 공격성의 표시이다.또, 같은 제스처는, 발생하는 문맥에 따라서 다른 의미를 가질 수 있습니다.예를 들어, 집에서 기르는 개의 꼬리 흔들기와 자세는 1872년에 출판된 찰스 다윈의 "인간과 동물의 감정 표현"에서 설명되었듯이 많은 의미를 전달하기 위해 다양한 방법으로 사용될 수 있다.다윈의 삽화 중 일부는 여기에 복제되어 있다.
- 개의 다른 감정을 나타내는 꼬리 위치의 예

"테이블 위에서 고양이를 보고 있는 작은 개"

"다른 개에게 적대적인 의도로 접근하는 개"
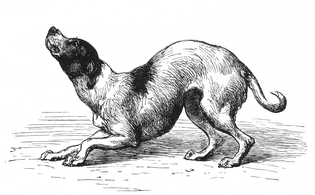
'겸손하고 애정 어린 강아지'
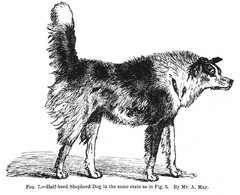
"이복종 목동 개"

"개가 주인을 어루만지다"





특정 간 통신


많은 동물들의 의사소통은 종내적인 것이다. 즉, 그것은 같은 종의 구성원들 사이에서 일어난다.종간 통신에 관해서는 포식자와 먹잇감 사이의 통신에 특히 관심이 있습니다.
포식자의 먹잇감
포식자가 탐지할 수 있는 방식으로 먹잇감이 움직이거나 소리나 진동을 일으키거나 냄새를 풍기면 포식자와 [68]통신하는 것이다.이것은, 상기의 「커뮤니케이션」의 정의와 일치합니다.이런 유형의 통신은 포식자가 동종 메시지를 가로채는 경우 가로채기 도청이라고 합니다.
하지만, 먹잇감의 몇몇 행동들은 분명히 실제 또는 잠재적인 포식자들에게 향합니다.좋은 예는 경고 색칠이다: 잠재적 포식자에게 해를 끼칠 수 있는 말벌과 같은 종들은 종종 밝은 색을 띠고, 이것은 본능적으로 또는 경험의 결과로 그러한 동물을 공격하는 것을 피할 수 있는 포식자의 행동을 변화시킨다.어떤 형태의 모방은 같은 범주에 속합니다: 예를 들어, 호버파리는 말벌과 같은 색을 띠고, 비록 그들이 침을 쏠 수는 없지만, 포식자에 의한 말벌의 강한 회피는 호버파리를 어느 정도 보호합니다.경고 색상과 유사한 방식으로 작용하는 행동 변화도 있습니다.예를 들어, 늑대와 코요테와 같은 송곳니는 필요하다면 싸울 것임을 나타내기 위해 이빨을 드러낸 채 으르렁거리는 것과 같은 공격적인 자세를 취할 수 있고 방울뱀은 그들의 잘 알려진 덜컹거림을 잠재적 포식자들에게 경고하기 위해 사용합니다.때로는 행동 변화와 경고 색채가 결합되기도 하는데, 밝은 색상의 배를 제외하고 몸의 대부분이 주변환경과 조화를 이루도록 색을 입힌 특정 양서류 종에서 그러하다.잠재적인 위협에 직면했을 때, 그들은 어떤 식으로든 독이 있다는 것을 나타내며 배를 드러낸다.
포식자 통신의 또 다른 먹잇감의 예는 추적-분리 신호이다.추적-감시 신호는 먹잇감이 포식자에게 신호를 보내는 사람이 탈출할 준비가 되어 있기 때문에 추격은 이득이 되지 않을 것이라고 지시할 때 발생합니다.추적 분리 신호는 시그널러와 수신자 모두에게 이점을 제공합니다.이러한 신호는 송신자가 도망치는 시간과 에너지를 낭비하는 것을 방지하고 수신자가 포착될 가능성이 낮은 비용이 많이 드는 추적에 투자하는 것을 방지합니다.먹잇감의 탈출 능력을 광고하거나 표현형 상태를 반영하거나(품질 광고), 먹잇감이 포식자를 탐지했다고 광고(퍼셉션 광고)[68]할 수 있다.추적 세부 신호는 물고기(Godin and Davis, 1995), 도마뱀(Cooper 등, 2004), 유제류(Caro, 1995), 토끼(Hally 1993), 영장류(Zuberbuhler 등 1997), 설치류(Shelley 및 Blumstein 2005, Clark, 2005, 2005), 버드류(alvarez)를 포함한 다양한 분류군에 대해 보고되었다.품질 광고 추적-디테런트 신호의 친숙한 예는 포식자가 있는 상태에서 톰슨의 가젤과 같은 일부 영양에 의해 동시에 점프를 하는 동안 뻣뻣한 다리로 달리는 것의 뚜렷한 조합인 스토팅(때로는 발음이라고도 함)입니다.스토팅에 대한 최소 11개의 가설이 제안되었습니다.오늘날 가장 유력한 이론은 포식자들에게 놀라움의 요소가 사라졌다는 것을 경고한다는 것입니다.치타와 같은 포식자들은 영양이 사냥할 때 추격전이 거의 성공적이지 않다는 사실로 증명된 기습 공격에 의존합니다.포식자들은 실패할 가능성이 높은 추적에 에너지를 낭비하지 않습니다(최적의 먹이찾기 행동).품질 광고는 비주얼 이외의 모드로 전달할 수 있습니다.깃발꼬리캥거루쥐는 여러 가지 다른 맥락으로 여러 가지 복잡한 발버둥 패턴을 만들어 내는데, 그 중 하나는 뱀과 마주쳤을 때입니다.발을 구르는 것은 근처의 자손들에게 경고를 줄 수 있지만, 아마도 쥐가 성공적인 공격을 하기에는 너무 경각심을 가지고 있기 때문에 뱀의 포식자 [69]추격을 막을 수 있다는 진동을 지면을 통해 전달했을 것이다.

