암피드로무스
Amphidromus| 암피드로무스 | |
|---|---|
 | |
| 암피드로무스 로즈골라비아투스 종은 덱스트랄 껍질 코일링을 가지고 있다. | |
 | |
| 암피드로무스 푸스콜라브리스 종은 부신성 껍데기를 코일링하고 있다. | |
| 과학적 분류 | |
| 킹덤: | 애니멀리아 |
| 망울: | 몰루스카 |
| 클래스: | 가스트로포다류 |
| 순서: | 질로마토포라속 |
| 슈퍼 패밀리: | 헬리코이데아과 |
| 패밀리: | 까마네과 |
| 하위 패밀리: | 까메니네아과 |
| 속: | 암피드로무스 앨버스, 1850년[1] |
| 종류종 | |
| 암피드로무스 퍼버서스 | |
| 종 | |
| 텍스트 보기 | |
| 다양성[2][3] | |
| 110여 종 | |
| 동의어 | |
| |
암피드로무스는 열대공기를 내뿜는 육지 달팽이, 카마네과과에 속하는 육상 풀몬산 위스트로포드 연체동물의 속이다.암피드로무스의 껍질은 최대 치수의 25mm(0.98인치)에서 75mm(3.0인치)까지 비교적 크고 특히 화려하다.18세기 동안, 그것들은 여행자들과 탐험가들이 유럽으로 가져온 최초의 인도네시아 육상 달팽이 껍질 중 하나였다.그 이후로, 그 속은 광범위하게 연구되었다: 19세기 초에서 20세기 중반까지의 기간 동안 자연주의자들과 동물학자들에 의해 몇 개의 포괄적인 모노그래프와 카탈로그가 저술되었다.현대 연구는 분류학 문제를 해결하는 것뿐만 아니라 그룹 내의 진화적 관계를 더 잘 이해하는 데 초점을 맞추었다.
암피드로무스속은 덱스트랄 껍데기를 가진 종과 부신각 껍데기를 가진 종을 포함한다는 점에서 특이하다.또한, 이 속 내의 일부 종은 그들의 개체군이 왼손잡이와 오른손잡이의 껍데기 코칭을 가진 개인을 동시에 포함하기 때문에 특히 주목할 만하다.이것은 극히 드문 현상이며 생물학자들에게는 매우 흥미로운 현상이다.암피드로무스의 부드러운 해부학에 초점을 맞춘 연구들이 산재하고 단편적이다.내부 해부학에 대한 정보는 몇 종에서만 알려져 있으며, 더 큰 규모의 비교 형태학 연구는 지금까지 수행된 적이 없다.
암피드로무스속에 있는 종들은 수목류다. 즉, 그들은 나무 달팽이다.그러나 이들의 습관에 대한 보다 자세한 정보는 여전히 부족하다.이러한 달팽이의 일반적인 먹이 습관은 알려져 있지 않지만, 몇몇 종은 미세한 곰팡이, 이끼 또는 육지 해조류를 먹고 사는 것으로 알려져 있다.암피드로무스 자체는 새, 뱀, 그리고 아마도 쥐와 같은 작은 포유류들에 의해 먹잇감이 될 것이다.
분류학 및 역사
.jpg)
총칭은 '양쪽으로'를 뜻하는 고대 그리스어 암피(í (μμ)와 '달린다'는 뜻의 드로모스( (δμος)에서 따온 것으로, 껍질의 서로 다른 치랄성을 암시하고 있다.[5]암피드로무스의 껍질은 비교적 크고 매우 다채롭다; 그 중 상당수는 18세기 동안 여행자와 탐험가들이 유럽으로 가져온 최초의 인도네시아 육상 달팽이 껍질 중 하나이다.비교적으로 말하면, 말라카학자들은 훨씬 적은 수의 표본을 모았다.[4]
1800년 이전에 몇몇 종과 형태가 설명되었는데, 그들 대부분은 불충분한 지역성 데이터를 가지고 있다.적어도 암피드로무스 라에부스 (Müler, 1774년)와 A. perversus f. aureus Martyn, 1784년 – 여전히 (2017년 현재) 정확한 지역으로부터는 아직 보고되지 않았다.19세기 전반기에 많은 종과 품종이 이름지어졌는데, 다시 보통 좋지 않은 지역성 데이터를 가지고 있었다.에두아르트 폰 마르텐스(1867)가 그의 모노그래프를[6] 출판하기 전까지는 이 속 안에 있는 종의 전체 콤플렉스를 다루려는 시도가 있었다.1867년 단문자에는 속내의 변동과 종의 지리적 분포의 문제에 관한 상당한 정보가 수록되어 있었다.폰 마르텐스에서 유래한 많은 개념들이 아직도 (2017년 현재) 사용되고 있다.[4]
1896년 휴 풀턴은[7] 142개의 특정 및 변종 명칭을 총 64종을 포함하는 18개 종으로 정리했다.헨리 아우구스투스 필스브리(Henry Augustus Philsbry)의 1900년 단문학[8] 해설서가 등장했을 때 암피드로무스(Ampidromus) 속종의 종수는 81종으로 늘어났고, 이 종들은 19개 조에 배치되었다.필스브리의 연구는 그 속에 대한 유일한 삽화적 모노그래프로 남아 있으며, 여전히 그 속에 대한 어떤 진지한 연구에도 없어서는 안 될 것으로 여겨지고 있다.[4]
1900년 이후 암피드로무스에 대한 주요 분류학 연구는 범위 내에서 공정론적(일부 영토나 지역의 동물에 대한 연구)이었다.미국의 말라리아학자 폴 바르츠흐(1917, 1918, 1919)[9][10][11]가 필리핀 종, 베른하르트 렌슈(1932),[12] 테라 판 벤트헴 주팅(1950, 1959년)[13][14]이 자바와 수마트라 종에 대해 쓴 논문은 특히 포괄적이다.잠재적으로 가장 가치[4] 있는 기여는 A. 콘트라리우스와 티모르에 대한 A. 반사요법 내의 변동을 논한 커트 하니엘(1921년)[15]의 공헌이다; 색과 형태의 변화는 일련의 색판에 잘 나타나 있었다.
