액티노르히잘 식물
Actinorhizal plant액티노르히잘 식물은 질소 고정액인 액티노박테리아 프랑키아와 공생관계를 형성하는 능력이 특징인 혈관신생식물군이다. 이 연관성은 질소고정 뿌리결절의 형성으로 이어진다.
분류
액티노르히잘 식물은 혈관조영술 3종, 8종, 24종에 분포하는 디코틸돈이다.[1]
이 세 가지 명령은 로시드 족 내에서 하나의 쇄도를 형성하는데, 이것은 다른 주요 질소 고정 명령인 파발레스의 자매 택시다. 다티스카속(Datisca)을 제외한 모든 액티노르히잘 종은 나무나 관목이다. 많은 식물들은 알더, 베이베리, 스위트페른, 에이벤스, 산악불행, 코리아리아와 같은 온대지방에서 흔한 식물들이다. 몇몇 엘레아그누스 종과 바다벅톤은 먹을 수 있는 과일을 생산한다. 열대지방에서는 카수아리나스(Casuarinas)가 널리 재배되고 있다.[citation needed]
분포와 생태

액티노르히잘 식물은 남극대륙을 제외한 모든 대륙에서 발견된다. 질소고정 결절을 형성하는 그들의 능력은 열악한 토양에서 선택적 우위를 보장한다. 그러므로 대부분의 활엽수 식물은 모레인, 화산 흐름, 모래언덕처럼 이용 가능한 질소가 부족한 어린 토양을 식민지화하는 개척자 종이다.[2] 이러한 교란된 환경을 식민지화한 최초의 종에 속하게 되는 것은, 토양에 풍부한 작용과 생태계의 계승에서 다른 종들의 설립을 가능하게 하는 중요한 역할을 한다.[1][2] 알더와 같은 액티노르히잘 식물은 또한 무리지어 사는 숲에서도 흔하다.[2] 액티노르히잘 식물은 세계의 넓은 지역에서 질소 고정의 주요 원인이며, 특히 온대림에서 중요하다.[1] 일부 다른 종에 대해 측정한 질소 고정률은 연간 300kg에2 달하는 N/ha/year로 보고된 가장 높은 비율에 가깝다.[3]
진화 기원
결절 관련 화석 기록은 없지만 8700만년 전 퇴적된 퇴적물에서 현대의 액티노르히잘 종과 유사한 식물들의 화석 꽃가루가 발견되었다. 공생조합의 기원은 여전히 불확실하다. 프랑키아와 교제할 수 있는 능력은 다혈질 성격이며 아마도 서로 다른 계층에서 독자적으로 진화했을 것이다.[4] 그럼에도 불구하고, 액티노르히잘 식물과 레구메스의 양대 질소 고정 집단들은 모두 질소 고정 쇄설이라고 불리는 로시드 안에 있는 쇄설물의 일부분이기 때문에 비교적 가까운 조상을 공유하고 있다. 이 조상은 질소고정 박테리아와 공생하기 위해 "예비화"를 개발했고, 이로 인해 액티노르히잘과 레구메 종의 조상들이 공생 능력을 독자적으로 획득하게 되었을지도 모른다. 공생을 확립하는 데 사용되는 유전 프로그램은 아마도 훨씬 오래되고 널리 분포된 식물과 곰팡이 사이의 공생 관계인 근막 공생의 요소들을 모집했을 것이다.[5]
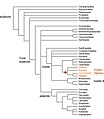
질소고정결절의 진화적 기원

공생결절
콩쿨에서와 같이, 끄덕임은 질소 결핍에 의해 선호되고 높은 질소 농도에 의해 억제된다. 식물 종에 따라 두 가지 감염 메커니즘이 설명되었다. 첫 번째는 카수아리나 알더에서 관찰되며 뿌리털 감염이라고 불린다. 이 경우 염증은 뿌리털의 세포내 감염으로 시작되어 특별한 조직인 혼전( pren戰)이 결여된 원시 공생기관의 형성이 뒤따른다.[6] 두 번째 감염 메커니즘은 세포간 침입이라고 불리며 디스카리아 종에 잘 설명되어 있다. 이 경우 박테리아는 세포외적으로 뿌리를 침투하여 표피세포 사이에서 자라고 피질세포 사이에서 성장한다. 나중에 프랭키아는 세포내부가 되었지만 혼전은 형성되지 않는다. 두 경우 모두 감염으로 인해 자전거의 세포 분열과 해부학적으로 측면 뿌리와 유사한 몇 개의 로브로 구성된 새로운 장기가 형성된다. 이 기관은 액티노르히잘 결절이라고도 불리는 액티노르히잘 결절이다. 결절의 피질 세포는 감염 부위나 혼전 부위에서 나오는 프랑키아 필라멘트의 침입을 받는다. 액티노르히잘 결절은 일반적으로 불확실한 성장을 하고, 따라서 새로운 세포가 정점에서 지속적으로 생산되어 연속적으로 감염된다. 결절의 성숙한 세포는 적극적으로 질소를 고정하는 박테리아 필라멘트로 채워진다. 결절로 이어지는 메커니즘에 관한 정보는 거의 없다. 리조비알 결절 인자에 해당하는 것은 발견되지 않았으나 레구메 결절(헤모글로빈 및 기타 결절의 부호화)의 형성과 기능에 관여하는 것으로 알려진 여러 유전자가 비슷한 역할을 해야 하는 액티노르히잘 식물에서도 발견된다.[7] Frankia과actinorhizal 종들을 유전자는 도구의 부족하다는 것의 주된 요인은 이 공생 때문입니다. 그런 가난한 understating이라고 설명했지만, 3Frankia의 최근의 염기 서열 결정 법[8]과 RNAi의 개발과actinorhizal 종의[9][10]다음 해에서 훨씬 더 잘 이해하는데 도움이 되어야 한다 유전자 도구들 밝혀.[표창 필요한]
메모들
참조
- Wall, L. (2000), "The actinorhizal symbiosis", Journal of Plant Growth and Regulation, 19 (2): 167–182, doi:10.1007/s003440000027, PMID 11038226, S2CID 12887261
- Schwintzer, C. R.; Tjepkema, J. (1990), The Biology of Frankia and Actinorhizal Plants, Academic Press, ISBN 978-0126332100