일반적으로 포식자들은 먹이와의 통신을 줄이려고 합니다. 이렇게 하면 사냥의 효과가 떨어지기 때문입니다.하지만, 어떤 형태의 포식자는 의사소통을 위해 먹잇감의 행동을 바꾸고 포획을 쉽게 하는 방식으로 발생합니다. 즉, 포식자에 의한 속임수입니다.잘 알려진 예로는 낚시꾼 물고기가 있는데, 낚시꾼은 먹잇감이 찾아오기를 기다리는 매복 포식자이다.그것은 이마에서 튀어나온 다육질의 생물 발광 성장을 가지고 있으며 턱 앞에 매달려 있다.작은 물고기들은 미끼를 잡으려고 하고, 낚시꾼 물고기들이 그들을 잡을 수 있는 더 나은 위치에 놓인다.기만적인 의사소통의 또 다른 예는 점핑거미속(Myrmarachne)에서 관찰된다.이 거미들은 더듬이를 흉내내기 위해 앞다리를 공중으로 흔드는 방식 때문에 흔히 "모방 거미"라고 불립니다.
★★★★★★★★★★★★★★▼
인간이 동물의 행동을 해석하거나 명령을 내리는 다양한 방법은 이종 간 의사소통의 정의와 일치한다.동물의 의사소통에 대한 능숙한 해석은 인간에 의해 보살펴지거나 훈련되는 동물의 복지에 매우 중요할 수 있다.예를 들어 고통을 나타내는 행동을 인식할 필요가 있다.실제로, 예를 들어, 인간이 임박한 공격의 신호를 인식하지 못한다면, 동물과 그 관리인의 생존이 위태로울 수 있다.또한 인간이 아닌 동물 종들이 인간의 신호를 인간 자신과는 다르게 해석할 수도 있다는 것을 고려하는 것이 중요하다.예를 들어, 포인팅 명령어는 [70]도그에서 객체가 아닌 위치를 참조합니다.
90년대 후반부터, 숀 세네찰이라는 과학자는 개와 말에서 학습된 가시적이고 표현력이 풍부한 언어를 개발하고, 연구하고, 사용해 왔다.이 동물들에게 몸짓과 같은 미국 수화를 가르침으로써,[71] 동물들은 그들이 필요로 하는 것을 얻기 위해 새로운 수화를 스스로 사용하는 것으로 밝혀졌다.동물 언어에 대한 최근의 실험은 자연 동물과의 의사소통에 대한 관계는 불확실하지만, 인간과 동물의 의사소통을 확립하기 위한 가장 정교한 시도일 것이다.
★★
의사소통의 중요성은 일부 동물이 이를 촉진하기 위해 진화한 매우 정교한 형태학, 행동 및 생리학에서 명백하다.이것들은 공작의 꼬리, 숫사슴의 뿔, 그리고 프릴목 도마뱀의 프릴과 같은 동물 왕국에서 가장 눈에 띄는 구조물들을 포함하지만, 유럽 청어 갈매기의 부리에 있는 약간의 붉은 반점까지도 포함합니다.두루미 춤, 갑오징어의 패턴 변화, 바우어새에 의한 재료의 수집과 배열 등 매우 정교한 행동이 의사소통을 위해 진화했다.동물에서 의사소통의 중요성에 대한 다른 증거는 이 기능에 대한 생리학적 특징의 우선순위 부여이다.예를 들어, 새소리는 전적으로 그것의 생산에 전념하는 뇌 구조를 가지고 있는 것으로 보인다.이 모든 적응은 진화적 설명을 필요로 한다.
설명에는 두 .
- 된 이 그것을할 수 하는 것.
- 동물이 의사소통을 촉진하고, 의사소통을 방출하며, 이에 반응하는 구조를 개발하도록 적응시키는 선택적 압력을 식별한다.
이러한 문제들 중 첫 번째에 대한 중요한 공헌은 콘라드 로렌츠와 다른 초기 동물행동학자들에 의해 이루어졌다.그룹 내 관련 종을 비교함으로써, 그들은 원시 형태에서 의사소통 기능이 없었던 움직임과 신체 부위가 한쪽 또는 양쪽 파트너에게 의사소통이 기능하는 맥락에서 "포착"될 수 있고, 보다 정교하고 전문화된 형태로 진화할 수 있다는 것을 보여주었다.예를 들어, Desmond Morris는 풀 핀치에 대한 연구에서 다양한 종에서 부리를 씻는 반응이 일어나 프리닝 기능을 하지만, 일부 종에서는 이것이 구애 [72]신호로 정교하게 만들어졌다는 것을 보여주었다.
두 번째 문제는 더 논란이 많았다.초기 동물행동학자들은 의사소통이 종 전체의 이익을 위해 일어났다고 가정했지만, 이것은 성적으로 번식하는 동물의 진화에서 수학적으로 불가능하다고 여겨지는 집단 선택의 과정을 필요로 할 것이다.무관한 집단에 대한 이타주의는 과학계에서 널리 받아들여지지 않고 오히려 집단생활의 이점인 다른 사람들로부터 같은 행동을 기대하는 상호 이타주의로 볼 수 있다.사회생물학자들은 전체 동물 집단에 혜택을 주는 행동은 개인에게만 작용하는 선택 압력의 결과로 나타날 수 있다고 주장했다.유전자 중심의 진화 관점은 개체나 종 전체에 미치는 영향이 [73]해롭더라도 개체군 내에서 유전자를 더 넓게 확립할 수 있도록 하는 행동이 긍정적으로 선택될 것이라고 제안한다.