1900년 이후에 출판된 문헌에는 새로운 색채 형태와 아종에 대한 산재된 설명이 많이 포함되어 있다.명목록에 수록된 309개의 이름 중 111개(35.9%)가 필스브리(1900) 이후 출판되었다.아돌프 마이클 질치(1953)는 [16]센켄베르그 박물관에 활자 표본을 나열하고, 이전에는 품위가 없던 많은 종들을 삽화했다.Frank Fortescue Laidlaw & Alan Solem(1961)은 74종의 이름을 인정했고, 반다 제도에서 나온 물질이 아마도 원하지 않는 종을 대표한다고 생각했다.필스브리의 모노그래프가 등장한 후 라이틀로앤솔렘이 인정한 종 중 11종이 묘사되었다.그러나 필스브리가 인정한 여러 종은 이후 하위특정 또는 변종 상태에 종속되어 왔으며, 몇몇 명칭은 보다 최근의 수집가들에 의해 입증되지 않은 100년 된 참조에 기초하기 때문에 인서태 세디스로 이전되었다.사실, Laidlaw및 공부;Solem(1961년), Laidlaw및 많은, 색 형태 묘사되는 관계에 관한 Solem의 결론 종들이 새로운 샘플에서 변화의 Haniel(1921년)에 의해 그의 pione에 알 수 있는 범위에서 별로 취해 진 태생의 논문에 광범위한 그의 접시들과 함께 추가적을 이루고 있다.스터드은 후 그는y.[4]
특성화
암피드로무스속에는 보통 부드럽고 윤기가 흐르며, 밝은 색상이 있으며, 길쭉한 껍질이나 원추, 덱스트랄 또는 불신적으로 코일된 껍질이 있다.조개 껍질은 적당히 크고, 최대 치수는 25mm(0.98인치)에서 70mm(2.8인치)까지이며, 볼록한 휘파람은 6~8개까지 있다.그들의 색채 패턴은 보통 단색의 황색이나 녹색을 띠지만, 얼룩덜룩해질 수 있다.개구부는 비스듬하거나 난형이며, 톱니나 접힘이 없으며, 개구부 높이는 전체 껍질 높이의 2/5에서 3분의 1에 이른다.근막은 확장 및/또는 반사되며, 때로는 두꺼워진다.콜럼벨라는 곧거나 재발할 수 있고, 두정맥은 잘 발달할 수 있도록 약하며, 탯줄은 열거나 닫을 수 있다.라둘라는 주걱으로 뱉어졌고, 톱니가 일렬로 배열되어 있으며, 보통 단첨판 중심 치아와 이첨판 또는 삼첨판 측면 치아를 가지고 있다.턱은 가늘고 약하며, 평평한 갈비뼈가 낮다.창백한 부위는 매우 길고 좁은 신장을 가진 S자형이다.성기는 전형적인 가마우지의 것으로, 긴 정맥 콘센트, 수축근의 삽입이 낮은 짧은 페니스, 짧거나 긴 경피용 카에쿰(평활근과 맹장)을 가지고 있다.정조세포는 단면에서 오각형의 윤곽을 가지고 있다.암피드로무스는 전형적으로 수목 동물이다.[4][17]
셸 설명
암피드로무스의 껍질은 높이가 1~3인치 정도로 비교적 크고 화려하다.암피드로무스는 5-8 Whorls의 신장-원뿔 또는 난산-원뿔 헬리코이드 껍질을 가지고 있다.껍질은 얇고 연약하거나 매우 무겁고 견고할 수 있으며, 껍질 구조와 분포 또는 서식지의 상관관계가 알려져 있지 않다.[4]
셸 코일링
이 속 안에 있는 어떤 종에서는 껍질이 항상 오른쪽으로 굽고, 다른 종에서는 항상 왼쪽으로 구부러진다.그러나, 이 속에서는 상당한 수의 종들이 "암피드로민"이다. 이 용어는 왼손잡이와 오른손잡이의 껍질 코일링이 동일한 개체군 내에서 발견된다는 것을 의미한다.껍질 코일링 방향으로는 '폴리모르픽'이라고 할 수 있지만, 가능한 껍질 코일링의 종류는 두 가지에 불과하기 때문에 코일링에서는 '다이모르픽'으로 표현된다.두 종류의 껍데기 코일링은 거의 동일한 숫자로 일부 종에서 발생하며, 다른 종은 한 단계의 뚜렷한 우세를 가지고 있다.암피드로무스에는 아직 이 인물의 유전에 대한 정보가 없다.[4]
extinct,[18]이 속 Amphidromus,을 110종을 포함하는, 독특한 비대칭 animals,[18]의 진화의 유학을 왜 이 종의 보전 필수적인 수입 이것은 유용하다 왜냐하면 그 속 Partula과 Achatinella 이내에 등amphidromine 고둥의 거의 모든 다른 종들이 이미 되었다.에 ance생물학자

A. 플로레시아누스, 하위유전 신드로무스에서 조개 코일링은 보통 불신이다.축척 막대 10 mm.

그러나 여기 보이는 것처럼 A. perversus에 코일링된 껍질은 또한 불신적일 수 있다.