- Benson, D. R.; Clawson, M. L. (2000), "Evolution of the actinorhizal plant nitrogen-fixing symbiosis", in Triplett, E. (ed.), Prokaryotic Nitrogen Fixation: A Model System for the Analysis of a Biological Process, Norfolk, UK: Horizon Scientific Press, pp. 207–224, ISBN 978-1-898486-19-0
- Zavitovski, J.; Newton, M. (1968), "Ecological importance of snowbrush Ceanothus velutinus in the Oregon cascade", Ecology, 49 (6): 1134–1145, doi:10.2307/1934497, JSTOR 1934497
- Kistner, C.; Parniske, M. (2002), "Evolution of signal transduction in intracellular symbiosis", Trends in Plant Science, 7 (11): 511–518, doi:10.1016/S1360-1385(02)02356-7, PMID 12417152
- Laplaze, L.; Duhoux, E.; Franche, C.; Frutz, T.; Svistoonoff, S.; Bisseling, T.; Bogusz, D.; Pawlowski, K. (2000), "Casuarina glauca prenodule cells display the same differentiation as the corresponding nodule cells", Molecular Plant-Microbe Interactions, 13 (1): 107–112, doi:10.1094/MPMI.2000.13.1.107, PMID 10656591
- Vessey, J. K.; Pawlowski, K.; Bergman, B. (2005), "Root-based N2-fixing symbioses: Legumes, actinorhizal plants, Parasponia sp. and cycads", Plant and Soil, 266 (1): 205–230, doi:10.1007/s11104-005-0871-1, S2CID 35247396
- Gherbi, H.; Markmann, K.; Svistoonoff, S.; Estevan, J.; Autran, D.; Giczey, G.; Auguy, F.; Péret, B.; Laplaze, L.; Franche, C.; Parniske, M.; Bogusz, D. (2008), "SymRK defines a common genetic basis for plant root endosymbioses with arbuscular mycorrhiza fungi, rhizobia, and Frankia bacteria", Proceedings of the National Academy of Sciences, 105 (12): 4928–4932, doi:10.1073/pnas.0710618105, PMC 2290763, PMID 18316735
- Hocher, V.; Auguy, F.; Argout, X.; Laplaze, L.; Franche, C.; Bogusz, D. (2006), "Expressed sequence-tag analysis in Casuarina glauca actinorhizal nodule and root", New Phytologist, 169 (4): 681–688, doi:10.1111/j.1469-8137.2006.01644.x, PMID 16441749
- Normand, P.; Lapierre, P.; Tisa, L. S.; Gogarten, J. P.; Alloisio, N.; Bagnarol, E.; Bassi, C. A.; Berry, A. M.; Bickhart, D. M.; Choisne, N.; Couloux, A.; Cournoyer, B.; Cruveiller, S.; Daubin, V.; Demange, N.; Francino, M. P.; Goltsman, E.; Huang, Y.; Kopp, O. R.; Labarre, L.; Lapidus, A.; Lavire, C.; Marechal, J.; Martinez, M.; Mastronunzio, J. E.; Mullin, B. C.; Niemann, J.; Pujic, P.; Rawnsley, T.; Rouy, Z. (2006). "Genome characteristics of facultatively symbiotic Frankia sp. Strains reflect host range and host plant biogeography". Genome Research. 17 (1): 7–15. doi:10.1101/gr.5798407. PMC 1716269. PMID 17151343.
외부 링크
| 위키미디어 커먼즈에는 액티노르히잘 공장과 관련된 미디어가 있다. |
| 무료 사전인 Wiktionary에 있는 Actinorhizal 공장을 찾아보십시오. |