의사소통의 경우, 존 크렙스와 리처드 도킨스의 중요한 논의는 개인의 선택 하에 나타나는 경보 호출과 구애 신호와 같이 겉으로 보기에 이타적이거나 상호주의적인 의사소통의 진화에 대한 가설을 확립했다.이것에 의해, 커뮤니케이션이 항상 「정직」한 것은 아닐지도 모른다는 것을 깨닫게 되었습니다(실제로, 모방과 같이, 그렇지 않은 몇개의 명백한 예가 있습니다).특히 아모츠 자하비가 장기적으로 존재할 수 없다고 주장하면서, 진화적으로 안정된 부정직한 의사소통의 가능성은 많은 논란의 대상이 되어 왔다.사회생물학자들은 또한 공작의 꼬리와 같은 명백한 과도한 신호 구조의 진화에 대해 염려해 왔습니다; 이것들은 성적 선택의 결과로 나타날 수 있을 뿐이며, 이것은 co에서 장점을 부여하는 특징의 빠른 과장으로 이어질 수 있는 긍정적인 피드백 과정을 만들 수 있다고 널리 생각됩니다.짝짓기 경쟁 상황
공작의 꼬리와 같은 특성의 진화를 설명하는 한 가지 이론은 '도망 선택'이다.이것은 두 가지 특성을 필요로 합니다: 밝은 꼬리와 같은 특성, 그리고 그 특성을 위해 선택하기 위해 암컷에 존재하는 기존의 편견.암컷들은 더 정교한 꼬리를 선호하기 때문에 수컷들은 성공적으로 짝짓기를 할 수 있다.여성의 심리를 이용해 양성 피드백 루프를 만들어 꼬리가 커지고 밝아진다.결국, 진화는 진정될 것입니다. 왜냐하면 수컷에 대한 생존 비용은 그 특성이 더 [74]이상 정교해지는 것을 허용하지 않기 때문입니다.폭주 선택을 설명하기 위해 두 가지 이론이 존재합니다.첫번째는 좋은 유전자 가설이다.이 이론은 정교한 전시가 건강의 정직한 신호이며 정말로 더 나은 짝이라고 말한다.두 번째는 핸디캡 가설입니다.이것은 공작의 꼬리가 핸디캡으로 유지하는데 에너지를 필요로 하고 포식자들에게 더 잘 보이게 한다는 것을 설명한다.따라서, 신호는 유지 비용이 많이 들고 시그널러의 상태를 나타내는 정직한 지표로 남아 있습니다.또 다른 가정은 신호가 고품질 남성이 생산하는 것보다 저품질 남성이 생성하는 데 비용이 더 많이 든다는 것이다.이는 단순히 고품질 남성이 비용이 많이 드는 [3]시그널링에 할당할 수 있는 에너지 비축량이 더 많기 때문입니다.
인지적 측면
동물행동학자와 사회생물학자들은 관련 동물들이 그들이 방출하고 받는 신호의 의미를 이해하는지에 대한 의문을 제기하지 않고 자극에 대한 다소 자동적인 반응의 관점에서 동물의 의사소통을 특징적으로 분석했다.그것은 동물 인식의 핵심 질문이다.보다 고도의 이해를 필요로 하는 신호 시스템이 몇 가지 있습니다.많이 논의된 예는 버벳 원숭이의 알람 호출 사용이다.Robert Seyfarth와 Dorothy Cheney는 이 동물들이 서로 다른 포식자(leopard, 독수리, 뱀)가 있을 때 서로 다른 경보음을 내고, 소리를 들은 원숭이들은 적절하게 반응한다는 것을 보여주었지만, 이 능력은 시간이 지남에 따라 발전하고, 소리를 내는 개인의 경험도 고려한다는 것을 보여주었다.위에서 논의한 메타커뮤니케이션 또한 보다 정교한 인지 과정을 필요로 하는 것으로 보인다.
병코돌고래는 호루라기의 특성을 제거하더라도 호루라기로부터 신원 정보를 인식할 수 있다고 보고되어 왔다. 돌고래는 발신자의 목소리나 위치로부터 독립적으로 신원 정보를 전달하는 것으로 보여진 사람이 아닌 유일한 동물이다.이 문서의 결론은 다음과 같습니다.
시그니처 휘파람 모양이 음성 특징과는 무관한 식별 정보를 가지고 있다는 사실은 이러한 휘파람을 사람의 이름을 사용하는 것과 유사하게 개인을 지칭하거나 참조하는 참조 신호로 사용할 수 있는 가능성을 제시합니다.병코돌고래의 인지 능력, 발성 학습과 복제 기술, 그리고 핵분열-융합 사회 구조를 고려할 때, 이 가능성은 더 많은 조사를 필요로 하는 흥미로운 것이다.
--
인간의 행동
또 다른 논란의 여지가 있는 문제는 인간의 행동이 동물의 의사소통과 비슷한 정도인지, 또는 우리의 언어 능력의 결과로 그러한 모든 의사소통이 사라졌는지 여부이다.우리의 신체적 특징들 중 일부는 눈동자, 수염, 콧수염, 깊은 성인 남성의 목소리, 아마도 여성의 가슴은 신호를 생성하는 것에 대한 적응과 매우 유사합니다.Irenaeus Eibl-Eibesfeldt와 같은 동물행동학자들은 미소짓기, 얼굴 찌푸리기, 그리고 인사에 눈썹을 깜박이는 것과 같은 얼굴 제스처는 다른 영장류에서 대응하는 신호와 관련이 있을 수 있는 보편적인 인간의 의사소통 신호라고 주장해왔다.얼마나 최근에 구어가 생겨났는지를 고려할 때, 인간의 바디 랭귀지는 우리가 가지고 [76]있는 의사소통과 비슷한 기원을 가진 다소 무의식적인 반응을 포함할 가능성이 매우 높다.