A. perversus의 부신각(왼쪽) 및 덱스트랄 껍질(오른쪽)의 구면도




조개 모양과 조각

암피드로무스 종의 껍데기 껍질은 적당히 볼록하고 몇 가지 예외만 있을 뿐 매끈하거나 희미한 성장선 조각이 있다.그러나 적당히 무거운 사선 방사형 갈비뼈의 조각은 속에서는 적어도 네 번 이상 따로 나타났으며, 다음과 같은 종에서 볼 수 있다.베트남 빈딘 성에서 암피드로무스의 대가 스미스, 캄보디아에서 온 비기니 모레트, A. bengie morlet; A. 셀레베에서 온 헤카리 타파론-카네프리와 자바와 수마트라에서 온 A. 팰리스-A. 윈터니 콤플렉스.늑골과 상관관계가 있는 것은 가볍고 단색이며, 구멍이 크고 입술이 벌겋게 부풀어 있는 얇은 껍질이다.다른 종의 많은 단단한 껍질들은 표면이 약간 거칠어지는 것을 보여주지만, 이것은 위에서 언급된 갈비뼈 조각과는 매우 다르다.[4]
조리개는 일반적으로 크며, 껍질 높이의 약 2/5에서 3분의 1까지 다양하며, 종종 같은 모집단 내에 있다.보통 입술은 적어도 어느 정도 확장되어 있으며, A.반사성 샤프만과 A. 윈터티(Pfeiffer) 바. inauris Fulton에서는 입술은 플레어링이라고만 부를 수 있다.A. perversus (Linnaeus)와 다른 대부분의 굵은 발톱 종에서는 입술이 내적으로 두꺼워져서 팽창하는 과정에서 "롤"을 형성하며, 두정맥이 매우 무겁다.얇은 껍질 종에서, 입술은 보통 단순한 반사된 가장자리 입니다.탯줄 부위는 부분적으로 열거나 거의 닫히거나 밀봉할 수 있다.이 기능은 때때로 특정 식별에 유용한 기준을 제공한다.두정벽의 각도는 다양하지만 이에 대한 정확한 정보는 집계되지 않았다.[4]
일반적으로 껍질의 고래는 크기가 다소 규칙적으로 증가하지만, A. 시니스트랄리스(리브)와 A. 헤카리 타파론-카네프리와 같이 밀접하게 연관되어 있는 종은 고래 증가의 정도가 상당히 다를 수 있다.이용 가능한 자료의 대부분이 통계적 처리에 불충분했기 때문에, 이러한 차이를 인문학적으로 표현하려는 어떠한 시도도 이루어지지 않았다.껍질의 실제 치수는 종 내부와 종 간에 크게 다르다.최소 성인 크기는 약 21mm 높이, 관찰된 최대 크기는 약 75mm이다.종 내에서 성인의 크기에는 큰 차이가 없다. 특히 A. 마쿨리퍼스, A. 시넨시스, A. 엔토밥투스 등 소수의 종만이 총 7~8mm 이상의 성인의 크기 변화를 가지고 있다.[4]
셸 배색


속 내에서 껍질 변동의 가장 중요한 단 하나의 측면은 색 패터닝이다.일반적으로 많은 수목 달팽이는 밝은 색상으로, 폭식성 생식기인 드라이마이우스와 리구우스, 세폴리드 폴리미타, 카마네이드 파푸이나 등이 명백한 예다.그러나 폴리미타, 리구우스, 암피드로무스는 특히 색의 변화로 유명하다.암피드로무스의 기본 바탕색은 노란색으로 나타나며, 마모된 표본은 거의 색이 없는 것으로 보이기 때문에 이 색은 대개 껍질의 표면층에 국한된 것이다(암피드로무스 엔토밥투스 제외).어떤 종에서는 배경색이 희고, 몇몇 종은 어두운 배경색을 가지고 있다.무형의 고래는 창백, 자주색, 갈색 또는 검은색이며, 이것은 때때로 모집단 내에서 다르다(A. 쿼드라시).예를 들어, A. Schomburgki와 같은 몇몇 종은 낙엽성 녹색 근막을 가지고 있다.[4]
연속적인 영역 패턴은 백색 부분부속 밴드(A. similis), 중주변 색소침착(A. perversus var. infraviridis), 부분부속 색상 라인(A. columnellaris), 넓은 나선 색상 밴드(A. metabletus, A. webbi) 또는 좁은 나선부(A. laevus)의 형태를 취할 수 있다.중단된 조닝은 (A. 마쿨라투스)의 점으로 밴드를 차단하거나 (A. perversus var. 술탄 및 인터럽투스) 구역을 매우 불규칙하게 분할하거나 (A. inversus) 또는 (A. lastrigatus) 증분 성장 선과 평행하는 (A. lastrigatus) 사선 줄의 형성 또는 거의 모든 예상 가능한 결합으로 구성될 수 있다.n과 이러한 요인의 변동.종종 그 패턴은 정점에서 신체 윙윙거림(A. (A. 쿼드라시 바에서).조리개, 두정골, 콜럼벨라, 입술, 탯줄 부위는 분홍색, 갈색, 보라색, 흰색 또는 검은색으로 다양하게 표시되어 있다.하니엘(1921년)[15]은 신드로무스형의 두 종 내에서 색의 변화 정도를 명확히 보여주는 여러 가지 색판을 포함하고 있다.A. perversus와 A. maculiperus 하위 유전자 암피드롬us는 똑같이 가변적인 반면, A. inversus와 A. similis와 같은 종은 거의 균일한 색채화다.[4]
아제너스 암피드로무스(Ampidromus)에 있는 대부분의 종의 껍질에서 휴식 단계는 바릭스(Varix)라고 불리는 갈색 또는 검은 방사상 띠의 퇴적으로 표시된다.A. 레뷔스의 껍질이 휴식기 이후 나선형 밴딩이 중단되었다는 증거를 보여주기는 하지만 이것은 하위 게놈 신드로무스에서 드문 것으로 보인다.[4]
종인식
종인식은 형상, 조리개, 등고선, 탯줄 부위, 색상 패턴의 사소한 구조적 변동의 조합에 기초한다.많은 종들이 안정된 색채 패턴을 가지고 있는 반면, 다른 종들은 엄청나게 다양해 보이는 경우인 것 같다.적절한 선택되지 않은 필드 샘플은 단일 지역에서 특정 종의 상대적 안정성 또는 가변성을 더 잘 이해할 수 있게 할 것이다.[4]
해부학
암피드로무스의 부드러운 해부학에 관한 정보는 널리 흩어져 있고 단편적이다.가장 완전한 설명은 그레이트 나투나(나투나 제도)와 제마드자(아남바 제도)의 표본에 대한 아놀드 자코비(1895)의 [19]설명이다.불행히도, 비록 자코비가 해부한 두 종에 해부학적 차이가 존재한다는 것은 분명하지만, 불행히도 우리는 그가 어떤 형태로 작업했는지 알지 못한다, 왜냐하면 그가 그의 소재를 잘못 식별했기 때문이다.그는 논문에서 이 두 종을 암피드로무스 클로로리스와 A. 퍼버서스의 인터럽투스 단계로 지칭했다.그러나 실제로는 암피드로무스 클로리스는 필리핀 제도에서만 발견되는 종이며, 나투나 제도에서는 A. 퍼버스의 인터럽투스 단계가 존재하지 않기 때문에 그럴 수 없다.[4]
칼 아렌드 프리드리히 위그만(1893년, 1898년)[20][21]은 A. 아담시, A. 포르셀라누스, A. 콘트라리우스, A. 시니스트랄리스의 해부학적 일부에 대해 토론하였다.월터 에드워드 콜링게(1901, 1902)는 [22][23]A. 팰리스와 A. 파라켄시스(A. perversus로 보고됨)의 특징을 간략히 언급했다.하니엘(1921년)[15]은 A. 콘트라리우스와 A. 반사요법을 해부했고, 베른하르트 렌슈는 그의 여러 가지 자유주의적 조사에서 몇 가지 흩어진 메모를 발표했다.필스브리(1900년)에는 몇 가지 초기의 노트가 언급되어 있다.[4][8]
반사요소와 폐쇄요소가 있는 길고 좁은 신장, 뚜렷한 성기가 있는 음경 콤플렉스, 경막과 연속되는 경막, 경막과 경막, 미부착 생식관, 질 부속기관의 부족, 신경과 수축근육의 기본 상태 등의 캐릭터ems는 암피드로무스가 까마네과에 포함되도록 지원한다.[17]이 달팽이 무리는 동아시아와 오스트랄라시아의 열대지방에서 매우 다양한 서식지에서 발생하며, 쇄골 스티클로마토포라에서 가장 다양한 가족 중 하나이다.[24]Laidlaw & Solem(1961)은 암피드로무스의 해부학에 대한 추가적인 세부사항을 제공하지 않았지만, 비숍(1977),[25] 솔렘(1983)과 같은 구별되는 저자들의 후속 연구는 생식 시스템이 종인식을 위한 귀중한 데이터를 제공할 수 있다는 것을 입증했다.[26][4]
- 해부를 통해 격리된 생식 시스템
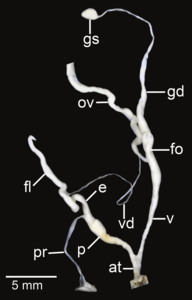
암피드로무스 푸스콜라브리스, at – at – epiphallus; fl – flagellum; fo – free oviditle; gd – gametolytic sac; g – gametolytic sac; 난형성낭; p – pr – penial retractractor 근육; v – vgy; vd; vd; vd; vd; vd; vd; vd; vd – vd – vd – vd – vd – varis

암피드로무스 로즈골라비아투스, ap – 부록, at – 아트리움, e – epiphallus, fl – flagellum, fo - fo - free 난소, gd – gametolytic sac, gs – gametolytic sac, 난소, p – p – pr – penial retractractractractor, v - v 질; vd; vd – vd – vals.
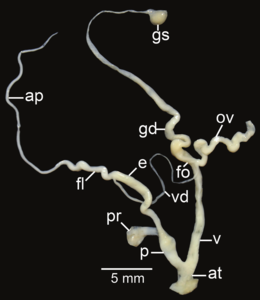
암피드로무스 신드로모이드, ap – 부록, at – 아트리움, e – e – epiphallus, fl – flagellum, fo – free ovolytor, gd – gametolythic sac, gs – gametolytic sac, ot - 난소, p – p – pr – penal retractractractractors; v - v - v - v - v - val.