인간은 또한 동물들과 상호작용하기 위해 종종 동물들의 의사소통 신호를 모방하려고 한다.예를 들어, 고양이는 천천히 눈을 감는 가벼운 제휴 반응을 보인다; 인간은 종종 관용적인 관계를 확립하기 위해 애완 고양이를 향해 이 신호를 흉내낸다.애완 동물을 쓰다듬고 쓰다듬고 문지르는 것은 아마도 그들의 고유 고유 의사소통 패턴을 통해 작용하는 행동일 것이다.
개는 인간의 의사소통을 이해하는 능력을 보여 왔다.객체 선택 작업에서 개는 숨겨진 음식과 [77]장난감을 찾기 위해 손가락과 시선 방향과 같은 인간의 의사소통 제스처를 활용합니다.하지만, 인간이 가리키는 것과는 대조적으로, 개는 방향이나 [78]위치를 가리키기 때문에 다른 의미를 가지고 있다.또한 개들은 사람의 얼굴을 볼 때 왼쪽 시선이 치우치는 것으로 나타나 그들이 인간의 [79]감정을 읽을 수 있다는 것을 보여주었다.개들은 다른 개들과 함께 시선 방향을 이용하거나 왼쪽 시선의 편견을 보이지 않는다.
21세기의 동물 의사소통 분야에서의 새로운 접근법은 응용 행동 분석, 특히 기능적 의사소통 훈련을 사용한다.이러한 형태의 훈련은 이전에 자폐증을 가진 아이들과 같이 특별한 요구를 가진 사람들이 언어를 발달시키는 것을 돕기 위해 학교와 클리닉에서 사용되어 왔다.AnimalSign Center의 Sean Senechal은 2004년부터 개, 말 등 가축화된 동물과의 기능적 의사소통 훈련과 유사한 접근방식을 사용해 동물과 사람에게 고무적인 결과와 혜택을 주고 있다.동물을 위한 기능적 의사소통 훈련인 Senechal은 "동물 수화"라고 부릅니다.여기에는 제스처(간소화된 미국 수화 등), Picture Exchange Communication System, 탭 및 발성을 통한 커뮤니케이션 교육이 포함됩니다.동물을 위한 과정은 단순화되고 수정된 [citation needed]기술을 포함한다.
| ★★★★★★ | |
|---|---|
언어학에서 동물 의사소통 시스템의 관심은 인간의 언어와의 유사성과 차이점에 있다.
- 인간의 언어는 이중 발음이 특징입니다(프랑스 언어학자 앙드레 마르티네의 특징).그것은 복잡한 언어적 표현이 의미 있는 요소(예: 형태소나 단어)로 분해될 수 있다는 것을 의미하는데, 이는 다시 의미에 영향을 미치는 음소라고 불리는 가장 작은 음성 요소들로 구성됩니다.그러나 동물의 신호는 이 이중 구조를 나타내지 않는다.
- 일반적으로, 동물의 발성은 외부의 자극에 대한 반응이며, 시공간에서 제거된 물질을 지칭하지 않는다.먼 거리에서의 관련성 문제, 예를 들어 사냥 전의 늑대 활동이나 꿀벌 춤 언어로 전달되는 정보 등은 몸짓 언어로 다른 개인에게 대신 나타나는 경향이 있다.따라서 발언이 어느 정도 자동 응답인지 그리고 어느 정도 의도적인 의도가 영향을 미치는지 불분명하다.
- 인간의 언어와는 대조적으로, 동물의 의사소통 시스템은 일반적으로 개념적인 일반화를 표현할 수 없다.(고양이와 일부 영장류는 주목할 [81]만한 예외일 수 있다.
- 인간의 언어는 요소를 결합하여 새로운 메시지(창의성이라고 알려진 속성)를 생성합니다.이것의 한 가지 요인은 인간의 언어 성장이 개념적 아이디어와 가상의 구조에 기초하고 있다는 것이다. 둘 다 동물보다 인간의 능력이 훨씬 더 크다.비록 동물 문화에 대한 현재의 연구가 여전히 많은 새로운 발견과 함께 진행 중인 과정이지만, 이것은 동물의 의사소통 체계에서 훨씬 덜 흔하게 나타난다.2009년에는 부착이 캠벨모나원숭이의 [82]호출 의미에 영향을 미칠 수 있다는 보고가 있었다.
in communication ( 오류)
특정 상황이 [83]적용되면 동물들 간의 의사소통에서 오류가 발생할 수 있습니다.이러한 상황에는, 통신하는 2개의 피사체간의 거리나 상황의 「청취자」에게 전달되는 신호의 복잡성이 포함됩니다."가수"가 통신 위치를 속여 더 많은 [84]오류를 발생시킬 수 있기 때문에 통신 위치가 "청취자"에게 항상 명확하지는 않을 수 있습니다.
「 」를 참조해 주세요.
레퍼런스
- ^ "Animal communication". Encyclopedia Britannica. Retrieved 2020-10-31.
- ^ Seyfarth, Robert M.; Cheney, Dorothy L. (2003-02-01). "Signalers and Receivers in Animal Communication". Annual Review of Psychology. 54 (1): 145–173. doi:10.1146/annurev.psych.54.101601.145121. ISSN 0066-4308. PMID 12359915.
- ^ a b Maynard-Smith and Harper, 2003
- ^ Tinbergen, N.; Perdeck, A. C. (1951-01-01). "On the Stimulus Situation Releasing the Begging Response in the Newly Hatched Herring Gull Chick (Larus Argentatus Argentatus Pont.)". Behaviour. 3 (1): 1–39. doi:10.1163/156853951X00197. ISSN 0005-7959.
- ^ Pollick, Amy S.; Waal, Frans B. M. de (2007-05-08). "Ape gestures and language evolution". Proceedings of the National Academy of Sciences. 104 (19): 8184–8189. Bibcode:2007PNAS..104.8184P. doi:10.1073/pnas.0702624104. ISSN 0027-8424. PMC 1876592. PMID 17470779.
- ^ "Facial display and blushing: Means of visual communication in blue-and-yellow macaws (Ara Ararauna)?". ProQuest 2091762411.
{{cite journal}}:Cite 저널 요구 사항journal=(도움말) - ^ Mogil, Jeffrey S. (April 2009). "Animal models of pain: progress and challenges". Nature Reviews Neuroscience. 10 (4): 283–294. doi:10.1038/nrn2606. ISSN 1471-003X. PMID 19259101. S2CID 205504814.
- ^ a b Shepherd, Stephen V. (2010-03-19). "Following Gaze: Gaze-Following Behavior as a Window into Social Cognition". Frontiers in Integrative Neuroscience. 4: 5. doi:10.3389/fnint.2010.00005. ISSN 1662-5145. PMC 2859805. PMID 20428494.
- ^ Range, Friederike; Virányi, Zsófia (2011-02-23). Wylie, Doug (ed.). "Development of Gaze Following Abilities in Wolves (Canis Lupus)". PLOS ONE. 6 (2): e16888. Bibcode:2011PLoSO...616888R. doi:10.1371/journal.pone.0016888. ISSN 1932-6203. PMC 3044139. PMID 21373192.
- ^ Cloney, R.A.; Florey, E. (1968). "Ultrastructure of cephalopod chromatophore organs". Z. Zellforsch Mikrosk. Anat. 89 (2): 250–280. doi:10.1007/bf00347297. PMID 5700268. S2CID 26566732.
- ^ Hanlon, R.T.; Messenger, J.B. (1996). Cephalopod Behaviour. Cambridge University Press. p. 121. ISBN 978-0-521-64583-6.
- ^ Williams, Sarah (2012). "Two-faced fish tricks competitors". Science Now. Archived from the original on March 8, 2013. Retrieved March 16, 2013.
- ^ Motluk, Alison (2001). "Big Bottom". New Scientist. 19 (7).
- ^ 심해 오징어가 빛나는 색소 침착을 통해 의사소통을 할 수 있다는 것을 연구원들이 발견하다
- ^ Mason, Julia (July 18, 2018). "Glowing in the Deep". The Dish on Science.
- ^ "Bioluminescence Smithsonian Ocean". ocean.si.edu. Retrieved 2020-10-25.
- ^ Ehrlich, Paul R.; David S. Dobkin & Darryl Wheye. ""Bird Voices" and "Vocal Development" from Birds of Stanford essays". Retrieved 9 Sep 2008.
- ^ Schwartzkopff, J (January 1977). "Auditory Communication in Lower Animals: Role of Auditory Physiology". Annual Review of Psychology. 28 (1): 61–84. doi:10.1146/annurev.ps.28.020177.000425. ISSN 0066-4308. PMID 324382.
- ^ Slabbekoorn, Hans; Smith, Thomas B (2002-04-29). "Bird song, ecology and speciation". Philosophical Transactions of the Royal Society B: Biological Sciences. 357 (1420): 493–503. doi:10.1098/rstb.2001.1056. ISSN 0962-8436. PMC 1692962. PMID 12028787.
- ^ Mikula, P.; Valcu, M.; Brumm, H.; Bulla, M.; Forstmeier, W.; Petrusková, T.; Kempenaers, B. & Albrecht, T. (2021). "A global analysis of song frequency in passerines provides no support for the acoustic adaptation hypothesis but suggests a role for sexual selection". Ecology Letters. 24 (3): 477–486. doi:10.1111/ele.13662. PMID 33314573.
- ^ Zuberbühler, Klaus (2001). "Predator-specific alarm calls in Campbell's monkeys, Cercopithecus campbelli" (PDF). Behavioral Ecology and Sociobiology. 5: 414–422.
- ^ Boughman, J W (1998-02-07). "Vocal learning by greater spear-nosed bats". Proceedings of the Royal Society B: Biological Sciences. 265 (1392): 227–233. doi:10.1098/rspb.1998.0286. ISSN 0962-8452. PMC 1688873. PMID 9493408.
- ^ Price, Tabitha; Wadewitz, Philip; Cheney, Dorothy; Seyfarth, Robert; Hammerschmidt, Kurt; Fischer, Julia (2015-08-19). "Vervets revisited: A quantitative analysis of alarm call structure and context specificity". Scientific Reports. 5: 13220. Bibcode:2015NatSR...513220P. doi:10.1038/srep13220. ISSN 2045-2322. PMC 4541072. PMID 26286236.
- ^ Jabr, Ferris (2017-05-12). "Can Prairie Dogs Talk? (Published 2017)". The New York Times. ISSN 0362-4331. Retrieved 2020-10-25.
- ^ "New Language Discovered: Prairiedogese". NPR.org. Retrieved 2020-10-25.
- ^ "YIPS, BARKS AND CHIRPS: THE LANGUAGE OF PRAIRIE DOGS". 2016-10-27. Archived from the original on 2016-10-27. Retrieved 2020-10-25.
- ^ "Prairie dogs' language decoded by scientists CBC News". CBC. Retrieved 2020-10-25.
- ^ January 2006, Bjorn Carey 03 (3 January 2006). "Whales Found to Speak in Dialects". livescience.com. Retrieved 2020-10-25.