다양성과 계통발생성
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 수차릿 등에 따른 암피드로무스의 계통발생과 관계(2007)[18] |
1900년 이전에는 남미의 나무 달팽이들과 암피드로무스 껍질의 모양이 비슷한 것이 분류학자들을 오도했었다.그러나 위그만과 야코비의 전파를 통해 암피드로무스의 내적 해부학적 특징이 아시아-인도네시아 카마네과와 같으며, 암피드로무스의 껍데기와 폭식체의 유사성이 단지 병렬주의의 한 예에 불과하다는 것을 분명히 알 수 있었다.[4]
- 하위게놈과 종
라이틀로와 솔렘(1961)은 암피드로무스속 75종을 인정하고, 또 다른 7종의 이름을 삽입성 세디스 밑에 두었다.2010년에는 암피드로무스속 87종이 인정되었다.[18][27][28][4]암피드로무스속 내의 종은 다음 목록과 같이 두 개의 하위 유전자로 나뉜다.[18]
- 서브게너스 암피드로무스 알베르스, 1850년
원래 1758년 린네오스에 의해 헬릭스 퍼버서스로 묘사된 암피드로무스 퍼버서스는 에두아르 폰 마르텐스(1860년)의 후속 명칭에 의해 암피드로무스속(Ampidromus perverus는 암피드로무스 속종의 일종이다.[29]하위유전 암피드로무스의 종은 암피드로민(좌손과 우손 달팽이가 개체군 내에서 발생함)으로 몇 가지 예외를 두고 있다.덱스트랄 세자는 A. givenchyi, A. protania, A. Schomburgki 덱스트로클로루스, A. inversus annamicutus; 그리고 하나의 부신: A. atricallosus classiaris이다.[18]이 종들은 보통 다음과 같은 특성을 가진 껍데기를 가지고 있다: 그들은 크다 (높이는 종종 35 mm (1.4 in)를 초과한다), 그들은 높이/폭 비율이 1.85 미만이다; 그리고 노란색이나 녹색인 껍데기 색깔을 가지고 있다.해부학적으로 그들은 긴 에피팔루스와 편평형을 가지고 있고, 보통 부록이 있다.[4][17]
하위 유전자 안에 있는 두 종을 제외한 모든 종은 신드롬이다.암피드로민 A. 글라우콜라린스와 덱스트랄 A. 크루니도 예외로 한다. kruehni)는 예외다.[28]아돌프 마이클 질치(1960년)의 후속 지명에 의해 하위 게놈 신드로무스의 종류는 A. 콘트라리우스 뮐러(A. contractarius Müler, 1774년)이다.[30]서브게너스 신드로무스의 종은 작은 껍질(높이 보통 35mm(1.4인치) 미만, 높이/폭 비율이 1.85보다 크며, 색상 패턴이 가변적이다.그들은 또한 충수가 부족한 짧은 에피할루스와 플라겔럼을 가지고 있다.[4][17]세 번째 가능한 하위 유전자인 고니오드로무스 뷔로우, 1905년(유형 암피드로무스 뷔로위 프루스토르페르, 1905년)[31]도 문헌에 인용되어 있으나 하위 유전적 상태는 아직 확인되지 않았다.[17]
- 필로제니
Sutcharit과 [18]동료(2007)가 수행한 18종의 16S rDNA의 부분 시퀀스에 대한 분자 분석은 암피드로무스가 단극성 집단임을 나타낸다.그들의 연구에서 최대 시차, 인접 결합 및 최대 가능성 등 구별되는 방법을 통해 얻은 서로 다른 클래도그램이 그들 사이에 일치했다.하위 유전자 암피드로무스에 대해 얻은 위상은 현재의 분류법과 상당히 일치했지만, 시니스트랄 신드로무스 종의 계통생성에서는 그러한 일치성이 보이지 않았다.또한 그들의 결과에 따르면, 에노모르피아는 암피드로무스속 속에 있는 조개껍질의 조개 상태인 것으로 보이며, 이는 덱스트럴적으로 조개껍데기를 조상의 조건으로 하는 일반적인 기대와는 반대되는 것이다.[18]
형태학적으로 동일함에도 불구하고, 암피드로무스 반미츠셀라투스, A. 신겐시스, A. 아레올라투스 등 3종에 속하는 일부 표본은 분명히 mtDNA happlotype의 다혈성 기원을 가지고 있었다.그 결과 토폴로지를 따라 뚜렷한 층에서 같은 종들이 동시에 나타났다. 예를 들어, A. A. areolatus는 각각 A. xiengiensis와 A. semitselatus와 함께 군집화된 Sutcharit와 동료(2007)의 Cladogram에서 각각 다른 두 개의 층에서 발견될 수 있다.저자들에 따르면, 이러한 결과는 수렴성 및 다형성 쉘 색상 패턴으로 설명될 수 있었다(예를 들어, 시료의 쉘은 mtDNA 마커가 유의미한 차이를 보였지만 색상과 모양이 매우 유사했다).또는 mtDNA의 내향적 잡종이나 조상 다형성의 결과일 수도 있다.어떤 경우든 이러한 문제를 더욱 명확히 하기 위해서는 다른 표식자(핵 표식자 또는 기타 mtDNA 표식자) 또는 추가적인 형태학적 문자를 사용한 식물학적 분석은 여전히 필요할 것이다.[18]
화석사
현재 암피드로무스의 신뢰할 수 있는 인간 이전의 화석 발생은 기록되지 않았다.테라 반 벤트헴 주팅(1932)은 [32]자바 중부 삼풍동굴에서 선사시대에 토착민들이 먹었던 A. 필로조나투스 표본을 여러 개 발견했으며, 몇 년 후(반 벤트헴 주팅, 1937년, 페이지 92–94) [33]같은 저자가 자바의 트리닐 침대에서 A. 팰러스의 표본을 한 개씩 발견했다고 보고했다.어떤 기록도 인간의 점유보다 앞서지 못하기 때문에 암피드로무스의 인류 이전의 역사를 조명하지 못한다.[4]
생태학



분배
암피드로무스속은 인도 동부에서 서남아시아(히말라야 산맥에 의해 북쪽으로 한정)에서 호주 북부(베버 라인에 의해 동쪽으로 한정)에 이르는 속이다.[17][26]Amphidromus species are found in localities as follows: from the Garo Hills and Khasi Hills of Meghalaya in northeastern India; throughout Burma, Malay Peninsula, Thailand, Laos, Cambodia, Vietnam, Indonesia as far east as the Sulawesi, Banda Islands, Timor and the Tenimber Islands (but not on Ceram, Buru, Halmahera, Batjan Island, the Obi Islands, 아루와 케이 제도 또는 탈라우 군도와 일부 셀레베시아 위성 섬); 특히 남부 필리핀, 민다나오와 발라박, 팔라완,[4] 그리고 북부 오스트레일리아(솔직히 암피드로무스 코냑으로 표현됨).[26]
서식지와 먹이 주기 습관
암피드로무스 종은 수목 육지 달팽이다.암피드로무스 종의 생활 습관과 생활 방식에 관한 추가 정보는 거의 존재하지 않지만, 이 달팽이들은 일반적으로 나무나 관목 위를 기어다니는 동안 수집되었다.암피드로무스의 식단은 알려져 있지 않지만 암피드로무스 아트리칼로스 페라켄시스는 미세한 곰팡이, 이끼 또는 육지 해조류를 먹고 사는 것으로 생각된다.[4][34]
라이프 사이클
이 속 내에서의 큰 다양성에도 불구하고 2017년 현재 암피드로무스 종에 대한 종합적인 생명사 연구는 여전히 부족하다. 속 내 종들의 행동에 대한 관찰은 극히 일부에 불과하며, 이러한 관찰은 문헌 전체에 흩어져 있다.Eugen Paravicini(1921)[35]의 연구는 웨스트 자바주 팔리만에서 암피드로무스 팰리스투스 바.푸라에서의 난자 발생 행위를 설명했다.파라비시니의 관찰에 따르면 1920년 10월, 서자바 지역 주민들은 이제 막 알을 낳기 시작한 달팽이가 들어 있는 두 개의 "둥지"를 가지고 왔는데, 한 달팽이는 어린 죽순의 바깥 잎을 접어서 뾰족한 코르넷으로 껌을 함께 만들었다.좁은 끝이 위로 향하도록 세로로 매달렸고, 아래 넓은 오프닝이 펼쳐졌다.[4]포대 윗부분은 수거할 때 알이 채워져 있었다.달팽이는 세로축을 중심으로 회전하면서 천천히 내려오더니 충치가 다 찰 때까지 알을 침전시켰다.바구니에 금이 가 달걀이 허공에 노출되면 금방 말라버렸다.포획된 지 이틀 만에 알을 낳는 작업이 끝났고, 달팽이는 더 많은 잎을 접어서 개구부를 닫았다.관찰을 시작할 때 충치가 반 정도 채워졌기 때문에 아마도 4일은 알을 낳는 데 보냈을 것이다.비슷하게 접힌 망고 잎의 두 번째 둥지에는 234개의 알이 들어 있었다.각각의 경우에서 달걀의 부피가 달팽이의 크기를 크게 초과하여, 달걀이 퇴적되기 직전에 캡슐화해야 함을 알 수 있었다.캡슐은 매우 얇았고, 공기에 노출되면 빠르게 건조되었다.10월은 장마가 시작되었고 아마도 지금이 정상적인 번식기일 것이다.포셀라누스 알은 밴 벤트헴 주팅([13]1950, 페이지 493)이 낳은 지 불과 열흘 만에 부화를 시작했다고 신고했다.다른 종들에게도 비슷한 둥지 짓는 습성이 보고되었지만, 생명 주기에 대한 완전한 연구는 발표되지 않았다.1961년까지 활동 주기, 수명, 성장 속도 등에 관한 정보는 입수할 수 없었다.[4]
실투이젠 외(2005)은 말레이시아의 A. inversus 인구의 공간 구조를 설명했다.[36]실투이젠 외(2007년)는 반대편 치질의 달팽이와 교미할 때 성적인 선택이 있다는 것을 발견했다.[37]이것은 왼손잡이 달팽이가 같은 코일링의 달팽이보다 오른손잡이 달팽이와 더 자주 교미한다는 것을 의미한다.또한 이러한 성숙의 성공을 위한 생식계의 정자와 여성 부분의 해부학적 적응이 있다.[37]
포식자
암피드로무스 달팽이의 포식자는 붉은 왕관을 쓴 바베트 메갈라이마 라플레시,[38] 그리고 아마도 다른 새 종들을 포함한다.[34]파레아속 속의 아시아 뱀들은 껍질에서 부드러운 부분을 제거하여 암피드로무스 종을 먹고 사는 것으로 알려져 있다.[39][17]말레이시아의 한 쥐의 소굴에서 암피드로무스의 많은 포탄이 발견되었다.[37]
참고 항목
참조
이 글에는 참고문헌의 공용 도메인 텍스트가 포함되어 있다.[4]
- ^
 앨버스 J. C. (1850).Die Heliceen nach Naturlicher Verwandtschaft: systematisch georduct: 138.
앨버스 J. C. (1850).Die Heliceen nach Naturlicher Verwandtschaft: systematisch georduct: 138. - ^ Sutcharit, Chirasak; Ablett, Jonathan; Tongkerd, Piyoros; Naggs, Fred; Panha, Somsak (30 March 2015). "Illustrated type catalogue of Amphidromus Albers, 1850 in the Natural History Museum, London, and descriptions of two new species". ZooKeys (492): 49–105. doi:10.3897/zookeys.492.8641. ISSN 1313-2970. PMC 4389215. PMID 25878542. Retrieved 8 July 2017.