- ^ Weilgart, Linda; Whitehead, H. (1997-05-01). "Group-specific dialects and geographical variation in coda repertoire in South Pacific sperm whales". Behavioral Ecology and Sociobiology. 40 (5): 277–285. doi:10.1007/s002650050343. ISSN 1432-0762. S2CID 11845118.
- ^ Ladich, Friedrich (2001). "Sound-generating and -detecting motor system in catfish: Design of swimbladder muscles in doradids and pimelodids". The Anatomical Record. 263 (3): 297–306. doi:10.1002/ar.1105. ISSN 1097-0185. PMID 11455539. S2CID 24896586.
- ^ a b c d e Searcy, William A. (March 2013). "Principles of Animal Communication. Second Edition. By Jack W. Bradbury and Sandra L. Vehrencamp. Sunderland (Massachusetts): Sinauer Associates. $99.95. xiv + 697 p.; ill. + C-1–C-6 (credits) + I-1–I-47 (index). ISBN: 978-0-87893-045-6. 2011". The Quarterly Review of Biology. 88 (1): 48. doi:10.1086/669301. ISSN 0033-5770.
- ^ 프레리독 알람은 포식자 색에 대한 라벨을 인코딩합니다.작성자 : N. Slobodchiko V · Andrea Paseka · Jennifer L. Verdolin.2008년 12월 31일 Springer-Verlag 경유로 발행
- ^ 노란배 마못을 부르는 알람: I.Danil T에 의한 상황변수 알람콜의 의미Blumstein & Kenneth B.캔자스 대학교 아미티지 시스템학 및 생태학과동물의 행동, UCLA 생명과학 출판
- ^ Brown, Grant E.; Chivers, Douglas P.; Smith, R. Jan F. (1995-02-01). "Localized defecation by pike: a response to labelling by cyprinid alarm pheromone?". Behavioral Ecology and Sociobiology. 36 (2): 105–110. doi:10.1007/BF00170715. ISSN 1432-0762. S2CID 31875357.
- ^ Brown, Grant; Adrian, James; Patton, Todd; Chivers, Douglas (Dec 2001). "Fathead minnows learn to recognize predator odor when exposed to concentrations of artificial alarm pheromone below their behavioral-response threshold". Canadian Journal of Zoology. 79 (12): 2239–2245. doi:10.1139/z01-194. ProQuest 220512135 – via ProQuest.
- ^ a b "Acid Rain Is Totally So Last Century, Right? Not Exactly: A Canadian scientist explains how acid rain is still making its mark – IISD Experimental Lakes Area". 16 May 2018. Retrieved 2020-10-26.
- ^ du P. Bothma, J.; le Richet, E. A. N. (1995-04-01). "Evidence of the use of rubbing, scent-marking andscratching-posts by Kalahari leopards". Journal of Arid Environments. 29 (4): 511–517. Bibcode:1995JArEn..29..511D. doi:10.1016/S0140-1963(95)80023-9. ISSN 0140-1963.
- ^ Gosling, L. Morris; Roberts, S. Craig (2001). "Scent-marking by male mammals: cheat-proof signals to competitors and mates" (PDF). Advances in the Study of Behavior. 30: 169–217. doi:10.1016/S0065-3454(01)80007-3. ISBN 9780120045303.
- ^ du P. Bothma, J.; Richet, E.A.N. le (1995). "Evidence of the use of rubbing, scent-marking andscratching-posts by Kalahari leopards". Journal of Arid Environments. 29 (4): 511–517. Bibcode:1995JArEn..29..511D. doi:10.1016/s0140-1963(95)80023-9.
- ^ Clapham, Melanie. Chemical signalling in brown bears, ursus arctos : an assessment of scent marking strategies and social function. OCLC 1065010384.
- ^ "Electrocommunication". archive.fo. 2012-05-29. Archived from the original on 2012-05-29. Retrieved 2020-10-26.
- ^ Worm, Martin; Kirschbaum, Frank; von der Emde, Gerhard (2017-09-13). "Social interactions between live and artificial weakly electric fish: Electrocommunication and locomotor behavior of Mormyrus rume proboscirostris towards a mobile dummy fish". PLOS ONE. 12 (9): e0184622. Bibcode:2017PLoSO..1284622W. doi:10.1371/journal.pone.0184622. PMC 5597219. PMID 28902915.
- ^ Donati, Elisa; Worm, Martin; Mintchev, Stefano; van der Wiel, Marleen; Benelli, Giovanni; von der Emde, Gerhard; Stefanini, Cesare (2016-12-01). "Investigation of Collective Behaviour and Electrocommunication in the Weakly Electric Fish, Mormyrus rume, through a biomimetic Robotic Dummy Fish". Bioinspiration & Biomimetics. 11 (6): 066009. Bibcode:2016BiBi...11f6009D. doi:10.1088/1748-3190/11/6/066009. ISSN 1748-3190. PMID 27906686. S2CID 26047199.
- ^ Kuo, Zing (Jan 1, 1960). "Studies on the basic factors in animal fighting: VI. Inter-species coexistence in birds". The Journal of Genetic Psychology. 97: 15. ProQuest 1297122533 – via ProQuest.
- ^ Yamamoto, Maria Emilia; Araujo, Arrilton; Arruda, Maria de Fatima; Lima, Ana Karinne Moreira; Siqueira, Jose de Oliveira; Hattori, Wallisen Tadashi (2014-11-01). "Male and female breeding strategies in a cooperative primate". Behavioural Processes. Neotropical Behaviour. 109: 27–33. doi:10.1016/j.beproc.2014.06.009. ISSN 0376-6357. PMID 25010563. S2CID 205979349.