- ^ a b Inkhavilay, Khamla; Sutcharit, Chirasak; Panha, Somsak (13 June 2017). "Taxonomic review of the tree snail genus Amphidromus Albers, 1850 (Pulmonata: Camaenidae) in Laos, with the description of two new species". European Journal of Taxonomy (330). doi:10.5852/ejt.2017.330. ISSN 2118-9773.
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac 라이틀로 F. F. & 솔렘 A. (1961년)."육지 달팽이속 암피드로무스: 시놉틱 카탈로그"필드아나 동물학 41(4): 505–720.
- ^ Brown, R. W. (1954). Composition of Scientific Words (PDF). Baltimore, Maryland, USA: Published by the author.
- ^ 마르텐스 E. 폰 (1867년).다이 프루시슈 탐험대 나흐 오스트아시엔Zoologischer Theil. 2: Xii, 447 pp, 22 플레이트.
- ^ 풀턴 H. (1896년)."Albers, Ampidromus의 종 목록, 그리고 지금까지 원하지 않았던 종과 품종에 대한 비판적인 메모와 설명이 포함되어 있다."실록과 자연사 잡지 (6)17: 66–94, 번호판 5–7.
- ^ a b c 필스브리 A. (1900년)체계적이고 체계적인 협착학 설명서, 종에 대한 삽화가 포함되어 있다. 두 번째 시리즈: 풀모나타.13권.오스트랄라시아불리물과:볼리엠브리온, 플라코스티유스헬리키과:암피드로무스.253쪽 72쪽 184쪽
- ^ 바트슈 P. (1917)."암피드로무스속 필리핀 육지 달팽이"U.S. Nat. Mus. Bull. 100(1) 파트 1-47, 22판.
- ^ 바트슈 P. (1918)."팔라완 통로 섬에서 암피드로무스속 육지 달팽이"주르. 워싱턴 아카드. 과학 8(11): 361–367.
- ^ 바트슈 P. (1919)."필리핀 섬 땅 포탄에 대한 비판적 발언"프로크. 비올. Soc. Washington 32: 177–184.
- ^ (독일어로) 렌슈 B. (1932년)."Die Molusken Fauna der Kleinen Sunda-Inseln, 발리, 롬복, Sumbawa, Flores und Sumba."졸. 자흐브, 시스템 63: 1-130, 3접시
- ^ a b 반 벤트헴 주팅 T. (1950)."헬리코리온과, 플뢰로돈과, 프루티콜과, 스트렙탁스과 등의 자바 펄몬산염 육지 껍질에 대한 비판적 연구"Treubia 20(3): 381–505, 107개의 무화과.
- ^ 반 벤트헴 주팅 T. (1959년)."수마트라의 비해상 몰루스카와 그 위성 섬의 카탈로그"보포르티아 7(83): 41–191, 1판, 11개의 무화과.
- ^ a b c (독일어로) 하니엘 C. (1921년)"변량학, Timoresischen Ampidromus Arten".Zeits. 인덕트. 에프텀. Vererbungsl 25(1–2): 88pp, 5판.
- ^ (독일어로) 질치 A. M. (1953)"Die Typen und Typoide des Natur-Museums Sencenberg. 10: 몰루스카, 플레로돈타과 (1)."아치브 퓌르 몰루스켄쿤데 82(4–6): 131–140, 플레이트 22–25.
- ^ a b c d e f g Sutcharit, C. (2005). "Taxonomic review of the tree snail Amphidromus Albers, 1850 (Pulmonata: Camaenidae) in Thailand and adjacent areas: subgenus Amphidromus". Journal of Molluscan Studies. 72: 1–30. doi:10.1093/mollus/eyi044.
- ^ a b c d e f g h i Sutcharit, C.; Asami, T.; Panha, S. (2007). "Evolution of whole-body enantiomorphy in the tree snail genus Amphidromus". Journal of Evolutionary Biology. 20 (2): 661–672. doi:10.1111/j.1420-9101.2006.01246.x. PMC 1920546. PMID 17305832.
- ^ (독일어로) 자코비 A. (1895)"Anatomische Untersuchen an Malayischen Landschnen."아치. 나튀르크 61: 293–318, 번호판 14.
- ^ (독일어로) 위그만 C. A. F.(1863년)."Beitrage jur Anatomie der Landschnechn des Underchen cords".인: 베버(1863년).졸. 어럽. 라이젠 무바첸 아치 3: 112–259, 번호판 9–16.
- ^ (독일어로) 위그만 C. A. F. (1898)"랜드몰러스켄(Stylommatophoren)"주토미셔 틸. 아블. 센켄. 나투르프. 제셀. 24(3): 289–557, 번호판 21–31. 514-527.
- ^ 콜링게 W. E. (1901)"암피드로무스 팰리스쿠스의 해부학에 관한 노트"주르. 말락 8(2): 50-52, 4번 판.
- ^ 콜링게 W. E. (1902)"말레이 반도의 '스케이트 원정대' 회원들이 수집한 비수술식 땅과 민물 연체동물에 대하여, 1899–1900"Op. cit. 9(3): 71–95, 플레이트 4–6.
- ^ Cuezzo, M.G. (2003). "Phylogenetic analysis of the Camaenidae (Mollusca: Stylommatophora) with special emphasis on the American taxa". Zoological Journal of the Linnean Society. 138 (4): 449–476. doi:10.1046/j.1096-3642.2003.00061.x.
- ^ Bishop, M.J. (1977). "Anatomical notes on some Javanese Amphidromus (Pulmonata: Camaenidae)". Journal of Conchology. 29: 199–205.
- ^ a b c Solem, A (1983). "First record of Amphidromus from Australia with anatomical note on several species (Mollusca: Pulmonata: Camaenidae)". Records of the Australian Museum. 35 (4): 153–166. doi:10.3853/j.0067-1975.35.1983.315. Archived from the original on 6 July 2011.