- ^ Dunbar, R. I. M. (2010-02-01). "The social role of touch in humans and primates: Behavioural function and neurobiological mechanisms". Neuroscience & Biobehavioral Reviews. Touch, Temperature, Pain/Itch and Pleasure. 34 (2): 260–268. doi:10.1016/j.neubiorev.2008.07.001. ISSN 0149-7634. PMID 18662717. S2CID 30450770.
- ^ "Ants' body odor, physical contact get worker ants working, study finds". news.stanford.edu. Retrieved 2020-10-26.
- ^ Sukhchuluun, Gansukh; Zhang, Xue-Ying; Chi, Qing-Sheng; Wang, De-Hua (2018-05-18). "Huddling Conserves Energy, Decreases Core Body Temperature, but Increases Activity in Brandt's Voles (Lasiopodomys brandtii)". Frontiers in Physiology. 9: 563. doi:10.3389/fphys.2018.00563. ISSN 1664-042X. PMC 5968109. PMID 29867585.
- ^ "Coral Reef Protection: What Are Coral Reefs? Habitat Protection US EPA". archive.epa.gov. Retrieved 2020-10-26.
- ^ Cocroft, Reginald (October 2001). "Vibrational Communication and the Ecology of Group-Living, Herbivorous Insects". Integrative and Comparative Biology. 41: 1215–1221 – via Oxford Academic.
- ^ Hill, Peggy (October 2001). "Vibration and Animal Communication: A Review". Integrative and Comparative Biology. 41: 1135–1142 – via Oxford Academic.
- ^ Narins, Peter (April 1990). "Seismic Communication in Anuran Amphibians". American Institute of Biological Sciences. 40: 268–274 – via Oxford Academic.
- ^ Kardong, Kenneth V.; Mackessy, Stephen P. (1991). "The Strike Behavior of a Congenitally Blind Rattlesnake". Journal of Herpetology. 25 (2): 208–211. doi:10.2307/1564650. ISSN 0022-1511. JSTOR 1564650.
- ^ Bakken, George S.; Krochmal, Aaron R. (2007-08-15). "The imaging properties and sensitivity of the facial pits of pitvipers as determined by optical and heat-transfer analysis". Journal of Experimental Biology. 210 (16): 2801–2810. doi:10.1242/jeb.006965. ISSN 0022-0949. PMID 17690227. S2CID 25037159.
- ^ Pough, F.; Andrews, Robin; Cadle, John; Crump, Martha; Savitzky, Alan; Wells, Kentwood (2003-01-01). "Herpetology". Herpetology (Third ed.): 726.
- ^ a b Gracheva, Elena O.; Ingolia, Nicholas T.; Kelly, Yvonne M.; Cordero-Morales, Julio F.; Hollopeter, Gunther; Chesler, Alexander T.; Sánchez, Elda E.; Perez, John C.; Weissman, Jonathan S.; Julius, David (April 2010). "Molecular basis of infrared detection by snakes". Nature. 464 (7291): 1006–1011. Bibcode:2010Natur.464.1006G. doi:10.1038/nature08943. ISSN 1476-4687. PMC 2855400. PMID 20228791.
- ^ Kürten, L.; Schmidt, U.; Schäfer, K. (1984-06-01). "Warm and cold receptors in the nose of the vampire bat Desmodus rotundas". Naturwissenschaften. 71 (6): 327–328. Bibcode:1984NW.....71..327K. doi:10.1007/BF00396621. ISSN 1432-1904. PMID 6472483. S2CID 31899356.
- ^ Wilson, Ben; Dill, Lawrence M. (March 2002). "Pacific herring respond to stimulated odontocete echolocation sounds". Canadian Journal of Fisheries & Aquatic Sciences. 59 (3): 542. doi:10.1139/f02-029.
- ^ Haldar, Vivekananda; Chakraborty, Niladri (2017-07-01). "A novel evolutionary technique based on electrolocation principle of elephant nose fish and shark: fish electrolocation optimization". Soft Computing. 21 (14): 3827–3848. doi:10.1007/s00500-016-2033-1. ISSN 1433-7479. S2CID 207013387.
- ^ Jones, Gareth (2005-07-12). "Echolocation". Current Biology. 15 (13): R484–R488. doi:10.1016/j.cub.2005.06.051. ISSN 0960-9822. PMID 16005275. S2CID 235311777.
- ^ Vanderelst, Dieter; Steckel, Jan; Boen, Andre; Peremans, Herbert; Holderied, Marc W (2016-08-02). Eichenbaum, Howard (ed.). "Place recognition using batlike sonar". eLife. 5: e14188. doi:10.7554/eLife.14188. ISSN 2050-084X. PMC 4970868. PMID 27481189.
- ^ Stritih, Natasa; Kosi, Alenka (Nov 2017). "Olfactory signaling of aggressive intent in male-male contests of cave crickets (Tropglophilus neglectus; Orthoptera: Rhaphidophoridae)". PLOS ONE. 12 (11): e0187512. Bibcode:2017PLoSO..1287512S. doi:10.1371/journal.pone.0187512. PMC 5675388. PMID 29112984. ProQuest 1961423777.
- ^ Clutton-Brock, Tim (2016-05-31). Mammal Societies. John Wiley & Sons. ISBN 978-1-119-09532-3.
- ^ Jenssen, Thomas A.; Orrell, Kimberly S.; Lovern, Matthew B. (2000). "Sexual Dimorphisms in Aggressive Signal Structure and Use by a Polygynous Lizard, Anolis carolinensis". Copeia. 2000 (1): 140–149. doi:10.1643/0045-8511(2000)2000[0140:SDIASS]2.0.CO;2. ISSN 0045-8511. JSTOR 1448245. S2CID 9813895.
- ^ Bowen, Jon; Heath, Sarah (2005). Behaviour problems in small animals : practical advice for the veterinary team. Edinburgh: Elsevier Saunders. p. 127. ISBN 978-0702027673. Retrieved 16 June 2022.