- ^ Chan S.-Y., Tan S.K. & Abbas J. B. (2008)."인도네시아 로티섬에서 온 암피드로무스(신드로무스)의 새로운 종(Gastropoda: Pulmonata: Camaenae)에 대하여"Molluscan Paper 1: 1–5. PDF 2011년 7월 24일 웨이백 머신, 인터넷 아카이브에 보관
- ^ a b Chan S.-Y. & Tan S.K.(2008)."인도네시아 섬에서 온 암피드로무스(신드로무스)의 새로운 종(Gastropoda: Pulmonata: Camaenae)에 대하여"때때로 몰러스칸 논문 1: 6–10.2011년 7월 24일 웨이백 머신, 인터넷 아카이브에 PDF 보관
- ^ 폰 마르텐스 E. (1860).다이 헬리센 2부 184쪽
- ^ 질치 A. M. (1960).가스트로포다, 에우티네우라 핸드b. 팔라오줄 (6)2(4): 601–834, 그림 2112–2515. 623페이지.
- ^ 벌로우(1905)나흐르 D. 말락 게셀37: 83.
- ^ 벤트헴 주팅 T. (1932년)."삼풍동굴(자바 중심)에서 날아온 선사시대 포탄에 대하여."Treubia 14(1): 103–108, 5개의 무화과.
- ^ 반 벤트헴 주팅 T. (1937)."자바에 있는 화석 지평선의 비해양 몰루스카, 트리닐 동물원에 대한 특별 참조.졸, 메이드, 20: 83–180, 페이지 4–12
- ^ a b Lok A. F. S. L. & Tan S. K.(2008)."녹색 나무 달팽이 암피드로무스 atricallosus perakensis Fulton, 1901년 싱가포르의 지위와 그 생물학에 대한 고찰"싱가포르의 자연 1: 225–230.PDF
- ^ 파라비시니 E. (1921년)."Die Eiablage zweier Javanischer Landschnecken".아치브 퓌르 몰루스켄쿤데 53: 113–116, 2판.
- ^ Schilthuizen, M; Scott, B J; Cabanban, A S; Craze, P G (2005). "Population structure and coil dimorphism in a tropical land snail". Heredity. 95 (3): 216–220. doi:10.1038/sj.hdy.6800715. PMID 16077741.
- ^ a b c Schilthuizen, M.; Craze, P. G.; Cabanban, A. S.; Davison, A.; Stone, J.; Gittenberger, E.; Scott, B. J. (2007). "Sexual selection maintains whole-body chiral dimorphism in snails". Journal of Evolutionary Biology. 20 (5): 1941–1949. doi:10.1111/j.1420-9101.2007.01370.x. PMC 2121153. PMID 17714311.
- ^ Wee J. (2006년)."달팽이를 먹고 있는 붉은 왕관 바벳"http://besgroup.talfrynature.com/2006/06/01/red-crowned-barbet-feeding-on-a-snail/ 웨이백 머신에 2011년 6월 13일 보관.2010년 5월 9일에 접속.
- ^ Danaisawadi, P.; Asami, T.; Ota, H.; Sutcharit, C.; Panha, Somsak (5 April 2016). "A snail-eating snake recognizes prey handedness". Scientific Reports. 6 (1): 23832. Bibcode:2016NatSR...623832D. doi:10.1038/srep23832. PMC 4820687. PMID 27046345.
추가 읽기
- Craze, Paul G.; Bin Elahan, Berjaya; Schilthuizen, Menno (2006). "Opposite shell-coiling morphs of the tropical land snail Amphidromus martensi show no spatial-scale effects" (PDF). Ecography. 29 (4): 477–486. doi:10.1111/j.0906-7590.2006.04731.x.
- Dharma, E. (2008). "Penerapan sistem pakar dalam perancangan program identifikasi jenis siput-pohon Amphidromus di Indonesia. Part 1". Berita Solaris. 11 (2): 12–16.
- Dumrongrojwatana P, Mutchacheep S. & Senapin R. (2006)."모포메트릭스(Pulmonata: Camaenae) 기법을 사용하여 7개의 태국 목각 달팽이속 암피드로무스 알버, 1850년"제44회 카세타트 대학 연차총회, 2006년 1월 30일 ~ 2월 2일, 주제 과학: 7 페이지방콕에 있는 카세타트 대학교.
- Goldberg, R. L.; Severns, M. (1997). "Isolation and evolution of the Amphidromus in Nusa Tenggara". American Conchologist. 25 (2): 3–7. Archived from the original on 25 November 2017. Retrieved 26 September 2019.
- Panha, S.; Sutcharit, C.; Tongkerd, P.; Burch, J. B. (2001). "Morphogeography of an endemic tree snail genus Amphidromus of Thailand (Pulmonata: Camaenidae)". Of Sea and Shore. 24 (2): 106–113.
- Prasankok, P; Ota, H; Toda, M; Panha, S (2007). "Allozyme variation in the camaenid tree snails Amphidromus atricallosus (Gould, 1843) and A. inversus (Müller, 1774)". Zoological Science. 24 (2): 189–97. doi:10.2108/zsj.24.189. hdl:2433/108565. PMID 17409732. S2CID 3470462.
- Severns, M. (2003). "A quick explanation of Amphidromus". Of Sea and Shore. 25 (4): 228–231.
- Severns, M. (2006). "A new species and a new subspecies of Amphidromus from Atauro Island, East Timor (Gastropoda, Pulmonata, Camaenidae)". Basteria. 70: 23–28.
외부 링크
| 위키미디어 커먼즈에는 암피드로무스와 관련된 미디어가 있다. |