- ^ Cäsar, Cristiane; Byrne, Richard W.; Hoppitt, William; Young, Robert J.; Zuberbühler, Klaus (2012-08-01). "Evidence for semantic communication in titi monkey alarm calls". Animal Behaviour. 84 (2): 405–411. doi:10.1016/j.anbehav.2012.05.010. ISSN 0003-3472. S2CID 45749417.
- ^ Sasaki, Takao; Hölldobler, Bert; Millar, Jocelyn G.; Pratt, Stephen C. (2014-09-15). "A context-dependent alarm signal in the ant Temnothorax rugatulus". Journal of Experimental Biology. 217 (18): 3229–3236. doi:10.1242/jeb.106849. ISSN 0022-0949. PMID 25013103. S2CID 2102542.
- ^ a b Hasson, O. (October 1991). "Pursuit-deterrent signals: communication between prey and predator". Trends in Ecology & Evolution. 6 (10): 325–329. doi:10.1016/0169-5347(91)90040-5. ISSN 0169-5347. PMID 21232498.
- ^ "Map of Life Vibrational communication in animals". Retrieved 2020-10-28.
- ^ Tauzin, Tibor; Csík, Andor; Kis, Anna; Topál, József (2015). "What or where? The meaning of referential human pointing for dogs (Canis familiaris)" (PDF). Journal of Comparative Psychology. 129 (4): 334–338. doi:10.1037/a0039462. ISSN 1939-2087. PMID 26147704.
- ^ Sean Senechal: 개들도 서명할 수 있다. 개와 소통하는 획기적인 방법, 2009, 랜덤 하우스/크라운/텐 스피드 프레스
- ^ Morris, Desmond (1958). "The Comparative Ethology of Grassfinches (erythrurae) and Mannikins (amadinae)". Proceedings of the Zoological Society of London. 131 (3): 389–439. doi:10.1111/j.1096-3642.1958.tb00695.x. ISSN 1469-7998.
- ^ 리처드 도킨스가 그의 책 이기적인 유전자라는 주제로 장황하게 논의했다.
- ^ Chandler, Christopher H.; Ofria, Charles; Dworkin, Ian (2013). "Runaway Sexual Selection Leads to Good Genes". Evolution. 67 (1): 110–119. doi:10.1111/j.1558-5646.2012.01750.x. ISSN 0014-3820. JSTOR 23327705. PMID 23289565. S2CID 15929198.
- ^ a b V. M. Janik, L. S. Sayigh 및 R. S. Wells: "시그니처 호루라기 모양은 병코돌고래에게 신원 정보를 전달한다", 미국국립과학원회보 제103권 제21호, 2006년 5월 23일
- ^ Smith, Eric Alden (2017-09-29). Evolutionary Ecology and Human Behavior. Routledge. ISBN 978-1-351-52132-1.
- ^ Hare, Brian; Call, Josep; Tomasello, Michael (1998-01-01). "Communication of Food Location Between Human and Dog (Canis Familiaris)". Evolution of Communication. 2 (1): 137–159. doi:10.1075/eoc.2.1.06har. ISSN 1387-5337.
- ^ Tauzin, Tibor; Csík, Andor; Kis, Anna; Topál, József (November 2015). "What or where? The meaning of referential human pointing for dogs (Canis familiaris)". Journal of Comparative Psychology. 129 (4): 334–338. doi:10.1037/a0039462. ISSN 1939-2087. PMID 26147704.
- ^ Guo, Kun; Meints, Kerstin; Hall, Charlotte; Hall, Sophie; Mills, Daniel (2009-05-01). "Left gaze bias in humans, rhesus monkeys and domestic dogs". Animal Cognition. 12 (3): 409–418. doi:10.1007/s10071-008-0199-3. ISSN 1435-9456. PMID 18925420. S2CID 5661394.
- ^ "Do animals have language? - Michele Bishop". TED Ed. 10 September 2015. Retrieved 11 September 2015.
- ^ Thompson, Roger K. R.; Oden, David L. (2000). "Categorical Perception and Conceptual Judgments by Nonhuman Primates: The Paleological Monkey and the Analogical Ape". Cognitive Science. 24 (3): 363–396. doi:10.1207/s15516709cog2403_2. ISSN 1551-6709.
- ^ "Rudiments of Language Discovered in Monkeys". Wired. ISSN 1059-1028. Retrieved 2020-10-31.
- ^ Real, Leslie A. (1994-12-15). Behavioral Mechanisms in Evolutionary Ecology. University of Chicago Press. ISBN 978-0-226-70595-8.
- ^ Gergely, Anna; Compton, Anna; Newberry, Ruth; Miklósi, Ádám (Apr 2016). "Social Interaction with an "Unidentified Moving Object" Elicits A-Not_B Error in Domestic Dogs". PLOS ONE. 11 (4): e0151600. Bibcode:2016PLoSO..1151600G. doi:10.1371/journal.pone.0151600. PMC 4830451. PMID 27073867. S2CID 16369609.
외부 링크
- 애니멀커뮤니케이터– 다큐멘터리
- Zoosemiotics: Web상의 동물 커뮤니케이션 2005-10-25 Wayback Machine에 보관
- 동물 소통 프로젝트
- 국제생물음향학회의 동물언어 연구.
- Animal 듣기와 다운로드 할 수 있는 동물의 소리가 다르다.
- 영국 도서관 사운드 아카이브(British Library Sound Archive)는 전 세계의 동물 소리와 자연 대기에 대한 15만 건 이상의 녹음을 보유하고 있습니다.
| 나뭇가지 | |
|---|---|
| 동물행동학자 | |
| 사회 | |
| 일지 | |


